
Current Status of Herbicide Resistance in the Iberian Peninsula: Future Trends and Challenges

 ,
,  , ,
, ,
Abstract
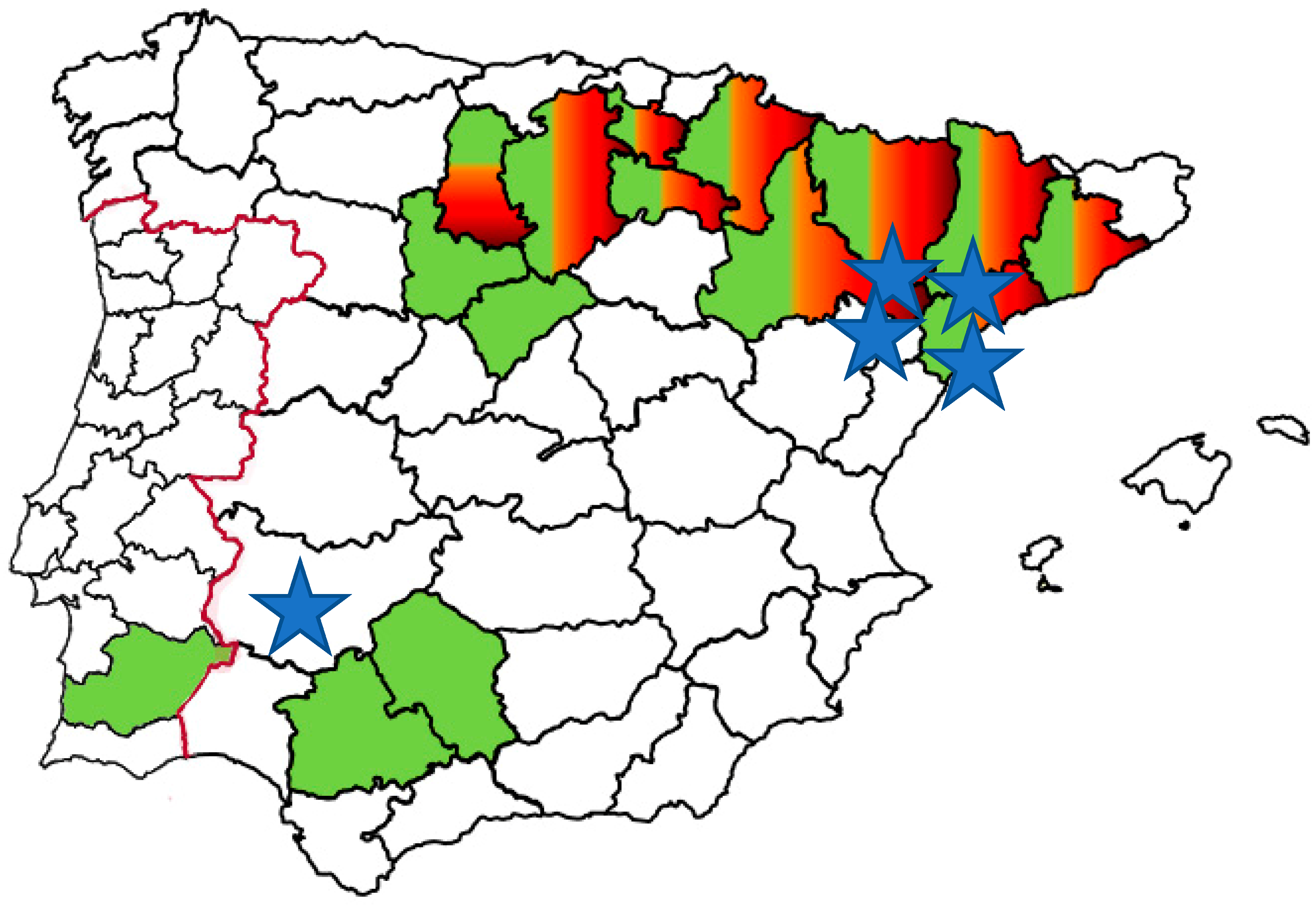
1. Introduction
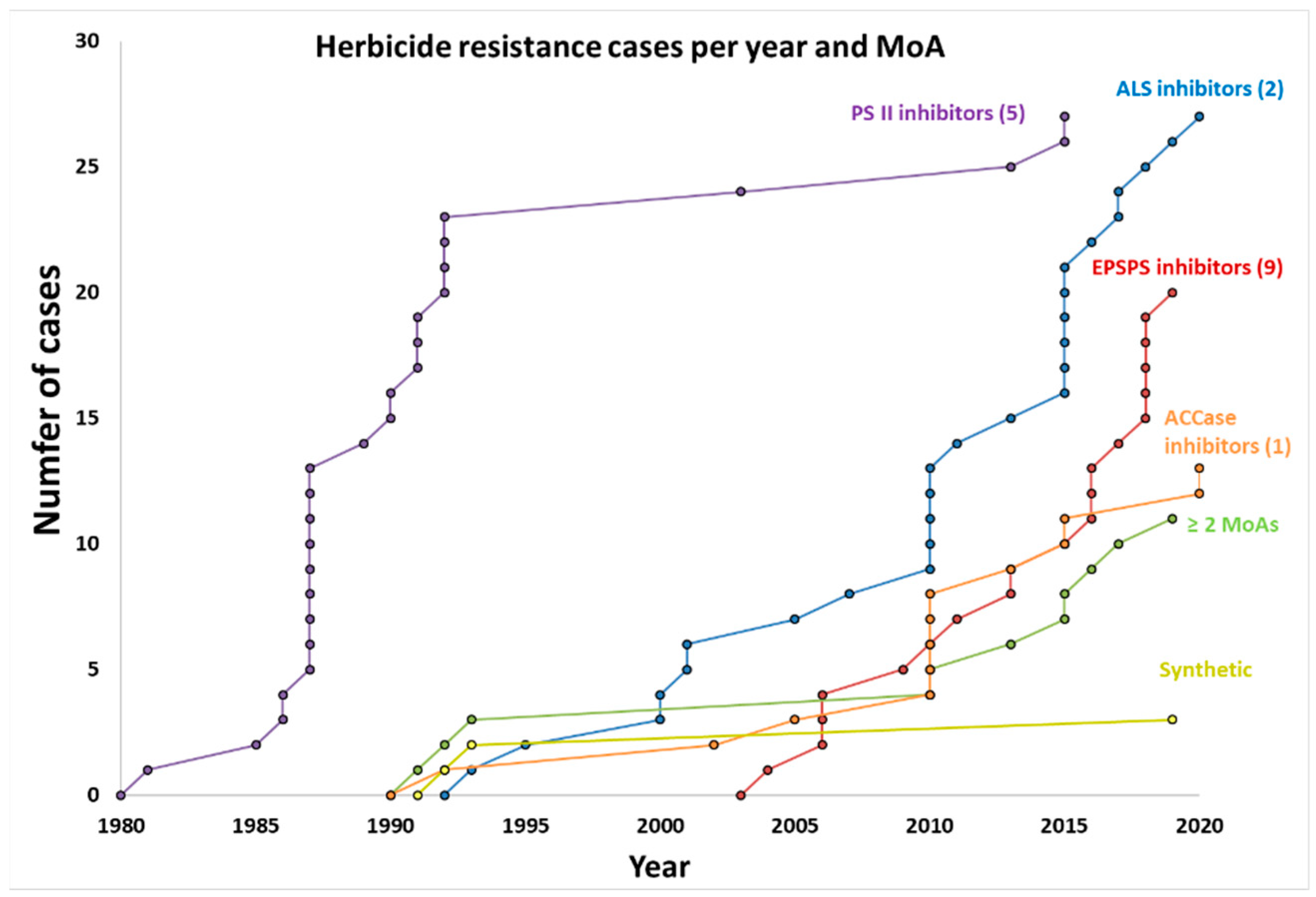
2. Resistance by Modes of Action
2.1. Resistance to ACCase Inhibitors (Group 1)
2.2. Resistance to ALS Inhibitors (Group 2)
2.3. Resistance to PSII Inhibitors (Groups 5 and 6)
2.4. Resistance to EPSPS Inhibitors (Group 9)
2.5. Resistance to Other Modes of Action
3. Multiple-Herbicide-Resistant Weeds
4. Main Herbicide-Resistant Weed Species
4.1. Lolium spp.
4.2. Echinochloa spp.
4.3. Conyza spp.
4.4. Papaver rhoeas
4.5. Amaranthus palmeri
5. Future Trends and Challenges
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Shaner, D.L.; Beckie, H.J. The future for weed control and technology. Pest Manag. Sci. 2014, 70, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Baucom, R.S. The remarkable repeated evolution of herbicide resistance. Am. J. Bot. 2016, 103, 181–183. [Google Scholar] [CrossRef] [PubMed]
- Torra, J.; Osuna, M.D.; Merotto, A.; Vila-Aiub, M. Multiple herbicide-resistant Weeds and non-target site resistance mecha-nisms: A global challenge for food production. Front. Plant Sci. 2021, 12, 763212. [Google Scholar] [CrossRef] [PubMed]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: https://www.weedscience.org (accessed on 12 December 2021).
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- European Commission (EC). Eurostat. 2022. Available online: https://ec.europa.eu/eurostat/web/products-eurostat-news (accessed on 4 April 2022).
- Ministerio de Agricultura, Pesca y Alimentación (MAPA). Estadísticas 2022. Agrarias. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/ (accessed on 4 April 2022).
- Calha, I.M.; Rocha, F.; Ruiz-Santaella, J.P.; Cruz-Hipólito, H.E. Two decades of herbicide resistance in the iberian peninsula. J. Plant Dis. Prot. Suppl. 2008, 21, 79–84. [Google Scholar]
- Calha, I.M.; Machado, C.; Rocha, F. Resistance of Alisma plantago aquatica to sulfonylurea herbicides in Portuguese rice fields. In Biology, Ecology and Management of Aquatic Plants; Springer: Dordrecht, The Netherlands, 1999; pp. 289–293. [Google Scholar] [CrossRef]
- Calha, I.M.; Osuna, M.D.; Serra, C.; Moreira, I.; De Prado, R.; Rocha, F. Mechanism of resistance to bensulfuron-methyl in Alisma plantago-aquatica biotypes from Portuguese rice paddy fields. Weed Res. 2007, 47, 231–240. [Google Scholar] [CrossRef]
- Fernández, P.; Alcántara, R.; Osuna, M.D.; Vila-Aiub, M.M.; De Prado, R. Forward selection for multiple resistance across the non-selective glyphosate, glufosinate and oxyfluorfen herbicides inLoliumweed species. Pest Manag. Sci. 2016, 73, 936–944. [Google Scholar] [CrossRef]
- Vázquez-García, J.G.; Rojano-Delgado, A.M.; Alcántara-de la Cruz, R.; Torra, J.; Dellaferrera, I.; Portugal, J.; De Prado, R. Dis-tribution of glyphosate-resistance in Echinochloa crus-galli across agriculture areas in the Iberian Peninsula. Front. Plant Sci. 2021, 12, 617040. [Google Scholar] [CrossRef]
- De Prado, R.; Romera, E.; Menéndez, J. Atrazine detoxification in Panicum dichotomiflorum and target site Polygonum lapathi-folium. Pestic. Biochem. Physiol. 1995, 52, 1–11. [Google Scholar] [CrossRef]
- Prado, R.; Dominguez, C.; Tena, M. Triazine resistance in biotypes of Solanum nigrum and four Amaranthus species found in Spain. Weed Res. 1993, 33, 17–24. [Google Scholar] [CrossRef]
- Giménez-Espinosa, R.; Romera, E.; Tena, M.; De Prado, R. Fate of atrazine in treated and pristine accessions of three Setaria species. Pestic. Biochem. Physiol. 1996, 56, 196–207. [Google Scholar] [CrossRef]
- De Prado, R.; Lopez-Martinez, N.; Gonzalez-Gutierres, J. Herbicide-resistant weeds in Europe: Agricultural, physiological and biochemical aspects. In Weed and Crop Resistance to Herbicides; De Prado, R., Jorrin, J., Garcia-Torres, L., Eds.; Kluwer: Dordrecht, The Netherlands, 1997; pp. 17–28. [Google Scholar]
- De Prado, R.; Sanchez, M.; Jorrin, J.; Dominguez, C. Negative cross-resistance to bentazone and pyridate in atrazine-resistant Amaranthus cruentus and Amaranthus hybridus biotypes. Pestic. Sci. 1992, 35, 131–136. [Google Scholar] [CrossRef]
