
Antagonism and Antimicrobial Capacity of Epiphytic and Endophytic Bacteria against the Phytopathogen Xylella fastidiosa

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation of Endophytic and Epiphytic Bacteria
2.2. Screening of Antagonistic Activity In Vitro
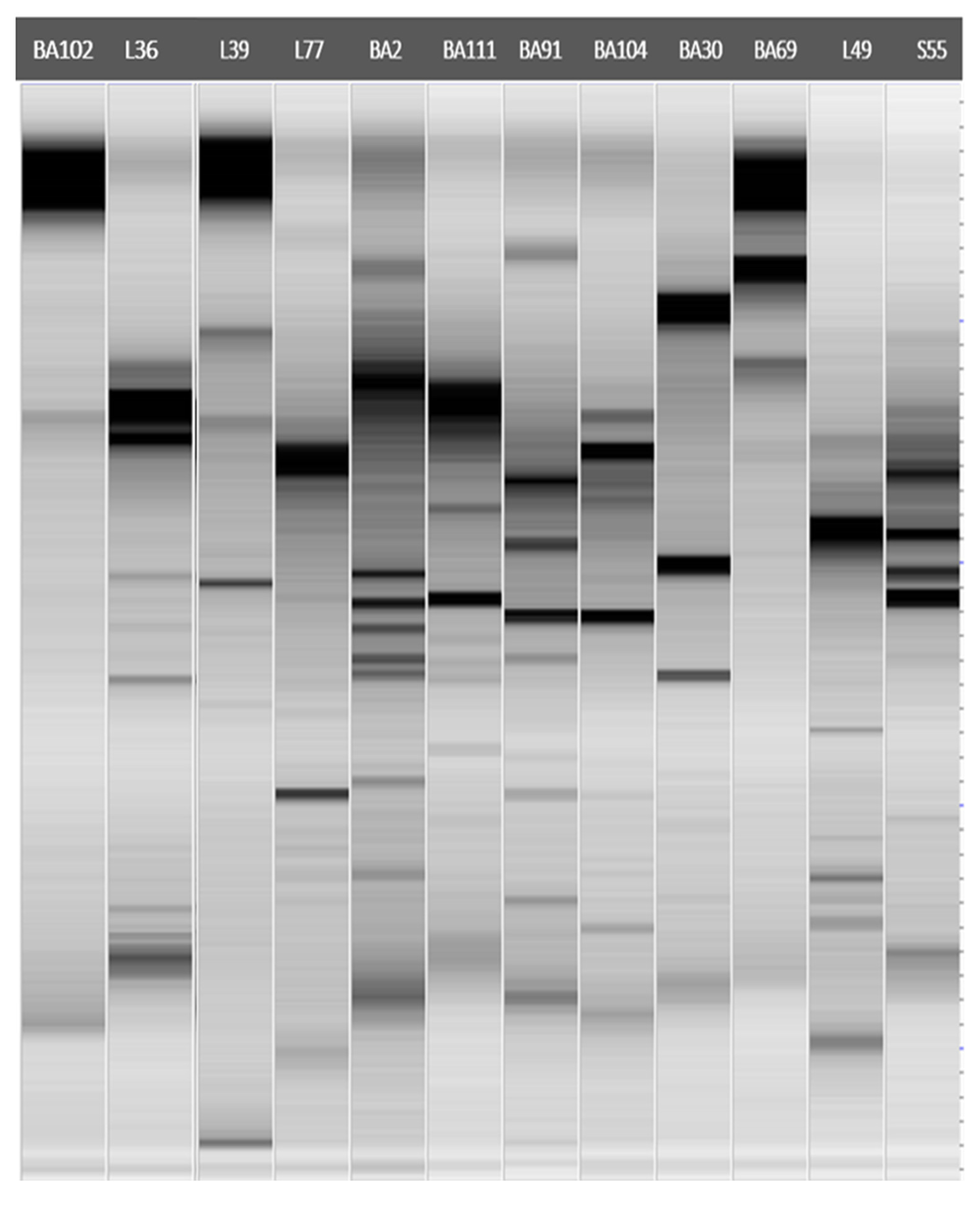
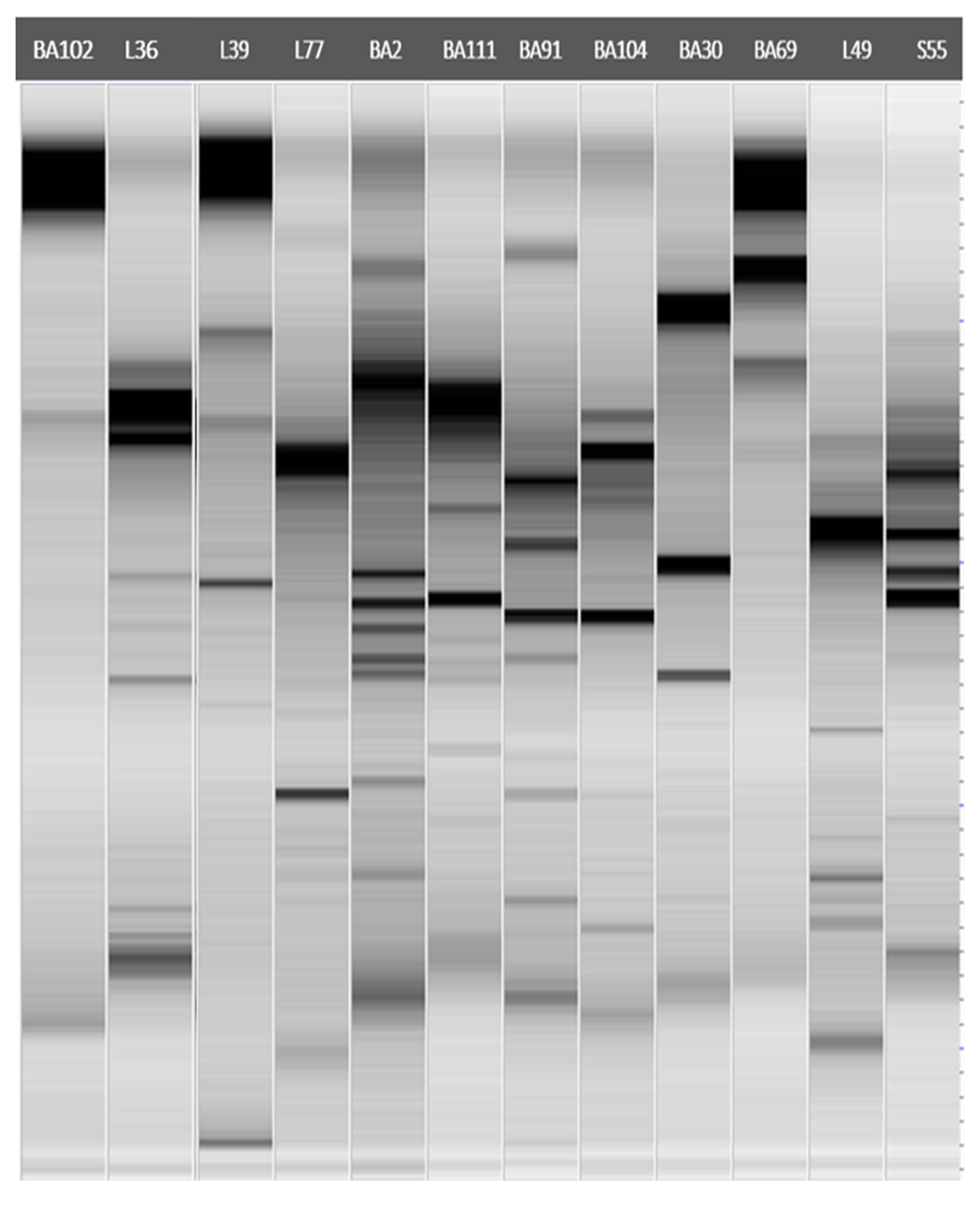
2.3. Molecular Characterization and Identification of Antagonistic Bacteria
2.4. Antimicrobial Activity against X. fastidiosa in Culture Filtrates of Antagonistic Strains
2.5. Statistical Analysis
3. Results
3.1. In Vitro Antagonistic Activity against X. fastidiosa
3.2. Molecular Characterization and Identification of Antagonistic Bacteria
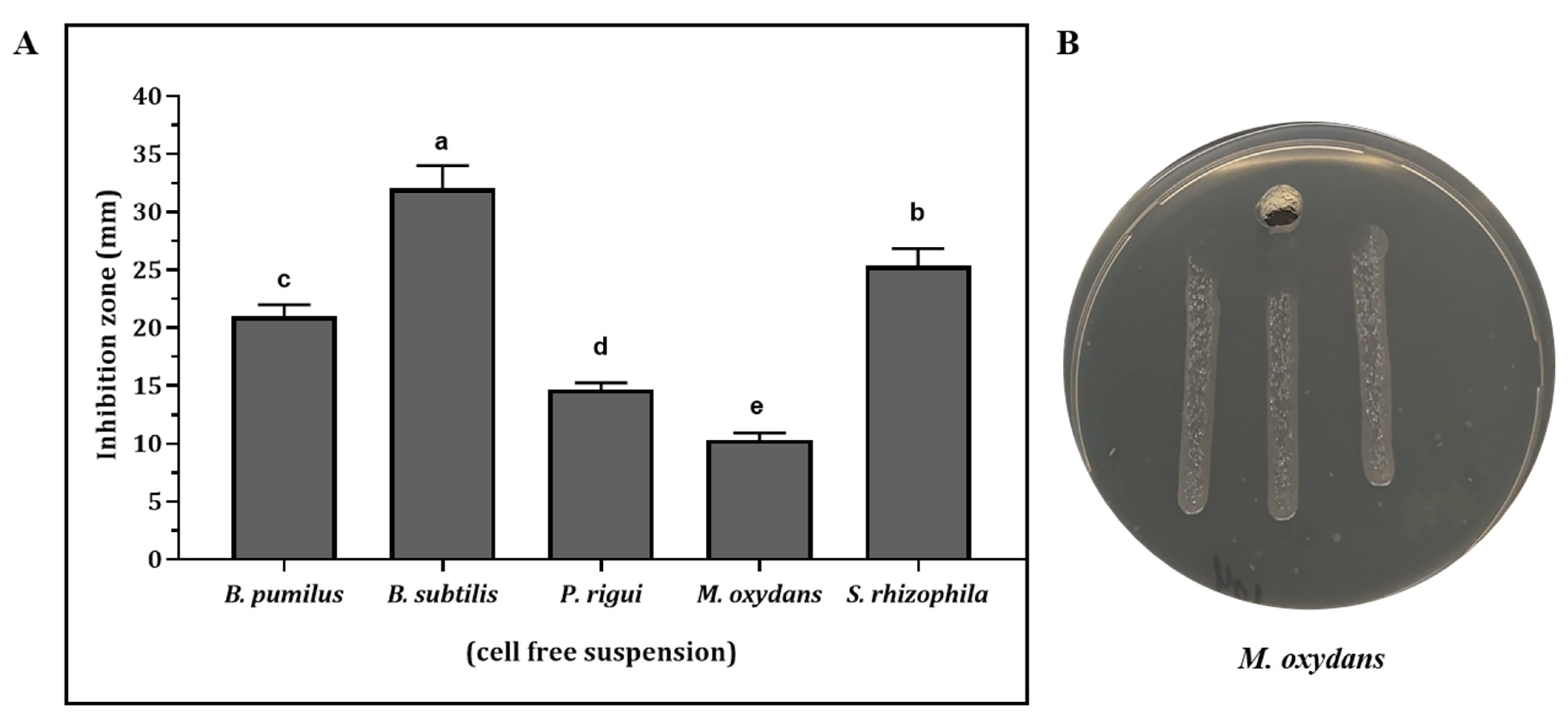
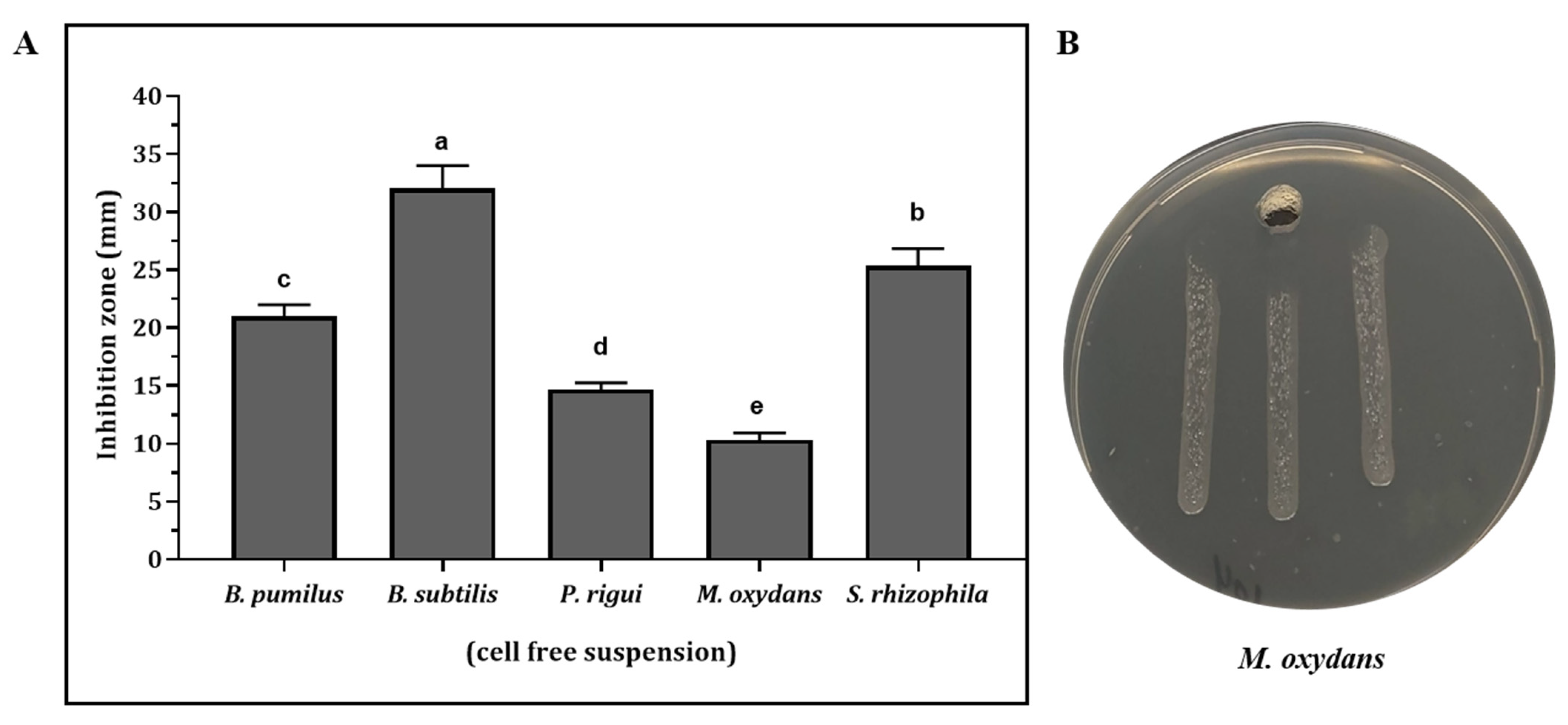
3.3. Antimicrobial Activity against X. fastidiosa in Culture Filtrates of Antagonistic Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saponari, M.; Boscia, D.; Nigro, F.; Martelli, G.P. Identification of DNA Sequences Related to Xylella fastidiosa in Oleander, Almond and Olive Trees Exhibiting Leaf Scorch Symptoms in Apulia (Southern Italy). J. Plant Pathol. 2013, 95, 659–668. [Google Scholar]
- Loconsole, G.; Potere, O.; Boscia, D.; Altamura, G.; Djelouah, K.; Elbeaino, T.; Frasheri, D.; Lorusso, D.; Palmisano, F.; Pollastro, P.; et al. Detection of Xylella fastidiosa in Olive Trees by Molecular and Serological Methods. J. Plant Pathol. 2014, 96, 8. [Google Scholar]
- Giampetruzzi, A.; Saponari, M.; Loconsole, G.; Boscia, D.; Savino, V.N.; Almeida, R.P.P.; Zicca, S.; Landa, B.B.; Chacón-Diaz, C.; Saldarelli, P. Genome-Wide Analysis Provides Evidence on the Genetic Relatedness of the Emergent Xylella fastidiosa Genotype in Italy to Isolates from Central America. Phytopathology 2017, 107, 816–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbeaino, T.; Valentini, F.; Abou Kubaa, R.; Moubarak, P.; Yaseen, T.; Digiaro, M. Multilocus Sequence Typing of Xylella fastidiosa Isolated from Olive Affected by” Olive Quick Decline Syndrome” in Italy. Phytopathol. Mediterr. 2014, 53, 533–542. [Google Scholar]
- Saponari, M.; Boscia, D.; Altamura, G.; Loconsole, G.; Zicca, S.; D’Attoma, G.; Morelli, M.; Palmisano, F.; Saponari, A.; Tavano, D.; et al. Isolation and Pathogenicity of Xylella fastidiosa Associated to the Olive Quick Decline Syndrome in Southern Italy. Sci. Rep. 2017, 7, 17723. [Google Scholar] [CrossRef]
