
Forage Yield, Quality, and Impact on Subsequent Cash Crop of Cover Crops in an Integrated Forage/Row Crop System

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
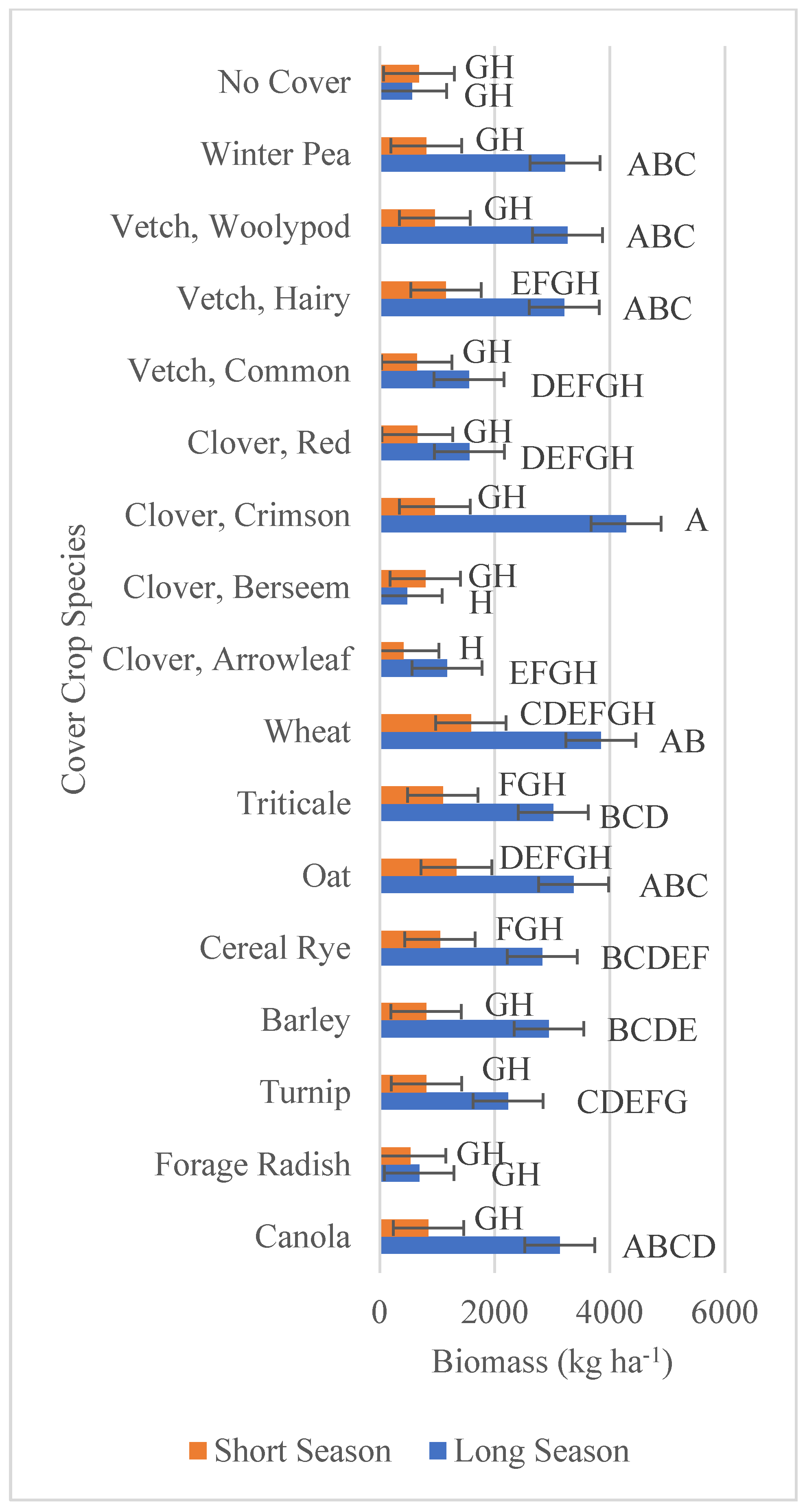
3.1. Forage Biomass and Nutritive Value
3.2. Cash Crop and Soil Attributes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaspar, T.C.; Bakker, M.G. Biomass production of 12 winter cereal cover crop cultivars and their effect on subsequent no-till corn yield. J. Soil Water Conserv. 2015, 70, 353–364. [Google Scholar] [CrossRef]
- Fraser, P.M.; Curtin, D.; Harrison-Kirk, T.; Meenken, E.D.; Beare, M.H.; Tabley, F.; Gillespie, R.N.; Franics, G.S. Winter nitrate leaching under different tillage and winter cover crop management practices. Soil Sci. Soc. Am. J. 2013, 77, 1391–1401. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C.; Bailey, B.A. Role of cover crops in improving soil and row crop productivity. Commun. Soil Sci. Plant Anal. 2005, 36, 2733–2757. [Google Scholar] [CrossRef]
- Baraibar, B.; Hunter, M.C.; Schipanski, M.E.; Hamilton, A.; Mortensen, D.A. Weed suppression in cover crop monocultures and mixtures. Weed Sci. 2018, 66, 121–133. [Google Scholar] [CrossRef]
- Snapp, S.S.; Swinton, S.M.; Labarta, R.; Mutch, D.; Black, J.R.; Leep, R.; Nyiraneza, J.; O’Neil, K. Evaluating cover crops for benefits, costs and performance within cropping system niches of crop and impact of foregoing a cash crop, some farmers express Michigan and New York producers are experimenting. Agron. J. 2005, 97, 322–332. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Barbercheck, M.; Douglas, M.R.; Finney, D.M.; Haider, K.; Kaye, J.P.; Kermaniam, A.R.; Mortensen, D.A.; Ryan, M.R.; Tooker, J.; et al. A framework for evaluating ecosystem services provided by cover crops in agroecosystems. Agric. Syst. 2014, 125, 12–22. [Google Scholar] [CrossRef]
- Hansen, M.J.; Owens, V.N.; Beck, D.; Sexton, P. Suitability of cover crop monocultures for late-season forage in South Dakota. Can. J. Plant Sci. 2013, 93, 589–597. [Google Scholar] [CrossRef]
- Appelgate, S.R.; Lenssen, A.W.; Wiedenhoeft, M.H.; Kaspar, T.C. Cover crop options and mixes for upper midwest corn–soybean systems. Agron. J. 2017, 109, 968–984. [Google Scholar] [CrossRef] [Green Version]
- Ketterings, Q.M.; Swink, S.N.; Duiker, S.W.; Czymmek, K.J.; Beegle, D.B.; Cox, W.J. Integrating cover crops for nitrogen management in corn systems on Northeastern U.S. dairies. Agron. J. 2015, 107, 1365–1376. [Google Scholar] [CrossRef] [Green Version]
- Sykes, V.R.; Wilson, A.; Bates, G.; McIntosh, D.; McClure, A.; Raper, T.; Blair, R.; Walker, F. Cover Crop Variety Tests in Tennessee 2020. In UT Ext. W953; University of Tennessee: Knoxville, TN, USA, 2020; Available online: https://search.utcrops.com/wp-content/uploads/2020/09/2020-TN-Cover-Crop-Variety-Test-FINAL-REPORT.pdf (accessed on 30 March 2022).