- Menendez, J.; Bastida, F.; De Prado, R. Resistance to chlortoluron in a downy brome (Bromus tectorum) biotype. Weed Sci. 2006, 54, 237–245. [Google Scholar] [CrossRef]
- Menendez, J.; Gonzalez-Gutierrez, J.; De Prado, R. Characterisation of a triazine-resistant biotype of Bromus tectorum found in Spain. Weed Res. 2007, 47, 113–121. [Google Scholar] [CrossRef]
- Menendez, J.; Jorrin, J.; Romera, E.; De Prado, R. Resistance to chlorotoluron of a slender foxtail (Alopecurus myosuroides) bio-type. Weed Sci. 1994, 42, 340–344. [Google Scholar] [CrossRef]
- Menendez, J.; De Prado, R. Diclofop-methyl cross-resistance in a chlorotoluron-resistant biotype of Alopecurus myosuroides. Pestic. Biochem. Physiol. 1996, 56, 123–133. [Google Scholar] [CrossRef]
- De Prado, R.; De Prado, J.L.; Menendez, J. Resistance to Substituted Urea Herbicides in Lolium rigidum Biotypes. Pestic. Biochem. Physiol. 1997, 57, 126–136. [Google Scholar] [CrossRef]
- De Prado, J.L.; Osuna, M.D.; De Prado, R. Cross resistance to accase herbicide in Lolium rigidum. Commun. Agric. Appl. Biol. Sci. 2004, 69, 97–102. [Google Scholar]
- Lopez-Martinez, N.; Marshall, G.; De Prado, R. Resistance of barnyardgrass (Echinochloa crus-galli) to atrazine and quinclorac. Pestic. Sci. 1997, 51, 171–175. [Google Scholar] [CrossRef]
- Duran-Prado, M.; Osuna, M.D.; De Prado, R.; Franco, A.R. Molecular basis of resistance to sulfonylureas in Papaver rhoeas. Pestic. Biochem. Physiol. 2004, 79, 10–17. [Google Scholar] [CrossRef]
- Ruiz-Santaella, J.P.; Bakkaliu, Y.; Osuna, M.D.; De Prado, R. Evaluation of resistance in Cyperus difformis populations to ALS inhibiting herbicides. Commun. Agric. Appl. Biol. Sci. 2004, 69, 91–96. [Google Scholar] [PubMed]
- Osuna, M.D.; De Prado, R. Conyza albida: A new biotype with ALS inhibitor resistance. Weed Res. 2003, 43, 221–226. [Google Scholar] [CrossRef]
- Osuna, M.D.; Fischer, A.J.; De Prado, R. Herbicide resistance in Aster squamatus conferred by a less sensitive form of aceto-lactate synthase. Pest Manag. Sci. 2003, 59, 1210–1216. [Google Scholar] [CrossRef] [PubMed]
- De Prado, R.; Osuna, M.D.; Fischer, A.J. Resistance to ACCase inhibitor herbicides in a green foxtail (Setaria viridis) biotype in Europe. Weed Sci. 2004, 52, 506–512. [Google Scholar] [CrossRef]
- Urbano, J.; Borrego, A.; Torres, V.; Leon, J.; Jimenez, C.; Dinelli, G.; Barnes, J. Glyphosate-resistant Hairy Fleabane (Conyza bonar-iensis) in Spain. Weed Technol. 2007, 21, 396–401. [Google Scholar] [CrossRef]
- González-Torralva, F.; Rojano-Delgado, A.M.; de Castro, M.D.L.; Mülleder, N.; De Prado, R. Two non-target mechanisms are involved in glyphosate-resistant horseweed (Conyza canadensis L. Cronq.) biotypes. J. Plant Physiol. 2012, 169, 1673–1679. [Google Scholar] [CrossRef]
- Cruz-Hipolito, H.; Rosario, J.; Ioli, G.; Osuna, M.D.; Smeda, R.J.; Torralva, F.G.; De Prado, R. Resistance Mechanism to Tribenuron-Methyl in White Mustard (Sinapis alba) from Southern Spain. Weed Sci. 2013, 61, 341–347. [Google Scholar] [CrossRef]
- Amaro-Blanco, I.; Fernández-Moreno, P.T.; Osuna-Ruiz, M.D.; Bastida, F.; De Prado, R. Mechanisms of glyphosate resistance and response to alternative herbicide-based management in populations of the three Conyza species introduced in southern Spain. Pest Manag. Sci. 2018, 74, 1925–1937. [Google Scholar] [CrossRef]
- de Barreda, D.G.; Pardo, G.; Osca, J.; Catala-Forner, M.; Consola, S.; Garnica, I.; López-Martínez, N.; Palmerín, J.; Osuna, M. An Overview of Rice Cultivation in Spain and the Management of Herbicide-Resistant Weeds. Agronomy 2021, 11, 1095. [Google Scholar] [CrossRef]
- Amaro, I.; Romano, Y.; Palmerin, J.A.; Quiles, J.M.; Osuna, M.D. Resistencia a herbicidas inhibidores de la ALS y ACCasa en poblaciones de Leptochloa spp. en arrozales de Extremadura. In Proceedings of the XVII Congreso de la Sociedad Española de Malherbología, Vigo, Spain, 8–10 October 2019. [Google Scholar]
- Amaro-Blanco, I.; Romano, Y.P.; Jose, A.; Gordo, R.; Palma-Bautista, C.; De Prado, R.; Osuna, M.D. Different Mutations Providing Target Site Resistance to ALS and ACCase Inhibiting Herbicides in Echinochloa spp. from Rice Fields. Agriculture 2021, 11, 382. [Google Scholar] [CrossRef]
- Loureiro, I.; Escorial, C.; Plaza, E.H.; Gonzalez-Andujar, J.L.; Chueca, M.C. Current status in herbicide resistance in Lolium rigidum in winter cereal fields in Spain: Evolution of resistance 12 years after. Crop Prot. 2017, 102, 10–18. [Google Scholar] [CrossRef]