- Martelli, G.P. The Current Status of the Quick Decline Syndrome of Olive in Southern Italy. Phytoparasitica 2016, 44, 1–10. [Google Scholar] [CrossRef]
- Maggiore, G.; Semeraro, T.; Aretano, R.; De Bellis, L.; Luvisi, A. GIS Analysis of Land-Use Change in Threatened Landscapes by Xylella fastidiosa. Sustainability 2019, 11, 253. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Plant Health (EFSA PLH Panel); Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.; Jaques Miret, J.A.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; et al. Effectiveness of in Planta Control Measures for Xylella fastidiosa. EFSA J. 2019, 17, 5666. [Google Scholar] [CrossRef]
- Pal, K.K.; McSpadden Gardener, B. Biological Control of Plant Pathogens. Plant Health Instr. 2006, 2, 1117–1141. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Klimova, E.; Rodríguez-Peña, K.; Sánchez, S. Endophytes as Sources of Antibiotics. Biochem. Pharmacol. 2017, 134, 1–17. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus Subtilis Group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biondi, E.; Gallipoli, L.; Mazzaglia, A.; Fuentealba, S.P.; Kuzmanović, N.; Bertaccini, A.; Balestra, G. Bacillus-Based Products for Management of Kiwifruit Bacterial Canker. Phytopathol. Mediterr. 2021, 60, 215–228. [Google Scholar] [CrossRef]
- Huang, R.; Feng, Z.; Chi, X.; Sun, X.; Lu, Y.; Zhang, B.; Lu, R.; Luo, W.; Wang, Y.; Miao, J.; et al. Pyrrolnitrin Is More Essential than Phenazines for Pseudomonas Chlororaphis G05 in Its Suppression of Fusarium graminearum. Microbiol. Res. 2018, 215, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Lacava, P.T.; Li, W.; Arau, W.L.; Hartung, J.S. The Endophyte Curtobacterium flaccumfaciens Reduces Symptoms Caused by Xylella fastidiosa in Catharanthus Roseus. Microbiology 2007, 45, 7. [Google Scholar]
- Baccari, C.; Antonova, E.; Lindow, S. Biological Control of Pierce’s Disease of Grape by an Endophytic Bacterium. Phytopathology 2019, 109, 248–256. [Google Scholar] [CrossRef] [Green Version]
- Zicca, S.; De Bellis, P.; Masiello, M.; Saponari, M.; Saldarelli, P.; Boscia, D.; Sisto, A. Antagonistic Activity of Olive Endophytic Bacteria and of Bacillus Spp. Strains against Xylella fastidiosa. Microbiol. Res. 2020, 236, 126467. [Google Scholar] [CrossRef]