- Sykes, V.R.; Wilson, A.; Palacios, F.; Keadle, S.B.; Bates, G.; McIntosh, D. Cover Crop Variety Tests in Tennessee 2021. In UT Ext. W 1022; University of Tennessee: Knoxville, TN, USA, 2021; Available online: https://search.utcrops.com/wp-content/uploads/2021/09/W_1022_Cover_Crop_Variety_Tests_in_TN_2021.pdf (accessed on 30 March 2022).
- Gieske, M.F.; Ackroyd, V.J.; Baas, D.G.; Mutch, D.R.; Wyse, D.; Durgan, B.R. Brassica cover crop effects on nitrogen availability and oat and corn yield. Agron. J. 2016, 108, 151–161. [Google Scholar] [CrossRef]
- Ball, D.M.; Hoveland, C.S.; Lacefield, G.D. Southern Forages Modern Concepts for Forage Crop Management, 5th ed.; International Plant Nutrition Institute: Peachtree Corners, GA, USA, 2015. [Google Scholar]
- Contreras-Govea, F.E.; Albrecht, K.A. Forage production and nutritive value of oat in autumn and early summer. Crop Sci. 2006, 46, 2382–2386. [Google Scholar] [CrossRef]
- Sulc, R.M.; Franzlubber, A.J. Exploring integrated crop-livestock systems in different ecoregions of the United States. Eur. J. Agron. 2014, 57, 21–30. [Google Scholar] [CrossRef]
- Fae, G.S.; Sulc, R.M.; Barker, D.J.; Dick, R.P.; Eastridge, M.L.; Lorenz, N. Integrating winter annual forages into a no-till corn silage system. Agron. J. 2009, 101, 1286–1296. [Google Scholar] [CrossRef]
- Iglesias, I.; Lloveras, J. Annual cool-season legumes for forage production in mild winter areas. Grass Forage Sci. 1998, 53, 318–325. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Stuedmann, J.A. Does grazing of cover crops impact biologically active soil carbon and nitrogen fractions under inversion or no tillage management? Soil Water Conserv. 2015, 70, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.N. Effects of cereal and legume cover crop residues on weeds, yield, and net return in soybean (Glycine max). Weed Technol. 2001, 15, 660–668. [Google Scholar] [CrossRef]
- Ruffo, M.L.; Bullock, D.G.; Bollero, G.A. Soybean yield as affected by biomass and nitrogen uptake of cereal rye in winter cover crop rotations. Agron. J. 2004, 96, 800–805. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Stuedmann, J.A. Crop and cattle responses to tillage systems for integrated crop-livestock production in the Southern Piedmont, USA. Renew. Agric. Food Syst. 2006, 22, 168–180. [Google Scholar] [CrossRef] [Green Version]
- Bracey, W. Evaluating Cool-Season Annual Species as Cover Crops and Dual-Use Forage Cover Crops in No-Till Tennessee Production Systems. University of Tennessee: Knoxville, TN, USA, 2020; Available online: https://trace.tennessee.edu/cgi/viewcontent.cgi?article=7174&context=utk_gradthes (accessed on 30 March 2022).
- Staff, S.S. Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; Natural Resources Conservation Service; U.S. Government Printing Office: Washington, DC, USA, 1999.
- Bates, G.; Harper, C.; McClure, A.; Raper, T.; Sykes, V.R. Forage and Field Crop Seeding Guide for Tennessee. In UT Ext. PB 378; University of Tennessee: Knoxville, TN, USA, 2016; Available online: https://extension.tennessee.edu/publications/Documents/pb378.pdf (accessed on 30 March 2022).