- Travlos, I.S.; Montull, J.M.; Kukorelli, G.; Malidza, G.; Dogan, M.N.; Cheimona, N.; Antonopoulos, N.; Kanatas, P.J.; Zannopoulos, S.; Peteinatos, G. Key Aspects on the Biology, Ecology and Impacts of Johnsongrass [Sorghum halepense (L.) Pers] and the Role of Glyphosate and Non-Chemical Alternative Practices for the Management of This Weed in Europe. Agronomy 2019, 9, 717. [Google Scholar] [CrossRef]
- Torra, J.; Montull, J.M.; Royo-Esnal, A.; Taberner, A.; Salas, M.L. Resistance to ALS inhibiting herbicides in a Spanish Echi-nochloa cruss-galli population from a corn field. In Proceedings of the Conference: Resistance’19, Poster 33, Rothamsted, UK, 18 October 2019. [Google Scholar]
- Vazquez-Garcia, J.G.; Palma-Bautista, C.; Rojano-Delgado, A.M.; De Prado, R.; Menendez, J. The first case of glyphosate re-sistance in Johnsongrass (Sorghum halepense (L.) Pers.) in Europe. Plants 2020, 9, 313. [Google Scholar] [CrossRef] [PubMed]
- Torra, J.; Montull, J.M.; Taberner, A.; Onkokesung, N.; Boonham, N.; Edwards, R. Target-Site and Non-target-Site Resistance Mechanisms Confer Multiple and Cross- Resistance to ALS and ACCase Inhibiting Herbicides in Lolium rigidum from Spain. Front. Plant Sci. 2021, 12, 138. [Google Scholar] [CrossRef] [PubMed]
- Torra, J.; Royo-Esnal, A.; Romano, Y.; Osuna, M.D.; León, R.G.; Recasens, J. Amaranthus palmeri a New Invasive Weed in Spain with Herbicide Resistant Biotypes. Agronomy 2020, 10, 993. [Google Scholar] [CrossRef]
- Amaro-Blanco, I.; Osuna, M.D.; Romano, Y.; Roldán-Gómez, R.; Palma-Bautista, C.; Portugal, J.; De Prado, R. Selection for glyphosate resistance in Conyza spp. occurring in the railway network of southern Spain. Can. J. Plant Sci. 2019, 99, 413–419. [Google Scholar] [CrossRef]
- Mora, D.A.; Cheimona, N.; Palma-Bautista, C.; Rojano-Delgado, A.M.; Osuna-Ruiz, M.D.; de la Cruz, R.A.; De Prado, R. Physiological, biochemical and molecular bases of resistance to tribenuron-methyl and glyphosate in Conyza canadensis from olive groves in southern Spain. Plant Physiol. Biochem. 2019, 144, 14–21. [Google Scholar] [CrossRef]
- Pardo, G.; Marí, A.I.; Aibar, J.; Vilaplana, L.; Cirujeda, A. Bastard Cabbage (Rapistrum rugosum L.) Resistance to Tribenu-ron-Methyl and Iodosulfuron-Methyl-Sodium in Spain and Alternative Herbicides for Its Control. Agronomy 2019, 9, 492. [Google Scholar] [CrossRef]
- Vázquez-García, J.G.; Castro, P.; Torra, J.; Alcántara-De la Cruz, R.; De Prado, R. Resistance evolution to EPSPS inhibiting herbicides in false barley (Hordeum murinum) harvested in Southern Spain. Agronomy 2020, 10, 992. [Google Scholar] [CrossRef]
- Vázquez-García, J.G.; Castro, P.; Cruz-Hipólito, H.E.; Millan, T.; Palma-Bautista, C.; De Prado, R. Glyphosate Resistance Confir-mation and Field Management of Red Brome (Bromus rubens L.) in Perennial Crops Grown in Southern Spain. Agronomy 2021, 11, 535. [Google Scholar] [CrossRef]
- Palma-Bautista, C.; Vázquez-García, J.G.; Domínguez-Valenzuela, J.A.; Mendes, K.F.; Alcántara de la Cruz, R.; Torra, J.; De Prado, R. Non-Target-Site Resistance mechanisms endow multiple herbicide resistance to five mechanisms of action in Con-yza bonariensis. J. Agric. Food Chem. 2021, 69, 14792–14801. [Google Scholar] [CrossRef] [PubMed]
- Herbicide Resistance Action Committee (HRAC). HRAC Mode of Action Classification 2020 Map. 2022. Available online: https://hracglobal.com/tools/hrac-mode-of-action-classification-2020-map (accessed on 5 April 2022).
- Menéndez, J.; De Prado, R. Characterization of two acetyl-CoA carboxylase isoforms in diclofop-methyl-resistant and -susceptible biotypes of Alopecurus myosuroides. Pestic. Biochem. Physiol. 1999, 65, 82–89. [Google Scholar] [CrossRef]
- De Prado, J.L.; Osuna, M.D.; Heredia, A.A.; De Prado, R. Lolium rigidum, a Pool of Resistance Mechanisms to ACCase Inhibitor Herbicides. J. Agric. Food Chem. 2005, 53, 2185–2191. [Google Scholar] [CrossRef]