- Hanani, A.; Valentini, F.; Sanzani, S.M.; Santoro, F.; Minutillo, S.A.; Gallo, M.; Cavallo, G.; Mourou, M.; El Moujabber, M.; D’Onghia, A.M.; et al. Community Analysis of Culturable Sapwood Endophytes from Apulian Olive Varieties with Different Susceptibility to Xylella fastidiosa. Agronomy 2021, 12, 9. [Google Scholar] [CrossRef]
- Brown, A.; Smith, H. Benson’s Microbiological Applications, Laboratory Manual in General Microbiology, Short Version; McGraw-Hill Education: New York, NY, USA, 2014. [Google Scholar]
- Etminani, F.; Harighi, B. Isolation and Identification of Endophytic Bacteria with Plant Growth Promoting Activity and Biocontrol Potential from Wild Pistachio Trees. Plant Pathol. J. 2018, 34, 208–217. [Google Scholar] [CrossRef]
- Balestra, G.M.; Agostini, R.; Bellincontro, A.; Mencarelli, F.; Varvaro, L. Bacterial Populations Related to Gerbera (Gerbera Jamesonii L.) Stem Break. Phytopathol. Mediterr. 2005, 44, 9. [Google Scholar]
- Schreiber, L.; Krimm, U.; Knoll, D.; Sayed, M.; Auling, G.; Kroppenstedt, R.M. Plant–Microbe Interactions: Identification of Epiphytic Bacteria and Their Ability to Alter Leaf Surface Permeability. New Phytol. 2005, 166, 589–594. [Google Scholar] [CrossRef]
- Minas, K.; McEwan, N.R.; Newbold, C.J.; Scott, K.P. Optimization of a High-Throughput CTAB-Based Protocol for the Extraction of QPCR-Grade DNA from Rumen Fluid, Plant and Bacterial Pure Cultures. FEMS Microbiol. Lett. 2011, 325, 162–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Versalovic, J.; Schneider, M.; De Bruijn, F.J.; Lupski, J.R. Genomic Fingerprinting of Bacteria Using Repetitive Sequence-Based Polymerase Chain Reaction. Methods Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Dymock, D.; Wade, W.G. Design and Evaluation of Useful Bacterium-Specific PCR Primers That Amplify Genes Coding for Bacterial 16S RRNA. Appl. Environ. Microbiol. 1998, 64, 2333. [Google Scholar] [CrossRef] [Green Version]
- Dagher, F.; Olishevska, S.; Philion, V.; Zheng, J.; Déziel, E. Development of a Novel Biological Control Agent Targeting the Phytopathogen Erwinia amylovora. Heliyon 2020, 6, e05222. [Google Scholar] [CrossRef]
- Borriss, R. Use of Plant-Associated Bacillus Strains as Biofertilizers and Biocontrol Agents in Agriculture. In Bacteria in Agrobiology: Plant Growth Responses; Springer: Berlin, Germany, 2011; pp. 41–76. [Google Scholar]