- McIntosh, D.; Anderson-Husmoen, B.J.; Kern-Lunbery, R.; Goldblatt, P.; Lemus, R.; Griggs, T.; Bauman, L.; Boone, S.; Shewmaker, G.; Teutsch, C. Guidelines for Optimal Use of NIRSC Forage and Feed Calibrations in Membership Laboratories, 2nd ed.; The University of Tennessee Press: Knoxville, TN, USA, 2022. [Google Scholar]
- National Academies of Sciences, Engineering, and Medicine. Nutrient Requirements of Beef Cattle, 8th ed.; The National Academies Press: Washington, DC, USA, 2016. [Google Scholar]
- Woodruff, L.K.; Hitchcock, R.; Sonon, L.; Saha, U.; Kissel, D.E.; Gaskin, J.; Romano, N.; Cabrera, M.L.; Habteselassie, M.Y.; Vigil, M.; et al. A web-based model of N mineralization from cover crop residue decomposition. Soil Sci. Soc. Am. J. 2018, 82, 983–993. [Google Scholar] [CrossRef]
- Gaskin, J.; Cabrera, M.; Kissel, D. Predicting Nitrogen Release from Cover Crops: The Cover Crop Nitrogen Availability Calculator. In UGA Ext. Bulletin 1466; University of Georgia: Athens, GA, USA, 2016; Available online: http://aesl.ces.uga.edu/publications/soil/B1466.pdf (accessed on 30 March 2022).
- Haney, R.; Brinton, W.; Evans, E. Soil CO2 respiration: Comparison of chemical titration, CO2 IRGA analysis and the Solvita gel system. Renew. Agric. Food Syst. 2008, 23, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Haney, R.; Haney, E.B.; Smith, D.R.; Harmel, R.D.; White, M.J. The soil health tool—Theory and initial broad-scale application. Appl. Soil Ecol. 2018, 125, 162–168. [Google Scholar] [CrossRef]
- Rakoczy-Trojanowska, M.; Swiecicka, M.; Bakera, B.; Kowalczyk, M.; Stochmal, A.; Bolibok, L. Cocultivating rye with berseem clover affects benzoxazinoid production and expression of related genes. Crop Sci. 2020, 60, 3228–3246. [Google Scholar] [CrossRef]
- Gill, W.; Lane, C.; Neel, J.; Fisher, A.; Bates, G.; Joines, D. Mineral Nutrition of Beef Cattle. In UT Ext. PB 1749; University of Tennessee: Knoxville, TN, USA, 2004; Available online: https://trace.tennessee.edu/cgi/viewcontent.cgi?article=1003&context=utk_agexani (accessed on 30 March 2022).
- Wyland, L.J.; Jackson, L.; Chaney, W.E.; Klonsky, K.; Koike, S.T.; Kimple, B. Winter cover crops in a vegetable cropping system: Impacts on yield, nitrate leaching, pests and management costs. Agric. Ecosyst. Environ. 1996, 59, 1–17. [Google Scholar] [CrossRef]
- Hu, S.; Grunwald, N.J.; Van Bruggen, A.H.C.; Gamble, G.R.; Drinkwater, L.E.; Shennan, C.; Demment, M.W. Short-term effects of cover crop incorporation on soil carbon pools and nitrogen availability. Soil Sci. Soc. Am. J. 1997, 61, 901–911. [Google Scholar] [CrossRef]
- Parr, M.; Grossman, J.M.; Reberg-Horton, S.C.; Brinton, C.; Crozier, C. Roller-crimper termination for legume cover crops in North Carolina: Impacts on nutrient availability to a succeeding corn crop. Commun. Soil Sci. Plant Anal. 2014, 45, 1106–1119. [Google Scholar] [CrossRef]
- Richter, D.D.B.; Hofmockel, M.; Callaham, M.A.; Powlson, D.S.; Smith, P. Long-term soil experiments: Keys to managing Earth’s rapidly chancing ecosystems. Soil Sci. Soc. Am. J. 2007, 71, 266–279. [Google Scholar] [CrossRef]
- Boyer, C.N.; Lambert, D.M.; Larson, J.A.; Tyler, D.D. Investment analysis of cover crop and no-tillage systems on Tennessee cotton. Agron. J. 2018, 1, 331–338. [Google Scholar] [CrossRef]

{kind=link}
| Cover Crop | Seeding Rate (kg ha−1) | Planting Depth (cm) | Variety | |
|---|---|---|---|---|
| 2017–2018 | 2018–2019 | |||
| Clover, arrowleaf | 11.21 | 1.27 | Yucchi | VNS |