- Rey-Caballero, J.; Menéndez, J.; Giné-Bordonaba, J.; Salas, M.; Alcántara, R.; Torra, J. Unravelling the resistance mechanisms to 2,4-D (2,4-dichlorophenoxyacetic acid) in corn poppy (Papaver rhoeas). Pestic. Biochem. Physiol. 2016, 133, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Rey-Caballero, J.; Menéndez, J.; Osuna, M.D.; Salas, M.; Torra, J. Target-site and non-target-site resistance mechanisms to ALS inhibiting herbicides in Papaver rhoeas. Pestic. Biochem. Physiol. 2017, 138, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Busi, R.; Vidotto, F.; Fischer, A.J.; Osuna, M.D.; De Prado, R.; Ferrero, A. Patterns of resistance to ALS herbicides in small-flower umbrella sedge (Cyperus difformis) and ricefield bulrush (Schoenoplectus mucronatus). Weed Technol. 2006, 20, 1004–1014. [Google Scholar] [CrossRef]
- Torra, J.; Rojano-Delgado, A.M.; Menéndez, J.; Salas, M.; De Prado, R. Cytochrome P450 metabolism-based herbicide resistance to ALS inhibitors and 2,4-D in Papaver rhoeas. Plant Physiol. Biochem. 2021, 160, 51–61. [Google Scholar] [CrossRef] [PubMed]
- De Prado, R.; Romera, E.; Jorrin, J. Effects of chloroacetamides and phytosynthesis-inhibiting herbicides on growth and photo-synthesis in sunflower (Helianthus animus L.) and Amaranthus hybridus L. Weed Res. 1993, 33, 369–374. [Google Scholar] [CrossRef]
- Dominguez, C.; Tena, M.; De Prado, R. Physiological and biochemical characteristics in triazine resistant biotypes of four Ama-ranthus species. J. Plant Physiol. 1994, 14, 632–636. [Google Scholar] [CrossRef]
- Dominguez, C.; Tena, M.; De Prado, R. Photosynthetic activity, growth and productivity in triazine-resistant biotype of Sola-num nigrum. Plant Physiol. Biochem. 1994, 32, 627–632. [Google Scholar]
- Calha, I.M.; Matias, A.; Alves, A.C.; Mora, L.; Rocha, F. Quick-Tests for Detection Weed Resistance to Sulfonylurea and Triazine Herbicides–Preliminary Results; EWRS: Athens, Greece, 2005. [Google Scholar]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Moreno, P.T.; Travlos, I.; Brants, I.; De Prado, R. Differentlevels of glyphosate-resistant Lolium rigidum L. among major crops in southern Spain and France. Sci. Rep. 2017, 7, 13116. [Google Scholar] [CrossRef] [PubMed]
- Palma-Bautista, C.; Rojano-Delgado, A.M.; Dellaferrera, I.; Rosario, J.M.; Vigna, M.R.; Torra, J.; De Prado, R. Resistance Mechanisms to 2,4-D in Six Different Dicotyledonous Weeds around the World. Agronomy 2020, 10, 566. [Google Scholar] [CrossRef]
- Busi, R.; Goggin, D.E.; Heap, I.M.; Horak, M.J.; Jugulam, M.; Masters, R.A.; Napier, R.M.; Riar, D.S.; Satchivi, N.M.; Torra, J.; et al. Weed resistance to synthetic auxin herbicides. Pest Manag. Sci. 2018, 74, 2265–2276. [Google Scholar] [CrossRef] [PubMed]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.G.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of evolved herbicide resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef]
- Menéndez, J.; De Prado, R. Detoxification of chlorotoluron in a chlorotoluron-resistant biotype of Alopecurus myosuroides. Comparison between cell cultures and whole plants. Physiol. Plant 1997, 99, 97–104. [Google Scholar] [CrossRef]
- Pan, L.; Yu, Q.; Han, H.; Mao, L.; Nyporko, A.; Fan, L.; Bai, L.; Powles, S.B. Aldo-keto Reductase Metabolizes Glyphosate and Con-fers Glyphosate Resistance in Echinochloa colona. Plant Physiol. 2019, 181, 1519–1534. [Google Scholar] [CrossRef]
- Atanackovic, V.; Juárez-Escario, A.; Recasens, J.; Torra, J. A survey of Lolium rigidum populations in citrus orchards: Factors explaining infestation levels. Weed Biol. Manag. 2015, 15, 122–131. [Google Scholar] [CrossRef]
- Mendes, A.; Portugal, J.; Calha, I.; Andrade, E. Potential basis of glyphosate resistance in Lolium perenne from Douro vineyards. In Proceedings of the Actas del XVII Congreso SEMh, Sociedad Española de Malherbología, Vigo, Spain, 8–10 October 2019. [Google Scholar]
- Comité Prevención Resistencia Herbicidas (CPRH). Working of the Spanish Weed Science Society; Spanish Weed Science Society: Vigo, Spain, 2022. [Google Scholar]
- Loureiro, I.; Escorial, M.-C.; Chueca, M.-C. Pollen-Mediated Movement of Herbicide Resistance Genes in Lolium rigidum. PLoS ONE 2016, 11, e0157892. [Google Scholar] [CrossRef]
- Escorial, M.C.; Hornos, Z.; Chueca, M.C. Loureiro Aptitud biológica de un biotipo de Lolium rigidum resistente a herbicidas inhibidores de la enzima acetolactato sintetasa. In Proceedings of the XVII Congreso de la Sociedad Española de Malherbología, Vigo, Spain, 8–10 October 2019. [Google Scholar]