- Gardener, B.B.M.; Fravel, D.R. Biological Control of Plant Pathogens: Research, Commercialization, and Application in the USA. Plant Health Prog. 2002, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Legein, M.; Smets, W.; Vandenheuvel, D.; Eilers, T.; Muyshondt, B.; Prinsen, E.; Samson, R.; Lebeer, S. Modes of Action of Microbial Biocontrol in the Phyllosphere. Front. Microbiol. 2020, 11, 1619. [Google Scholar] [CrossRef]
- Thomashow, L.S.; Weller, D.M. Current Concepts in the Use of Introduced Bacteria for Biological Disease Control: Mechanisms and Antifungal Metabolites. In Plant-Microbe Interactions; Springer: New York, NY, USA, 1996; pp. 187–235. [Google Scholar]
- Daranas, N.; Roselló, G.; Cabrefiga, J.; Donati, I.; Francés, J.; Badosa, E.; Spinelli, F.; Montesinos, E.; Bonaterra, A. Biological Control of Bacterial Plant Diseases with Lactobacillus Plantarum Strains Selected for Their Broad-Spectrum Activity. Ann. Appl. Biol. 2019, 174, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Schiavi, D.; Balbi, R.; Giovagnoli, S.; Camaioni, E.; Botticella, E.; Sestili, F.; Balestra, G.M. A Green Nanostructured Pesticide to Control Tomato Bacterial Speck Disease. Nanomaterials 2021, 11, 1852. [Google Scholar] [CrossRef]
- Schiavi, D.; Ronchetti, R.; Di Lorenzo, V.; Salustri, M.; Petrucci, C.; Vivani, R.; Giovagnoli, S.; Camaioni, E.; Balestra, G.M. Circular Hazelnut Protection by Lignocellulosic Waste Valorization for Nanopesticides Development. Appl. Sci. 2022, 12, 2604. [Google Scholar] [CrossRef]
- Schiavi, D.; Francesconi, S.; Taddei, A.R.; Fortunati, E.; Balestra, G.M. Exploring Cellulose Nanocrystals Obtained from Olive Tree Wastes as Sustainable Crop Protection Tool against Bacterial Diseases. Sci. Rep. 2022, 12, 6149. [Google Scholar] [CrossRef]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant Beneficial Endophytic Bacteria: Mechanisms, Diversity, Host Range and Genetic Determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant Growth-Promoting Bacterial Endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Lumactud, R.; Shen, S.Y.; Lau, M.; Fulthorpe, R. Bacterial Endophytes Isolated from Plants in Natural Oil Seep Soils with Chronic Hydrocarbon Contamination. Front. Microbiol. 2016, 7, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, C.S.; Mrnka, L.; Lovecká, P.; Frantík, T.; Fenclová, M.; Demnerová, K.; Vosátka, M. Bacterial and Fungal Endophyte Communities in Healthy and Diseased Oilseed Rape and Their Potential for Biocontrol of Sclerotinia and Phoma Disease. Sci. Rep. 2021, 11, 3810. [Google Scholar] [CrossRef]