| Clover, crimson | 28.03 | 1.27 | Dixie | Dixie |
| Clover, red | 11.21 | 1.27 | VNS | VNS |
| Vetch, common | 33.63 | 5.08 | VNS | VNS |
| Vetch, hairy | 33.63 | 5.08 | VNS | VNS |
| Vetch, woolypod | 33.63 | 5.08 | Lana | Lana |
| Winter pea | 56.05 | 5.08 | VNS | VNS |
| Barley | 168.15 | 5.08 | Bob Winter | Bob Winter |
| Oat | 168.15 | 5.08 | Arivat | Secretariat |
| Triticale | 168.15 | 5.08 | Fridge | Fridge |
| Cereal rye | 168.15 | 5.08 | VNS | VNS |
| Wheat | 201.78 | 5.08 | Arthur | VNS |
| Canola | 8.97 | 1.27 | Edimax CL | Edimax CL |
| Forage radish | 16.82 | 1.27 | Torpedo | Torpedo |
| Turnip | 6.73 | 1.27 | Purple top white Globe | Purple top white Globe |
| Cover Crops | Cash Crops | |||||||
|---|---|---|---|---|---|---|---|---|
| Year | Location | Season | Establishment | Forage Harvest | Termination | Planting | Harvest | Crop |
| 2017–2018 | ETREC | Short | 27/10/17 | 11/4/18 | 18/4/18 | 7/5/18 | 3/10/18 | Corn |
| MTREC | 25/10/17 | 4/4/18 | 11/4/18 | 30/4/18 | 11/9/18 | |||
| ETREC | Long | 25/9/17 | 20/4/18 | 30/4/18 | 24/5/18 | 9/10/18 | Soybean | |
| MTREC | 25/917 | 18/4/18 | 27/4/18 | 11/5/18 | 22/10/18 | |||
| 2018–2019 | ETREC | Short | 31/10/18 | 1/4/19 | 11/4/19 | 16/5/19 | 18/9/19 | Corn |
| MTREC | 24/10/18 | 28/3/19 | 2/4/19 | 18/4/19 | 19/9/19 | |||
| ETREC | Long | 12/10/18 | 15/4/19 | 24/4/19 | 9/5/19 | 2/10/19 | Soybean | |
| MTREC | 5/10/18 | 11/4/19 | 18/4/19 | 8/5/19 | 25/9/19 | |||
| Effect | Season | Species | Season × Species |
|---|---|---|---|
| Cover Crop Biomass | 0.05 | <0.001 | <0.001 |
| Carbon | 0.51 | 0.39 | 0.98 |
| Nitrogen | 0.98 | <0.001 | <0.001 |
| Est. N Release—2 wks | 0.07 | <0.001 | <0.001 |
| Est. N Release—4 wks | 0.08 | <0.001 | <0.001 |
| Est. N Release—12 wks | 0.10 | <0.001 | <0.001 |
| Ca | 0.21 | <0.001 | 0.95 |
| K | 0.30 | <0.001 | 0.007 |
| P | 0.31 | <0.001 | 0.02 |
| Mg | 0.94 | <0.001 | 0.11 |
| CP | 0.80 | <0.001 | <0.001 |
| Ash | 0.74 | <0.001 | 0.02 |
| Fat | 0.41 | 0.21 | 0.81 |
| Lignin | 0.83 | <0.001 | 0.36 |
| ADF | 0.34 | <0.001 | <0.001 |
| NDF | 0.13 | <0.001 | 0.11 |
| dNDF48 | 0.17 | <0.001 | 0.13 |
| IVTDM48 | 0.36 | 0.02 | 0.37 |
| TDN | 0.33 | <0.001 | 0.007 |
| Species | Ca (g kg−1) | Mg (g kg−1) | Lignin (g kg−1) | NDF (g kg−1) | dNDF48 (g kg−1) | IVTDM48 (g kg−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Canola | 10 | AB | 2 | EF | 61 | BCDE | 456 | DE | 238 | DEF | 802 | BCDE |
| Forage Radish | 11 | AB | 3 | ABC | 69 | A | 462 | CDE | 239 | DEF | 788 | E |
| Turnip | 11 | AB | 3 | ABCD | 64 | ABC | 464 | BCDE | 243 | DEF | 791 | DE |
| Barley | 4 | C | 2 | EFG | 42 | F | 523 | A | 347 | AB | 816 | ABCDE |
| Cereal Rye | 4 | C | 2 | FG | 38 | F | 520 | AB | 356 | A | 832 | ABCD |
| Oat | 5 | C | 2 | FG | 39 | F | 449 | DE | 315 | C | 844 | AB |
| Triticale | 4 | C | 2 | FG | 40 | F | 515 | ABC | 353 | AB | 825 | ABCDE |
| Wheat | 4 | C | 1 | G | 45 | F | 494 | ABCD | 330 | BC | 821 | ABCDE |
| Clover, Arrowleaf | 10 | AB | 2 | CD | 66 | AB | 452 | DE | 246 | DE | 800 | CDE |
| Clover, Berseem | 10 | AB | 2 | DE | 66 | AB | 452 | DE | 229 | DEF | 782 | E |
| Clover, Crimson | 12 | A | 3 | ABC | 56 | DE | 393 | F | 218 | F | 837 | ABC |
| Clover, Red | 12 | A | 3 | ABC | 64 | ABCD | 413 | EF | 222 | EF | 810 | ABCDE |
| Vetch, Common | 9 | B | 3 | ABC | 58 | CDE | 423 | EF | 251 | D | 837 | ABC |
| Vetch, Hairy | 10 | AB | 3 | AB | 55 | E | 423 | EF | 253 | D | 847 | A |
| Vetch, Woolypod | 11 | AB | 3 | A | 58 | CDE | 426 | EF | 246 | DE | 846 | A |