- Recasens, J.; Caimons, O.; Torra, J.; Taberner, A. Variation in seed germination and early growth between and within acetolac-tate synthase herbicide resistant and susceptible Lolium rigidum accessions. Seed Sci. Technol. 2007, 35, 32–47. [Google Scholar] [CrossRef]
- Piasecki, C.; Yang, Y.; Benemann, D.P.; Kremer, F.S.; Galli, V.; Millwood, R.J.; Cechin, J.; Agostinetto, D.; Maia, L.C.; Vargas, L.; et al. Transcriptomic Analysis Identifies New Non-Target Site Glyphosate-Resistance Genes in Conyza bonariensis. Plants 2019, 8, 157. [Google Scholar] [CrossRef] [PubMed]
- Calha, I.M.; Osuna, M.D. Herbicide weed resistance in portuguese olive groves. In Proceedings of the 21th Columa Conference, Dijon, France, 8–9 September 2010. [Google Scholar]
- Simões, M.; Portugal, J.; Monteiro, A.; Calha, I.M. Citrus orchard with Conyza bonariensis resistant to glyphosate: 211–216. In Proceedings of the Actas del XIV Congreso SEMh, Sociedad Española de Malherbología, Valencia, Spain, 5–7 November 2013. [Google Scholar]
- Domingos, D.; Rodrigues, M.; Andrade, E.; Monteiro, A.; Calha, I.M. Genetic Characterization of EPSPs from Conyza canadensis resistant to glyphosate. In Proceedings of the 17th EWRS Symposium “Weed Management in Changing Environments”, Montpellier, France, 23–26 June 2015. [Google Scholar]
- Rey-Caballero, J.; Royo-Esnal, A.; Recasens, J.; González, I.; Torra, J. Management Options for Multiple Herbicide–Resistant Corn Poppy (Papaver rhoeas) in Spain. Weed Sci. 2017, 65, 295–304. [Google Scholar] [CrossRef]
- Torra, J.; Esnal, A.R.; Guinjuan, J.R.; Recasens, J. Management of herbicide-resistant Papaver rhoeas in dry land cereal fields. Agron. Sustain. Dev. 2011, 31, 483–490. [Google Scholar] [CrossRef]
- Cirujeda, A.; Recasens, J.; Torra, J.; Taberner, A. A germination study of herbicide-resistant field poppies in Spain. Agron. Sustain. Dev. 2008, 28, 207–220. [Google Scholar] [CrossRef][Green Version]
- Torra, J.; Gonzalez-Andujar, J.L.; Recasens, J. Modelling the population dynamics of Papaver rhoeas under various weed man-agement systems in a Mediterranean climate. Weed Res. 2008, 48, 136–146. [Google Scholar] [CrossRef]
- Torra, J.; Recasens, J. Demography of Corn Poppy (Papaver rhoeas) in Relation to Emergence Time and Crop Competition. Weed Sci. 2008, 56, 826–833. [Google Scholar] [CrossRef]
- Torra, J.; Cirujeda, A.; Recasens, J.; Taberner, A.; Powles, S.B. PIM (Poppy Integrated Management): A bio-economic decision support model for the management of Papaver rhoeas in rain-fed cropping systems. Weed Res. 2010, 50, 127–139. [Google Scholar] [CrossRef]
- Torra, J.; Cirujeda, A.; Taberner, A.; Recasens, J. Evaluation of herbicides to manage herbicide-resistant corn poppy (Papaver rhoeas) in winter cereals. Crop Prot. 2010, 29, 731–736. [Google Scholar] [CrossRef]
- Torra, J.; Royo-Esnal, A.; Rey-Caballero, J.; Recasens, J.; Salas, M. Management of Herbicide-Resistant Corn Poppy (Papaver rhoeas) under Different Tillage Systems Does Not Change the Frequency of Resistant Plants. Weed Sci. 2018, 66, 764–772. [Google Scholar] [CrossRef]
- Recasens, J.; Royo-Esnal, A.; Valencia-Gredilla, F.; Torra, J. Efficiency, profitability and carbon footprint of different manage-ment programs under no-till to control herbicide resistant Papaver rhoeas. Plants 2020, 9, 433. [Google Scholar] [CrossRef]
- Torra, J.; Rojano-Delgado, A.M.; Rey-Caballero, J.; Royo-Esnal, A.; Salas, M.L.; De Prado, R. Enhanced 2,4-D Metabolism in Two Resistant Papaver rhoeas Populations from Spain. Front. Plant Sci. 2017, 8, 1584. [Google Scholar] [CrossRef] [PubMed]
- Leon, R.G. Of Amaranthus palmeri and other demons. Inf. Téc. Econ. Agrar. 2020, 116, 241–255. [Google Scholar]
- Peterson, M.A.; Collavo, A.; Ovejero, R.; Shivrain, V.; Walsh, M.J. The challenge of herbicide resistance around the world: A current summary. Pest Manag. Sci. 2018, 74, 2246–2259. [Google Scholar] [CrossRef] [PubMed]
- European Commission (EC). Commission Implementing Regulation (EU) 2021/824 of 21 May 2021 Amending Imple-Menting Regulations (EU) No 540/2011 and (EU) No 820/2011 as Regards the Conditions of Approval of the Active Substance Terbuthylazine. 2022. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32021R0824 (accessed on 4 April 2022).