- Woźniak, M.; Gałązka, A.; Tyśkiewicz, R.; Jaroszuk-Ściseł, J. Endophytic Bacteria Potentially Promote Plant Growth by Synthesizing Different Metabolites and Their Phenotypic/Physiological Profiles in the Biolog GEN III MicroPlateTM Test. Int. J. Mol. Sci. 2019, 20, 5283. [Google Scholar] [CrossRef] [Green Version]
- Maignien, L.; DeForce, E.A.; Chafee, M.E.; Eren, A.M.; Simmons, S.L. Ecological Succession and Stochastic Variation in the Assembly of Arabidopsis Thaliana Phyllosphere Communities. MBio 2014, 5, e00682-13. [Google Scholar] [CrossRef] [Green Version]
- Delmotte, N.; Knief, C.; Chaffron, S.; Innerebner, G.; Roschitzki, B.; Schlapbach, R.; von Mering, C.; Vorholt, J.A. Community Proteogenomics Reveals Insights into the Physiology of Phyllosphere Bacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 16428–16433. [Google Scholar] [CrossRef] [Green Version]
- Lindow, S.E.; Brandl, M.T. Microbiology of the Phyllosphere. Appl Env. Microbiol 2003, 69, 9. [Google Scholar] [CrossRef] [Green Version]
- Vorholt, J.A. Microbial Life in the Phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Walterson, A.M.; Stavrinides, J. Pantoea: Insights into a Highly Versatile and Diverse Genus within the Enterobacteriaceae. FEMS Microbiol. Rev. 2015, 39, 968–984. [Google Scholar] [CrossRef] [Green Version]
- Irshad, A.; Ahmad, I.; Kim, S.B. Isolation, Characterization and Antimicrobial Activity of Halophilic Bacteria in Foreshore Soils. Afr. J. Microbiol. Res. 2013, 7, 164–173. [Google Scholar]
- Ozsahin, E.; Sezen, K.; Demir, I.; Demirbag, Z. Bacterial Isolates from Palomena prasina (Hemiptera: Pentatomidae) Include Potential Microbial Control Agents. Biocontrol Sci. Technol. 2014, 24, 1039–1051. [Google Scholar] [CrossRef]
- Kai, M.; Effmert, U.; Berg, G.; Piechulla, B. Volatiles of Bacterial Antagonists Inhibit Mycelial Growth of the Plant Pathogen Rhizoctonia solani. Arch. Microbiol. 2007, 187, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Perez, J.J.; Hernandez-Montiel, L.G.; Vero, S.; Noa-Carrazana, J.C.; Quiñones-Aguilar, E.E.; Rincón-Enríquez, G. Postharvest Biocontrol of Colletotrichum gloeosporioides on Mango Using the Marine Bacterium Stenotrophomonas rhizophila and Its Possible Mechanisms of Action. J. Food Sci. Technol. 2019, 56, 4992–4999. [Google Scholar] [CrossRef] [PubMed]