| Winter Pea | 10 | AB | 3 | BCD | 56 | DE | 427 | EF | 249 | D | 845 | AB |
| Mean | 8 | 2 | 55 | 456 | 271 | 820 | ||||||
| Standard Error | 3 | 0 | 9 | 34 | 18 | 18 | ||||||
| N (g kg−1) | K (g kg−1) | P (g kg−1) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Short | Long | Short | Long | Short | Long | |||||||||
| Canola | 24 | BCD | 21 | DEFG | 19 | AB | 21 | BCDE | 3.0 | ABC | 2.3 | DE | * | ||
| Forage Radish | 26 | ABCD | 24 | CDE | 17 | AB | 18 | CDE | 2.8 | BCDE | 2.6 | CD | |||
| Turnip | 21 | DEF | 17 | GHI | 19 | AB | 19 | CDE | 2.9 | ABCD | 2.3 | DE | |||
| Barley | 22 | DE | 18 | FGH | 16 | B | 17 | EF | 2.6 | CDE | 2.1 | EF | |||
| Cereal Rye | 23 | CDE | 19 | EFGH | 17 | AB | 18 | DEF | 2.6 | BCDE | 2.2 | EF | |||
| Oat | 16 | F | 7 | J | * | 17 | AB | 13 | FG | 2.6 | CDE | 1.9 | EF | * | |
| Triticale | 18 | EF | 13 | HIJ | 18 | AB | 17 | EF | 2.5 | DE | 2.1 | EF | |||
| Wheat | 18 | EF | 12 | IJ | 18 | AB | 12 | G | * | 2.4 | E | 1.8 | F | * | |
| Clover, Arrowleaf | 22 | DEF | 27 | BCD | 16 | B | 18 | CDE | 2.9 | ABCD | 3.1 | ABC | |||
| Clover, Berseem | 24 | BCDE | 24 | CDEF | 15 | B | 19 | CDE | 2.8 | BCDE | 2.8 | BCD | |||
| Clover, Crimson | 28 | ABC | 29 | BC | 17 | AB | 25 | AB | * | 3.0 | ABC | 2.9 | BC | ||
| Clover, Red | 22 | CDE | 33 | AB | * | 17 | AB | 23 | ABC | 3.0 | AB | 3.2 | AB | ||
| Vetch, Common | 20 | DEF | 28 | BC | * | 20 | AB | 25 | AB | 3.0 | AB | 3.1 | ABC | ||
| Vetch, Hairy | 29 | AB | 36 | A | 18 | AB | 24 | AB | 3.3 | A | 3.4 | A | |||
| Vetch, Woolypod | 30 | A | 32 | AB | 22 | A | 26 | A | 3.3 | A | 3.4 | A | |||
| Winter Pea | 26 | ABCD | 29 | BC | 17 | B | 22 | ABCD | 3.0 | ABC | 2.9 | BC | |||
| Mean | 23 | 23 | 18 | 20 | 2.9 | 2.6 | |||||||||
| Standard Error | 6 | 6 | 4 | 4 | 0 | 0 | |||||||||
| Ash (g kg−1) | ADF (g kg−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | Short | Long | Short | Long | ||||||
| Canola | 64 | CD | 64 | EFGH | 293 | BCDEF | 328 | ABC | ||
| Forage Radish | 82 | AB | 80 | BCD | 301 | ABCDE | 325 | ABCD | ||
| Turnip | 77 | ABCD | 70 | DEFG | 277 | CDEFG | 350 | A | * | |
| Barley | 66 | BCD | 69 | DEFG | 275 | CDEFG | 304 | BCDEF | ||
| Cereal Rye | 61 | D | 59 | FGH | 243 | G | 276 | EFG | ||
| Oat | 65 | CD | 55 | GH | 242 | G | 250 | G | ||
| Triticale | 67 | BCD | 60 | FGH | 254 | FG | 286 | DEFG | ||
| Wheat | 72 | ABCD | 53 | H | 274 | DEFG | 268 | FG | ||
| Clover, Arrowleaf | 72 | ABCD | 82 | BCD | 340 | A | 307 | BCDEF | ||
| Clover, Berseem | 67 | BCD | 79 | BCDE | 319 | AB | 311 | ABCDE | ||
| Clover, Crimson | 78 | ABC | 87 | ABC | 274 | DEFG | 298 | CDEF | ||
| Clover, Red | 69 | BCD | 81 | BCD | 316 | ABC | 274 | EFG | ||
| Vetch, Common | 71 | ABCD | 98 | A | * | 313 | ABCD | 291 | CDEFG | |
| Vetch, Hairy | 87 | A | 87 | ABC | 265 | EFG | 320 | ABCD | * | |
| Vetch, Woolypod | 76 | ABCD | 92 | AB | 264 | EFG | 339 | AB | * | |
| Winter Pea | 74 | ABCD | 73 | CDEF | 268 | EFG | 271 | EFG | ||
| Mean | 72 | 74 | 282 | 300 | ||||||
| Standard Error | 8 | 8 | 18 | 19 | ||||||
| CP (g kg−1) | TDN (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | Short | Long | Short | Long | ||||||
| Canola | 175 | BCD | 131 | FGHI | * | 66 | CDEF | 63 | FGH | |
| Forage Radish | 168 | BCDE | 156 | EFG | 65 | DEF | 63 | EFGH | ||
| Turnip | 175 | BCD | 135 | FGH | * | 67 | BCD | 61 | H | * |
| Barley | 124 | G | 110 | HI | 70 | ABC | 67 | BCDE | ||
| Cereal Rye | 152 | DEFG | 123 | GHI | 73 | A | 70 | ABC | ||