- Melander, B.; Munier-Jolain, N.; Charles, R.; Wirth, J.; Schwarz, J.; Van Der Weide, R.; Bonin, L.; Jensen, P.K.; Kudsk, P. European Perspectives on the Adoption of Nonchemical Weed Management in Reduced-Tillage Systems for Arable Crops. Weed Technol. 2013, 27, 231–240. [Google Scholar] [CrossRef]
- Gaines, T.A.; Busi, R.; Küpper, A. Can new herbicide discovery allow weed management to outpace resistance evolution? Pest Manag. Sci. 2021, 77, 3034–3039. [Google Scholar] [CrossRef]
- Dayan, F.E. Current Status and Future Prospects in Herbicide Discovery. Plants 2019, 8, 341. [Google Scholar] [CrossRef]
- Moss, S. Integrated weed management (IWM): Why are farmers reluctant to adopt non-chemical alternatives to herbicides? Pest Manag. Sci. 2019, 75, 1205–1211. [Google Scholar] [CrossRef]
- Vazquez-Garcia, J.G.; Torra, J.; Palma-Bautista, C.; De Prado, R. Comparison between the mechanisms of Clearfield® wheat and Lolium rigidum multiple resistant to ACCase and ALS inhibiting herbicides. Environ. Pollut. 2022; minor revision. [Google Scholar]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.-P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef]
- Tétard-Jones, C.; Sabbadin, F.; Moss, S.; Hull, R.; Neve, P.; Edwards, R. Changes in the proteome of the problem weed black-grass correlating with multiple–herbicide resistance. Plant J. 2018, 94, 709–720. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
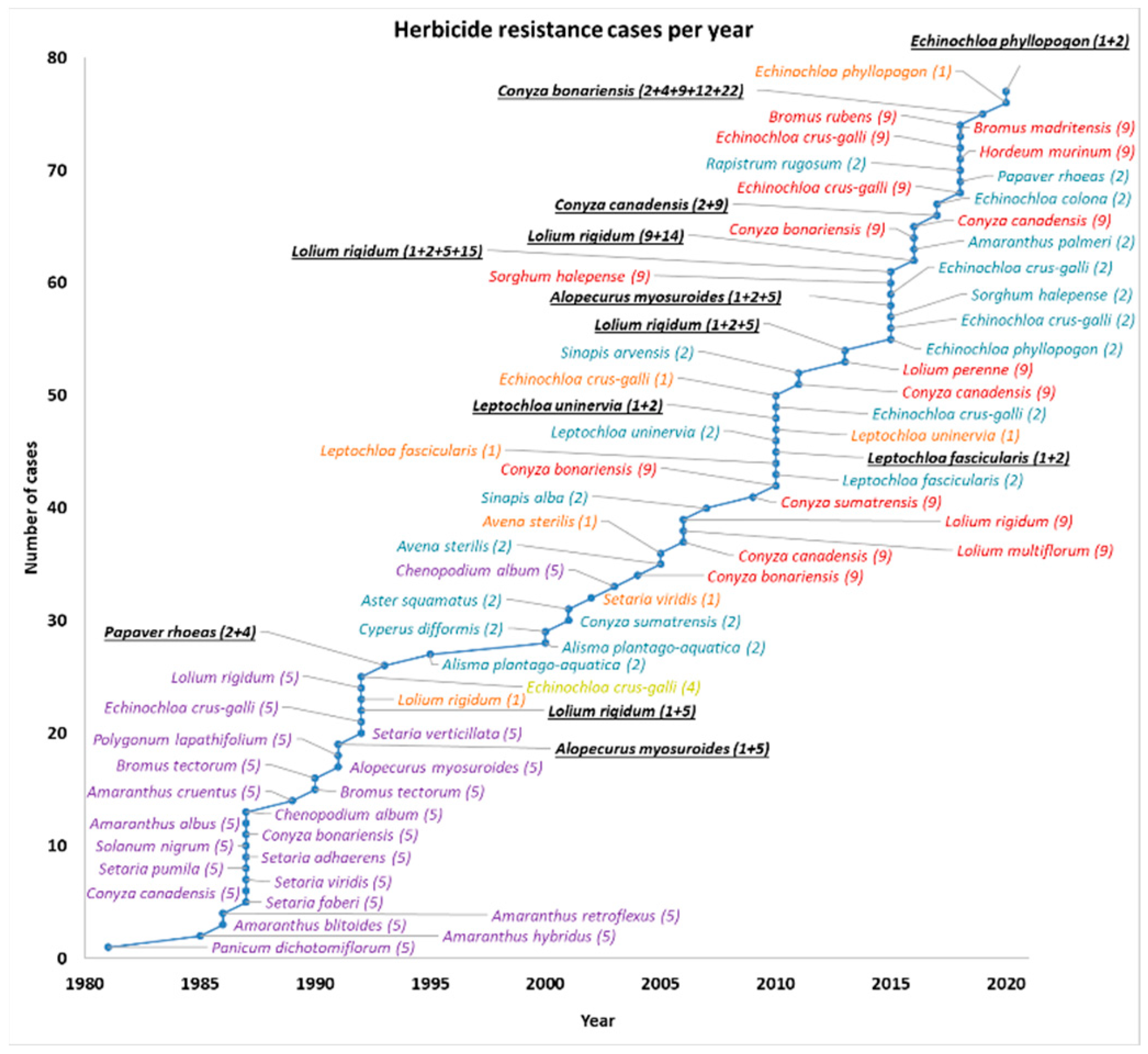
| Number | Species | Year | MoAs | Herbicides | Crops/Situations | References |
|---|---|---|---|---|---|---|
| 1 | Alisma plantago-aquatica | 1995 | ALS inhibitors (2) | Bensulfuron | Rice | [9,10] |
| 2 | Chenopodium album | 2003 | Photosystem II inhibitors (5) | Atrazine | Corn | [8] |
| 3 | Conyza bonariensis | 2010 | EPSP synthase inhibitors (9) | Glyphosate | Orchards | [4] |
| 4 | Conyza canadensis | 2011 | EPSP synthase inhibitors (9) | Glyphosate | Olive | [4] |
| 5 | Lolium perenne | 2013 | EPSP synthase inhibitors (9) | Glyphosate | Vineyards | [11] |
| 6 | Echinochloa phyllopogon | 2015 | ALS inhibitors (2) | Penoxsulam | Rice | Calha * |
| 7 | Echinochloa crus-galli | 2015 | ALS inhibitors (2) | Penoxsulam | Rice | Calha * |
| 8 | Echinochloa crus-galli | 2018 | EPSP synthase inhibitors (9) | Glyphosate | Pomegranates, corn, vineyards | [12] |
| 9 | Papaver rhoeas | 2018 | ALS inhibitors (2) | Tribenuron | Cereals | Portugal * |
| 10 | Echinochloa phyllopogon | 2020 | ACCase inhibitors (1) | Profoxydim | Rice | Calha * |
| 11 | Echinochloa phyllopogon | 2020 | ALS inhibitors (2), ACCase inhibitors (1) | Profoxydim, penoxsulam | Rice | Calha * |
| Number | Species | Year | MoAs | Herbicides | Crops/Situations | References |
|---|---|---|---|---|---|---|
| 1 | Panicum dichotomiflorum | 1981 | Photosystem II inhibitors (5) | Atrazine | Corn, cropland | [13] |
| 2 | Amaranthus hybridus | 1985 | Photosystem II inhibitors (5) | Atrazine | Corn | [14] |
| 3 | Amaranthus blitoides | 1986 | Photosystem II inhibitors (5) | Simazine | Orchards | [14] |
| 4 | Amaranthus retroflexus | 1986 | Photosystem II inhibitors (5) | Atrazine | Corn | [14] |
| 5 | Setaria faberi | 1987 | Photosystem II inhibitors (5) | Atrazine | Corn | [15] |
| 6 | Conyza canadensis | 1987 | Photosystem II inhibitors (5) | Atrazine, simazine | Corn, orchards, roadsides | [4,16] |
| 7 | Setaria viridis | 1987 | Photosystem II inhibitors (5) | Atrazine | Corn | [15] |
| 8 | Setaria pumila | 1987 | Photosystem II inhibitors (5) | Atrazine | Corn | [15] |
| 9 | Setaria adhaerens | 1987 | Photosystem II inhibitors (5) | Atrazine | Corn | [15] |
| 10 | Solanum nigrum | 1987 | Photosystem II inhibitors (5) | Atrazine | Corn | [14] |
| 11 | Conyza bonariensis | 1987 | Photosystem II inhibitors (5) | Simazine | Orchards | [4,16] |
| 12 | Amaranthus albus | 1987 | Photosystem II inhibitors (5) | Simazine | Orchards | [14] |
| 13 | Chenopodium album | 1987 | Photosystem II inhibitors (5) | Atrazine | Corn | [16] |
| 14 | Amaranthus cruentus | 1989 | Photosystem II inhibitors (5) | Atrazine | Corn | [17] |