- Vida, C.; Cazorla, F.M.; de Vicente, A. Characterization of Biocontrol Bacterial Strains Isolated from a Suppressiveness-Induced Soil after Amendment with Composted Almond Shells. Res. Microbiol. 2017, 168, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Raza, W.; Ling, N.; Liu, D.; Wei, Z.; Huang, Q.; Shen, Q. Volatile Organic Compounds Produced by Pseudomonas Fluorescens WR-1 Restrict the Growth and Virulence Traits of Ralstonia solanacearum. Microbiol. Res. 2016, 192, 103–113. [Google Scholar] [CrossRef]
- Enespa; Chandra, P. Microbial Volatiles as Chemical Weapons Against Pathogenic Fungi. In Volatiles and Food Security; Choudhary, D.K., Sharma, A.K., Agarwal, P., Varma, A., Tuteja, N., Eds.; Springer: Singapore, 2017; pp. 227–254. ISBN 978-981-10-5552-2. [Google Scholar]
- Etschmann, M.; Bluemke, W.; Sell, D.; Schrader, J. Biotechnological Production of 2-Phenylethanol. Appl. Microbiol. Biotechnol. 2002, 59, 1–8. [Google Scholar]
- Cho, K.M.; Hong, S.Y.; Lee, S.M.; Kim, Y.H.; Kahng, G.G.; Lim, Y.P.; Kim, H.; Yun, H.D. Endophytic Bacterial Communities in Ginseng and Their Antifungal Activity Against Pathogens. Microb. Ecol. 2007, 54, 341–351. [Google Scholar] [CrossRef]
- Collazo, C.; Abadias, M.; Aguiló-Aguayo, I.; Alegre, I.; Chenoll, E.; Viñas, I. Studies on the Biocontrol Mechanisms of Pseudomonas graminis Strain CPA-7 against Food-Borne Pathogens in Vitro and on Fresh-Cut Melon. LWT Food Sci. Technol. 2017, 85, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Sun, L.; Dong, X.; Cai, Z.; Sun, X.; Yang, H.; Wang, Y.; Song, W. Characterization of a Novel Plant Growth-Promoting Bacteria Strain Delftia tsuruhatensis HR4 Both as a Diazotroph and a Potential Biocontrol Agent against Various Plant Pathogens. Syst. Appl. Microbiol. 2005, 28, 66–76. [Google Scholar] [CrossRef]
- Sahu, K.P.; Kumar, A.; Patel, A.; Kumar, M.; Gopalakrishnan, S.; Prakash, G.; Rathour, R.; Gogoi, R. Rice Blast Lesions: An Unexplored Phyllosphere Microhabitat for Novel Antagonistic Bacterial Species against Magnaporthe oryzae. Microb. Ecol. 2021, 81, 731–745. [Google Scholar] [CrossRef]
- Szentes, S.; Radu, G.-L.; Laslo, É.; Lányi, S.; Mara, G. Selection and Evaluation of Potential Biocontrol Rhizobacteria from a Raised Bog Environment. Crop. Prot. 2013, 52, 116–124. [Google Scholar] [CrossRef]