| Oat | 129 | G | 101 | HI | 73 | A | 72 | A | ||
| Triticale | 140 | EFG | 110 | HI | 72 | A | 69 | ABCD | ||
| Wheat | 131 | FG | 97 | I | 70 | AB | 71 | AB | ||
| Clover, Arrowleaf | 149 | DEFG | 179 | DE | 61 | G | 64 | EFGH | ||
| Clover, Berseem | 165 | CDEF | 163 | EF | 63 | FG | 64 | EFGH | ||
| Clover, Crimson | 200 | AB | 201 | CD | 67 | BCDEF | 65 | EFG | ||
| Clover, Red | 165 | CDEF | 216 | BC | * | 63 | FG | 67 | CDEF | |
| Vetch, Common | 171 | BCDE | 210 | BCD | * | 63 | EFG | 65 | DEF | |
| Vetch, Hairy | 223 | A | 255 | A | 67 | BCD | 63 | FGH | ||
| Vetch, Woolypod | 217 | A | 237 | AB | 67 | BCD | 61 | GH | * | |
| Winter Pea | 193 | ABC | 207 | BCDEF | 67 | BCDE | 67 | BCDE | ||
| Mean | 167 | 165 | 67 | 66 | ||||||
| Standard Error | 27 | 27 | 2 | 2 | ||||||
| Short-Season | Long-Season | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | CP (g kg−1) | ADF (g kg−1) | NDF (g kg−1) | CP (g kg−1) | ADF (g kg−1) | NDF (g kg−1) | ||||||
| Canola | 175 | BCD | 293 | BCDEF | 415 | DE | 131 | FGHI | 328 | ABC | 496 | DE |
| Forage Radish | 168 | BCDE | 301 | ABCDE | 439 | CDE | 156 | EFG | 325 | ABCD | 484 | CDE |
| Turnip | 175 | BCD | 277 | CDEFG | 419 | BCDE | 135 | FGH | 350 | A | 509 | BCDE |
| Barley | 124 | G | 275 | CDEFG | 477 | A | 110 | HI | 304 | BCDEF | 570 | A |
| Cereal Rye | 152 | DEFG | 243 | G | 475 | AB | 123 | GHI | 276 | EFG | 565 | AB |
| Oat | 129 | G | 242 | G | 437 | DE | 101 | HI | 250 | G | 462 | DE |
| Triticale | 140 | EFG | 254 | FG | 472 | ABC | 110 | HI | 286 | DEFG | 557 | ABC |
| Wheat | 131 | FG | 274 | DEFG | 487 | ABCD | 97 | I | 268 | FG | 500 | ABCD |
| Clover, Arrowleaf | 149 | DEFG | 340 | A | 475 | DE | 179 | DE | 307 | BCDEF | 430 | DE |
| Clover, Berseem | 165 | CDEF | 319 | AB | 453 | DE | 163 | EF | 311 | ABCDE | 451 | DE |
| Clover, Crimson | 200 | AB | 274 | DEFG | 379 | F | 201 | CD | 298 | CDEF | 407 | F |
| Clover, Red | 165 | CDEF | 316 | ABC | 436 | EF | 216 | BC | 274 | EFG | 390 | EF |
| Vetch, Common | 171 | BCDE | 313 | ABCD | 440 | EF | 210 | BCD | 291 | CDEFG | 407 | EF |
| Vetch, Hairy | 223 | A | 265 | EFG | 392 | EF | 255 | A | 320 | ABCD | 455 | EF |
| Vetch, Woolypod | 217 | A | 264 | EFG | 387 | EF | 237 | AB | 339 | AB | 464 | EF |
| Winter Pea | 193 | ABC | 268 | EFG | 422 | EF | 207 | BCDEF | 271 | EFG | 432 | EF |
| Est. Nitrogen Release over 2 Weeks kg ha−1 | Est. Nitrogen Release over 4 Weeks kg ha−1 | Est. Nitrogen Release over 12 Weeks kg ha−1 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Short | Long | Short | Long | Short | Long | |||||||||
| Canola | 4 | A | 15 | B | * | 7 | A | 24 | BC | 10 | A | 35 | BC | * | |
| Forage Radish | 2 | A | 4 | DE | 4 | A | 5 | EF | 6 | A | 7 | G | |||
| Turnip | 3 | A | 7 | BCDE | 6 | A | 12 | CDEF | 9 | A | 17 | DEFG | |||
| Barley | 3 | A | 7 | CDE | 4 | A | 11 | DEF | 7 | A | 16 | DEFG | |||
| Cereal Rye | 4 | A | 8 | BCDE | 6 | A | 13 | CDEF | 9 | A | 20 | CDEFG | |||
| Oat | 3 | A | 0 | E | 5 | A | 2 | F | 9 | A | 7 | G | |||
| Triticale | 3 | A | 3 | E | 5 | A | 6 | EF | 9 | A | 11 | FG | |||
| Wheat | 3 | A | 3 | E | 5 | A | 6 | EF | 9 | A | 13 | EFG | |||
| Clover, Arrowleaf | 1 | A | 12 | BCD | 2 | A | 18 | CDE | 3 | A | 25 | CDEF | |||
| Clover, Berseem | 2 | A | 2 | E | 4 | A | 4 | F | 6 | A | 5 | G | |||
| Clover, Crimson | 6 | A | 29 | A | * | 9 | A | 45 | A | * | 13 | A | 63 | A | * |
| Clover, Red | 2 | A | 15 | BC | * | 4 | A | 21 | CD | 7 | A | 30 | CD | ||