| 15 | Bromus tectorum | 1990 | PSII inhibitor (ureas and amides) (5) | Chlorotoluron | Wheat | [18] |
| 16 | Bromus tectorum | 1990 | Photosystem II inhibitors (5) | Simazine | Orchards | [19] |
| 17 | Alopecurus myosuroides | 1991 | PSII inhibitor (ureas and amides) (5) | Chlorotoluron | Wheat | [20] |
| 18 | Polygonum lapathifolium | 1991 | Photosystem II inhibitors (5) | Atrazine | Corn | [13] |
| 19 | Alopecurus myosuroides | 1991 | ACCase inhibitors (1), PSII inhibitor (Ureas and amides) (5) | Diclofop, chlorotoluron | Wheat | [21] |
| 20 | Setaria verticillata | 1992 | Photosystem II inhibitors (5) | Atrazine | Corn | [15] |
| 21 | Echinochloa crus-galli | 1992 | Photosystem II inhibitors (5) | Atrazine | Corn | [15] |
| 22 | Lolium rigidum | 1992 | ACCase inhibitors (1), PSII inhibitor (ureas and amides) (5) | Diclofop, chlorotoluron | Wheat | [22] |
| 23 | Lolium rigidum | 1992 | ACCase inhibitors (1) | Diclofop | Wheat | [23] |
| 24 | Lolium rigidum | 1992 | Photosystem II inhibitors (5) | Simazine | Orchards | [22] |
| 25 | Echinochloa crus-galli | 1992 | Synthetic Auxins (4) | Quinclorac | Rice | [24] |
| 26 | Papaver rhoeas | 1993 | ALS inhibitors (2), Synthetic Auxins (4) | Tribenuron, 2,4-D | Cereals, wheat | [25] |
| 27 | Alisma plantago-aquatica | 2000 | ALS inhibitors (2) | Bensulfuron | Rice | [16] |
| 28 | Cyperus difformis | 2000 | ALS inhibitors (2) | Bensulfuron, penoxsulam | Rice | [26] |
| 29 | Conyza sumatrensis | 2001 | ALS inhibitors (2) | Imazapyr | Roadsides | [27] |
| 30 | Aster squamatus | 2001 | ALS inhibitors (2) | Imazapyr | Roadsides | [28] |
| 31 | Setaria viridis | 2002 | ACCase inhibitors (1) | Sethoxydim, haloxyfop | Onion | [29] |
| 32 | Avena sterilis | 2005 | ALS inhibitors (2) | Imazamethaben | Winter cereals | [8] |
| 33 | Avena sterilis | 2005 | ACCase inhibitors (1) | Fenoxaprop | Winter cereals | [8] |
| 34 | Conyza bonariensis | 2004 | EPSP synthase inhibitors (9) | Glyphosate | Orchards | [30] |
| 35 | Conyza canadensis | 2006 | EPSP synthase inhibitors (9) | Glyphosate | Orchards | [31] |
| 36 | Lolium multiflorum | 2006 | EPSP synthase inhibitors (9) | Glyphosate | Orchards | [11] |
| 37 | Lolium rigidum | 2006 | EPSP synthase inhibitors (9) | Glyphosate | Orchards | [11] |
| 38 | Sinapis alba | 2007 | ALS inhibitors (2) | Tribenuron, iodosulfuron | Winter wheat | [32] |
| 39 | Conyza sumatrensis | 2009 | EPSP synthase inhibitors (9) | Glyphosate | Orchards | [33] |
| 40 | Leptochloa fascicularis | 2010 | ALS inhibitors (2) | Penoxsulam | Rice | [34,35] |
| 41 | Leptochloa fascicularis | 2010 | ACCase inhibitors (1) | Profoxydim, cyhalofop | Rice | [34,35] |
| 42 | Leptochloa fascicularis | 2010 | ALS inhibitors (2), ACCase inhibitors (1) | Profoxydim, cyhalofop, penoxsulam | Rice | [34,35] |
| 43 | Leptochloa uninervia | 2010 | ALS inhibitors (2) | Penoxsulam | Rice | [34,35] |
| 44 | Leptochloa uninervia | 2010 | ACCase inhibitors (1) | Cyhalofop | Rice | [34,35] |
| 45 | Leptochloa uninervia | 2010 | ALS inhibitors (2), ACCase inhibitors (1) | Profoxydim, cyhalofop, penoxsulam | Rice | [34,35] |
| 46 | Echinochloa crus-galli | 2010 | ALS inhibitors (2) | Penoxsulam | Rice | [36] |
| 47 | Echinochloa crus-galli | 2010 | ACCase inhibitors (1) | Profoxydim, cyhalofop | Rice | [36] |
| 48 | Sinapis arvensis | 2011 | ALS inhibitors (2) | Tribenuron, iodosulfuron | Cereals | [4] |
| 49 | Lolium rigidum | 2013 | ALS inhibitors (2), ACCase inhibitors (1), PSII inhibitor (ureas and amides) (5) | Chlortoluron, chlorsulfuron, diclofop | Cereals | [37] |
| 50 | Sorghum halepense | 2015 | ALS inhibitors (2) | Nicosulfuron | Corn | [38] |
| 51 | Alopecurus myosuroides | 2015 | ACCase inhibitors (1), ALS inhibitors (2), PSII inhibitor (ureas and amides) (5) | Fops, dens, cloransulam, ureas, SUs | Wheat, canola, peas, winter barley, faba beans | [4] |
| 52 | Echinochloa crus-galli | 2015 | ALS inhibitors (2) | Nicosulfuron | Corn | [4,39] |
| 53 | Sorghum halepense | 2015 | EPSP synthase inhibitors (9) | Glyphosate | Railways, freeways | [40] |