- Fiedler, S.; Heerklotz, H. Vesicle Leakage Reflects the Target Selectivity of Antimicrobial Lipopeptides from Bacillus subtilis. Biophys. J. 2015, 109, 2079–2089. [Google Scholar] [CrossRef] [Green Version]
- Pandin, C.; Le Coq, D.; Deschamps, J.; Védie, R.; Rousseau, T.; Aymerich, S.; Briandet, R. Complete Genome Sequence of Bacillus velezensis QST713: A Biocontrol Agent That Protects Agaricus Bisporus Crops against the Green Mould Disease. J. Biotechnol. 2018, 278, 10–19. [Google Scholar] [CrossRef]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.-C. Current Knowledge and Perspectives of Paenibacillus: A Review. Microb. Cell Factories 2016, 15, 203. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Rojas, F.U.; Salazar-Gómez, A.; Vargas-Díaz, M.E.; Vásquez-Murrieta, M.S.; Hirsch, A.M.; De Mot, R.; Ghequire, M.G.K.; Ibarra, J.A.; Estrada-de los Santos, P. Broad-Spectrum Antimicrobial Activity by Burkholderia cenocepacia TAtl-371, a Strain Isolated from the Tomato Rhizosphere. Microbiology 2018, 164, 1072–1086. [Google Scholar] [CrossRef]
- Korenblum, E.; von Der Weid, I.; Santos, A.L.S.; Rosado, A.S.; Sebastián, G.V.; Coutinho, C.; Magalhaes, F.C.M.; De Paiva, M.M.; Seldin, L. Production of Antimicrobial Substances by Bacillus Subtilis LFE-1, B. Firmus H2O-1 and B. Licheniformis T6-5 Isolated from an Oil Reservoir in Brazil. J. Appl. Microbiol. 2005, 98, 667–675. [Google Scholar] [CrossRef]
- Von der Weid, I.; Alviano, D.S.; Santos, A.L.S.; Soares, R.M.A.; Alviano, C.S.; Seldin, L. Antimicrobial Activity of Paenibacillus peoriae Strain NRRL BD-62 against a Broad Spectrum of Phytopathogenic Bacteria and Fungi. J. Appl. Microbiol. 2003, 95, 1143–1151. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of Plant Growth-Promoting Bacteria for Biocontrol of Plant Diseases: Principles, Mechanisms of Action, and Future Prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Poppe, L.; Vanhoutte, S.; Höfte, M. Modes of Action of Pantoea agglomerans CPA-2, an Antagonist of Postharvest Pathogens on Fruits. Eur. J. Plant Pathol. 2003, 109, 963–973. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents Against Plant Diseases: Relevance Beyond Efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Bacteria | Isolate | Host Plant | Accession Number | % of 16S rRNA identity | Inhibition Zone (mm) | |