| Vetch, Common | 3 | A | 14 | BC | 5 | A | 21 | CD | 7 | A | 29 | CDE | |||
| Vetch, Hairy | 8 | A | 25 | A | * | 13 | A | 39 | A | * | 18 | A | 52 | AB | * |
| Vetch, Woolypod | 7 | A | 26 | A | * | 10 | A | 39 | A | * | 15 | A | 52 | AB | * |
| Winter Pea | 6 | A | 24 | A | * | 10 | A | 37 | AB | * | 14 | A | 50 | AB | * |
| Mean | 4 | 12 | 6 | 19 | 9 | 27 | |||||||||
| Standard Error | 4 | 4 | 6 | 6 | 8 | 8 | |||||||||
| Cash Crop | Soil | ||||||
|---|---|---|---|---|---|---|---|
| Effect | Yield | Protein | Oil | Moisture | pH | OM | WEC |
| Year | 0.97 | 0.14 | 0.51 | 0.28 | 0.06 | 0.77 | 0.05 |
| Season | 0.04 | <0.001 | <0.001 | 0.007 | 0.11 | <0.001 | 0.14 |
| Year × Season | 0.77 | 0.75 | 0.23 | 0.45 | 0.08 | <0.001 | 0.16 |
| Harvest | 0.17 | 0.12 | 0.43 | 0.45 | 0.27 | 0.45 | 0.82 |
| Year × Harvest | 0.72 | 0.93 | 0.52 | 0.73 | 0.61 | 0.41 | 0.96 |
| Season × Harvest | 0.61 | <0.001 | 0.38 | 0.69 | 0.24 | 0.69 | 0.50 |
| Year × Season × Harvest | 0.33 | 0.15 | 0.70 | 0.62 | 0.85 | 0.65 | 0.58 |
| Species | 0.81 | 0.02 | 0.59 | 0.03 | 0.01 | 0.48 | 0.86 |
| Year × Species | 0.56 | 0.75 | 0.52 | 0.99 | 0.98 | 0.77 | 0.84 |
| Season × Species | 0.94 | 0.32 | 0.39 | 0.82 | 0.70 | 1.00 | 0.92 |
| Year × Season × Species | 0.19 | 0.47 | 0.81 | 0.17 | 0.08 | 0.76 | 0.91 |
| Harvest × Species | 0.83 | 0.27 | 0.41 | 0.93 | 0.48 | 0.10 | 0.40 |
| Year × Harvest × Species | 0.93 | 0.59 | 0.72 | 0.93 | 0.98 | 0.98 | 0.37 |
| Season × Harvest × Species | 0.78 | 0.34 | 0.47 | 0.98 | 0.96 | 1.00 | 0.93 |
| Year x Season × Harvest × Species | 0.92 | 0.12 | 0.74 | 0.50 | 0.66 | <0.01 | 0.36 |
| Short (Corn) | Long (Soy) | Mean | SE | |
| Yield (kg ha−1) | 11,519 | 3393 | 7456 | 1451 |
| Oil (g kg−1) | 35 | 204 | 120 | 2 |
| Protein (g kg−1) | 78 | 333 | 206 | 3 |
| Moist (%) | 10.6 | 12.5 | 11.6 | 3.5 |
| NO3 (mg kg−1) | 26 | 8 | 17 | 5 |
| 2018 | 2019 | Mean | SE | |
| WEC (mg kg−1) | 145 | 106 | 126 | 10 |
| K (mg kg−1) | 129 | 76 | 103 | 7 |
| Mg (mg kg−1) | 112 | 78 | 95 | 6 |
| NO3-N (mg kg−1) | 24 | 10 | 17 | 5 |
| Soil NO3-N (mg kg−1) | Soil Moisture (%) | Cash Crop Protein (g kg−1) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Species | Short (Corn) | Long (Soy) | MS | Short (Corn) | Long (Soy) | MS | Short (Corn) | Long (Soy) | MS |
| Canola | 25 | 7 | BCD | 10.7 | 12.9 | ABCD | 76 | 333 | CDE |
| Forage Radish | 26 | 8 | ABCD | 10.6 | 12.3 | ABCDEF | 79 | 332 | ABCDE |
| Turnip | 27 | 6 | CD | 10.8 | 12.8 | ABCDE | 76 | 331 | E |
| Barley | 28 | 9 | CD | 10.0 | 12.6 | BCDEF | 77 | 334 | ABC |
| Cereal Rye | 24 | 6 | CD | 10.3 | 12.1 | DEF | 79 | 332 | ABCD |
| Oat | 26 | 9 | D | 10.9 | 12.9 | ABC | 79 | 334 | A |
| Triticale | 25 | 8 | D | 10.4 | 12.9 | ABCDE | 78 | 333 | ABCD |
| Wheat | 27 | 8 | CD | 10.3 | 12.0 | CDEF | 78 | 334 | AB |
| Clover, Arrowleaf | 27 | 8 | ABC | 10.4 | 12.0 | BCDEF | 78 | 333 | ABCD |
| Clover, Berseem | 23 | 5 | CD | 11.1 | 13.0 | A | 76 | 332 | DE |
| Clover, Crimson | 25 | 5 | ABC | 10.8 | 12.9 | ABCD | 77 | 333 | CDE |
| Clover, Red | 23 | 7 | ABCD | 10.6 | 11.7 | EF | 78 | 333 | ABCD |
| Vetch, Common | 24 | 8 | ABCD | 10.4 | 11.5 | F | 78 | 334 | AB |
| Vetch, Hairy | 28 | 11 | AB | 10.3 | 12.4 | BCDEF | 77 | 335 | ABCD |
| Vetch, Woolypod | 26 | 12 | A | 10.9 | 12.8 | AB | 77 | 333 | ABCDE |
| Winter Pea | 27 | 6 | AB | 10.4 | 12.9 | ABCDE | 78 | 332 | BCDE |
| No Cover | 29 | 11 | ABC | 10.4 | 12.5 | ABCDEF | 79 | 334 | ABC |
| Mean | 26 | 8 | 10.6 | 12.5 | 78 | 333 | |||
| SE | 5 | 5 | 3.5 | 3.5 | 3 | 3 | |||