| 54 | Lolium rigidum | 2015 | ALS inhibitors (2), ACCase inhibitors (1), PSII inhibitor (Ureas, amides) (5), Very long-chain fatty acid synthesis inhibitors (15) | fops, dims, pyroxsulam, mesosulfuron, chlortoluron, prosulfocarb | Winter cereals | [41] |
| 55 | Lolium rigidum | 2016 | EPSP synthase inhibitors (9), PPO inhibitors (14) | Oxyfluorfen, glyphosate | Olive | [11] |
| 56 | Amaranthus palmeri | 2016 | ALS inhibitors (2) | Nicosulfuron | Corn, roadsides | [42] |
| 57 | Conyza bonariensis | 2016 | EPSP synthase inhibitors (9) | Glyphosate | Railways | [43] |
| 58 | Conyza canadensis | 2016 | EPSP synthase inhibitors (9) | Glyphosate | Railways | [43] |
| 59 | Conyza canadensis | 2017 | ALS inhibitors (2), EPSP synthase inhibitors (9) | Tribenuron, glyphosate | Olive | [44] |
| 60 | Echinochloa colona | 2017 | ALS inhibitors (2) | Penoxsulam | Rice | [36] |
| 61 | Rapistrum rugosum | 2018 | ALS inhibitors (2) | Tribenuron, iodosulfuron | Winter cereals | [45] |
| 62 | Hordeum murinum | 2018 | EPSP synthase inhibitors (9) | Glyphosate | Orchards, olive | [46] |
| 63 | Echinochloa crus-galli | 2018 | EPSP synthase inhibitors (9) | Glyphosate | Olive, citrus, orchards, corn, rice | [12] |
| 64 | Bromus madritensis | 2018 | EPSP synthase inhibitors (9) | Glyphosate | Winter cereals | [4] |
| 65 | Bromus rubens | 2018 | EPSP synthase inhibitors (9) | Glyphosate | Orchards, olive | [47] |
| 66 | Conyza bonariensis | 2019 | ALS inhibitors (2), Synthetic Auxins (4), EPSP synthase inhibitors (9), PDS inhibitors (12), PSI electron diverters (22) | Tribenuron, 2,4-D, glyphosate, diflufenican, paraquat | Olive | [48] |
| HRAC & WSSA | Legacy HRAC | Molecular Target |
|---|---|---|
| 1 | A | Inhibition of Acetyl-CoA carboxylase (ACCase) |
| 2 | B | Inhibition of Acetolactate synthase (ALS) |
| 3 | K1 | Inhibition of microtubule assembly |
| 4 | O | Auxin mimics |
| 5 | C1, C2 | Inhibition of photosynthesis. Photosystem II (PSII)—Serine 264 binder |
| 6 | C3 | Inhibition of photosynthesis. PSII—Histidine 215 binder |
| 9 | G | Inhibition of 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) |
| 10 | H | Inhibition of glutamine synthetase |
| 12 | F1 | Inhibition of phytoene desaturase (PDS) |
| 13 | F4 | Inhibition of 1-deoxy-D-xyulose 5-phosphate synthase (DOXP) synthase |
| 14 | E | Inhibition of protoporphyrinogen oxidase (PPO) |
| 15 | K3 | Inhibition of very long-chain fatty acid (VLCFA) synthesis |
| 18 | I | Dihydropteroate (DHP) synthase inhibition |
| 19 | P | Auxin transport inhibitors |
| 22 | D | Photosystem I (PSI) electron diversion |
| 23 | K2 | Inhibition of microtubule organization |
| 24 | M | Uncouplers |
| 27 | F2 | Inhibition of 4-Hydroxyphenylpyruvate dioxygenase (HPPD) |
| 29 | L | Inhibition of cellulose synthesis |
| 30 | Q | Inhibition of fatty acid thioesterase |
| 31 | R | Inhibition of serine threonine protein phosphatase |
| 32 | S | Inhibition of solanesyl diphosphate synthase |
| 33 | T | Inhibition of homogentisate solanesyltransferase |
| 34 | F3 | Inhibition of lycopene cyclase |
| 0 | Z | Unknown mode of action |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torra, J.; Montull, J.M.; Calha, I.M.; Osuna, M.D.; Portugal, J.; de Prado, R. Current Status of Herbicide Resistance in the Iberian Peninsula: Future Trends and Challenges. Agronomy 2022, 12, 929. https://doi.org/10.3390/agronomy12040929
Torra J, Montull JM, Calha IM, Osuna MD, Portugal J, de Prado R. Current Status of Herbicide Resistance in the Iberian Peninsula: Future Trends and Challenges. Agronomy. 2022; 12(4):929. https://doi.org/10.3390/agronomy12040929
Chicago/Turabian StyleTorra, Joel, José M. Montull, Isabel M. Calha, María D. Osuna, Joao Portugal, and Rafael de Prado. 2022. "Current Status of Herbicide Resistance in the Iberian Peninsula: Future Trends and Challenges" Agronomy 12, no. 4: 929. https://doi.org/10.3390/agronomy12040929
APA StyleTorra, J., Montull, J. M., Calha, I. M., Osuna, M. D., Portugal, J., & de Prado, R. (2022). Current Status of Herbicide Resistance in the Iberian Peninsula: Future Trends and Challenges. Agronomy, 12(4), 929. https://doi.org/10.3390/agronomy12040929