|---|---|---|---|---|---|---|
| Epiphytes | Pseudomonas graminis | BA2 | Rosmarinus officinalis | MW769929 | 98.28 | 6 ± 0.82 f |
| Delftia acidovorans | BA30 | Rosmarinus officinalis | MW769937 | 100.0 | 8 ± 0.82 ef | |
| Pantoea agglomerans | BA69 | Nerium oleander | MW769928 | 99.91 | 34 ± 1.63 a | |
| Microbacterium oleivorans | BA91 | Olea europaea S | MW769930 | 99.03 | 4 ± 0.81 j | |
| Stenotrophomonas rhizophila | BA102 | Polygala myrtifolia | MW769932 | 100.0 | 12.3 ± 1.63 de | |
| Microbacterium oxydans | BA104 | Polygala myrtifolia | MW769934 | 99.82 | 16.6 ± 1.26 bc | |
| Microbacterium phyllosphaerae | BA111 | Polygala myrtifolia | MW769936 | 99.91 | 7 ± 0.81 f | |
| Endophytes | Paenibacillus naphthalenovorans | L49 | Olea europaea L | MZ047304 | 99.91 | 38.6 ± 0.94 a |
| Paenibacillus rigui | S55 | Olea europaea S | MZ047306 | 99.25 | 21.6 ± 0.47 b | |
| Pseudomonas hibiscicola | L77 | Olea europaea L | MZ047305 | 99.50 | 15 ± 0.4 cd | |
| Bacillus pumilus | L36 | Olea europaea L | MZ047307 | 99.69 | 9.8 ± 0.23 ef | |
| Bacillus subtilis | L39 | Olea europaea L | MZ047308 | 100.0 | 19 ± 0.4 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mourou, M.; Hanani, A.; D’Onghia, A.M.; Davino, S.W.; Balestra, G.M.; Valentini, F. Antagonism and Antimicrobial Capacity of Epiphytic and Endophytic Bacteria against the Phytopathogen Xylella fastidiosa. Agronomy 2022, 12, 1266. https://doi.org/10.3390/agronomy12061266
Mourou M, Hanani A, D’Onghia AM, Davino SW, Balestra GM, Valentini F. Antagonism and Antimicrobial Capacity of Epiphytic and Endophytic Bacteria against the Phytopathogen Xylella fastidiosa. Agronomy. 2022; 12(6):1266. https://doi.org/10.3390/agronomy12061266
Chicago/Turabian StyleMourou, Marwa, Arafat Hanani, Anna Maria D’Onghia, Salvatore Walter Davino, Giorgio Mariano Balestra, and Franco Valentini. 2022. "Antagonism and Antimicrobial Capacity of Epiphytic and Endophytic Bacteria against the Phytopathogen Xylella fastidiosa" Agronomy 12, no. 6: 1266. https://doi.org/10.3390/agronomy12061266
APA StyleMourou, M., Hanani, A., D’Onghia, A. M., Davino, S. W., Balestra, G. M., & Valentini, F. (2022). Antagonism and Antimicrobial Capacity of Epiphytic and Endophytic Bacteria against the Phytopathogen Xylella fastidiosa. Agronomy, 12(6), 1266. https://doi.org/10.3390/agronomy12061266