| WEN (mg kg−1) | Cash Crop Protein (g kg−1) | Fe (mg kg−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Harvest | Est. | MS | Short (Corn) | Long (Soy) | 2018 | 2019 | ||||
| Forage | 44 | A | 78 | A | 332 | B | 210 | A | 128 | A |
| Non-Forage | 43 | B | 77 | A | 334 | A | 197 | B | 127 | A |
| Mean | 43 | 78 | 333 | 203 | 127 | |||||
| SE | 8 | 3 | 3 | 43 | 43 | |||||
| Effect | WEN | NO3-N | Fe | K | Mg | Na | P |
|---|---|---|---|---|---|---|---|
| Year | 0.48 | 0.047 | <0.001 | 0.007 | <0.001 | 0.01 | 0.18 |
| Season | 0.08 | 0.02 | 0.07 | 0.54 | 0.86 | 0.17 | 0.91 |
| Year × Season | 0.85 | 0.27 | 0.10 | 0.56 | 0.91 | 0.39 | 0.18 |
| Harvest | 0.04 | 0.52 | 0.02 | 0.42 | 0.71 | 0.20 | 0.21 |
| Year × Harvest | 0.66 | 0.47 | 0.03 | 0.84 | 0.37 | 0.54 | 0.87 |
| Season × Harvest | 0.59 | 0.21 | 0.70 | 0.22 | 0.85 | 0.28 | 0.64 |
| Year × Season × Harvest | 0.18 | 0.16 | 0.83 | 0.57 | 0.58 | 0.07 | 0.35 |
| Species | 0.03 | <0.001 | 0.23 | 0.88 | 0.25 | 0.52 | 0.09 |
| Year × Species | 0.60 | 0.46 | 0.49 | 0.98 | 0.94 | 0.69 | 0.99 |
| Season × Species | 0.97 | 0.91 | 0.83 | 0.94 | 0.97 | 0.90 | 0.96 |
| Year × Season × Species | 0.93 | 0.90 | 0.63 | 0.69 | 0.92 | 0.61 | 0.03 |
| Harvest × Species | 0.82 | 0.80 | 0.10 | 0.33 | 0.79 | 0.60 | 0.42 |
| Year × Harvest × Species | 0.82 | 0.71 | 0.08 | 0.98 | 0.97 | 0.04 | 0.95 |
| Season × Harvest × Species | 0.29 | 0.44 | 0.72 | 0.97 | 1.00 | 0.58 | 0.99 |
| Year × Season × Harvest × Species | 0.26 | 0.48 | 0.83 | 0.12 | 0.45 | 1.00 | 0.006 |
| Species | WEN (mg kg−1) | Soil pH | ||
|---|---|---|---|---|
| Canola | 44 | BCDE | 5.8 | AB |
| Forage Radish | 43 | ABCDE | 5.8 | ABCD |
| Turnip | 42 | BCDE | 5.8 | ABC |
| Barley | 42 | ABCDE | 5.8 | ABCD |
| Cereal Rye | 42 | CDE | 5.8 | ABCD |
| Oat | 40 | ABCD | 5.7 | DE |
| Triticale | 41 | CDE | 5.7 | DE |
| Wheat | 42 | BCDE | 5.8 | BCDE |
| Clover, Arrowleaf | 45 | ABCDE | 5.8 | BCDE |
| Clover, Berseem | 42 | E | 5.9 | A |
| Clover, Crimson | 46 | DE | 5.8 | AB |
| Clover, Red | 42 | CDE | 5.8 | ABCD |
| Vetch, Common | 43 | BCDE | 5.8 | ABCDE |
| Vetch, Hairy | 46 | ABC | 5.7 | DE |
| Vetch, Woolypod | 47 | AB | 5.7 | E |
| Winter Pea | 48 | CDE | 5.8 | ABC |
| No Cover | 45 | A | 5.7 | CDE |
| Mean | 43 | 5.8 | ||
| SE | 8 | 0.2 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bracey, W.M.; Sykes, V.R.; Yin, X.; Bates, G.E.; Butler, D.M.; McIntosh, D.W.; Willette, A.R. Forage Yield, Quality, and Impact on Subsequent Cash Crop of Cover Crops in an Integrated Forage/Row Crop System. Agronomy 2022, 12, 1214. https://doi.org/10.3390/agronomy12051214
Bracey WM, Sykes VR, Yin X, Bates GE, Butler DM, McIntosh DW, Willette AR. Forage Yield, Quality, and Impact on Subsequent Cash Crop of Cover Crops in an Integrated Forage/Row Crop System. Agronomy. 2022; 12(5):1214. https://doi.org/10.3390/agronomy12051214
Chicago/Turabian StyleBracey, Weston M., Virginia R. Sykes, Xinhua Yin, Gary E. Bates, David M. Butler, David W. McIntosh, and Allison R. Willette. 2022. "Forage Yield, Quality, and Impact on Subsequent Cash Crop of Cover Crops in an Integrated Forage/Row Crop System" Agronomy 12, no. 5: 1214. https://doi.org/10.3390/agronomy12051214
APA StyleBracey, W. M., Sykes, V. R., Yin, X., Bates, G. E., Butler, D. M., McIntosh, D. W., & Willette, A. R. (2022). Forage Yield, Quality, and Impact on Subsequent Cash Crop of Cover Crops in an Integrated Forage/Row Crop System. Agronomy, 12(5), 1214. https://doi.org/10.3390/agronomy12051214