
Description of Filenchus Species from Agroecosystem of Southern Alberta, Canada



Abstract
:1. Introduction
2. Materials and Methods
2.1. Nematode Isolation and Morphological Studies
2.2. DNA Extraction, PCR, and Sequencing
2.3. Phylogenetic Analyses
3. Results
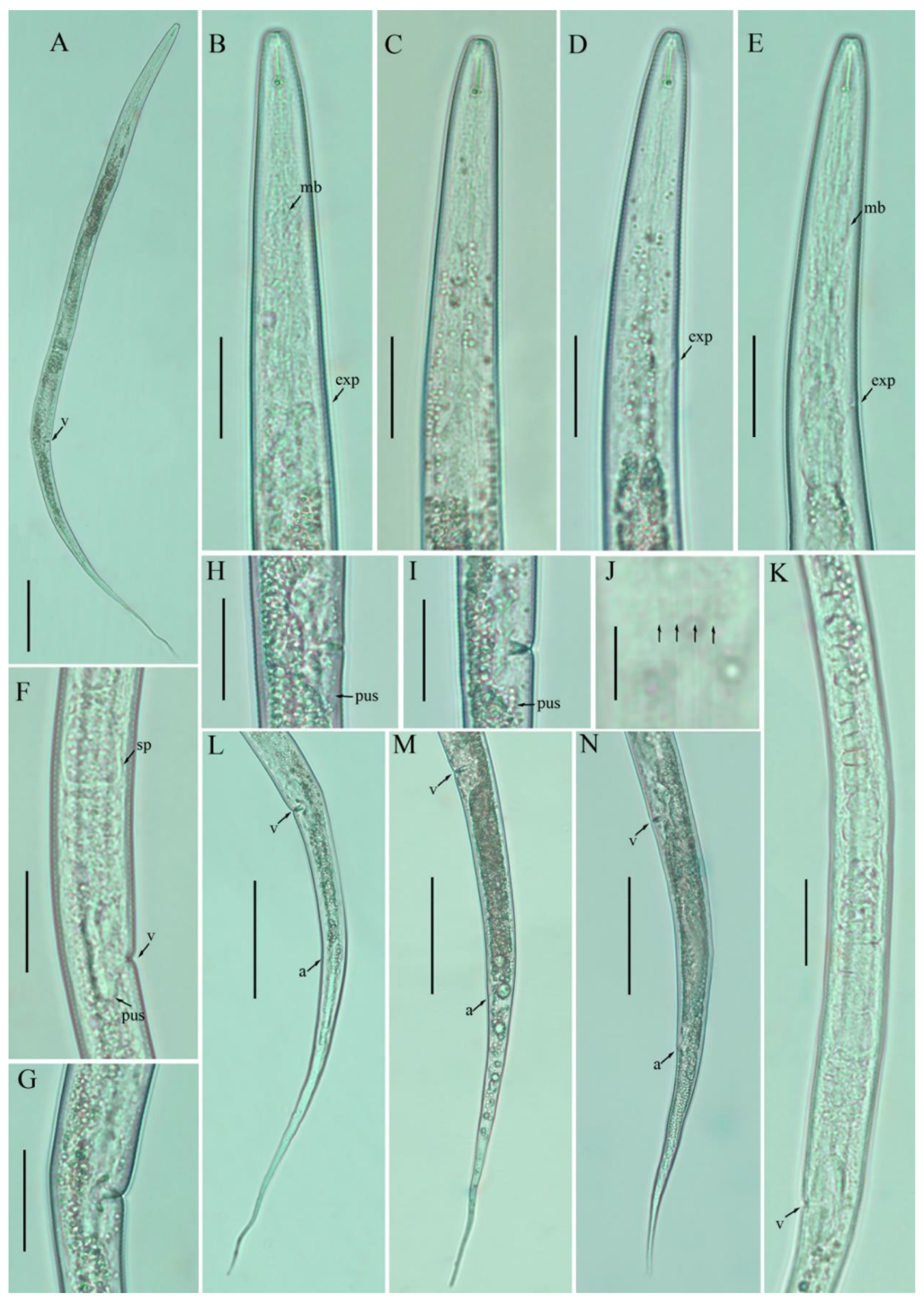
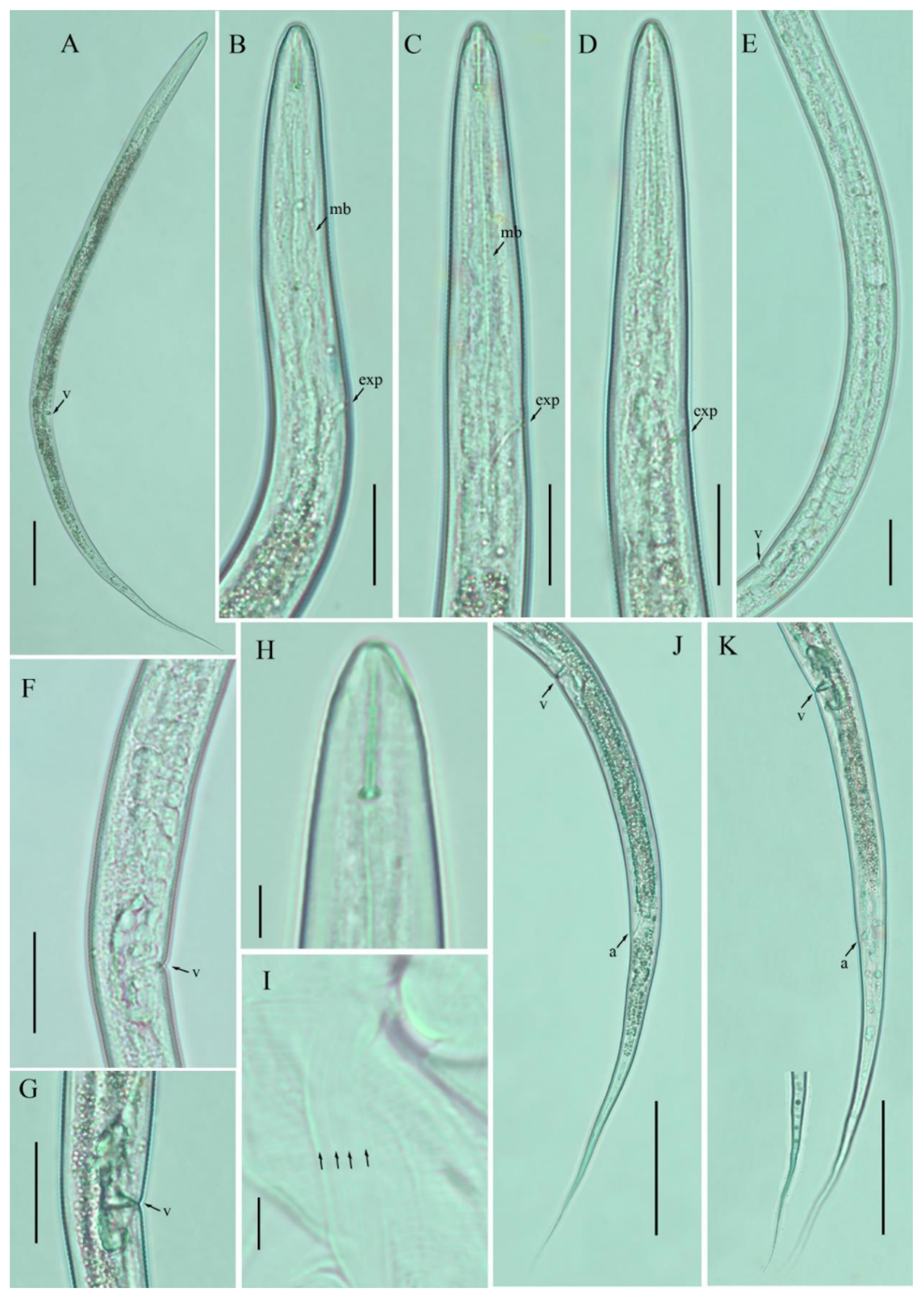
3.1. Description of Filenchus cylindricus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | This Study | Thorne and Malek [19] | Elmiligy [41] | Raski and Geraert [6] | Zeidan and Geraert [42] | Dobrin and Geraert [43] |
|---|---|---|---|---|---|---|
| Locality | Alberta, Canada | South Dakota, USA | Iowa, USA | Colorado, USA | Sudan | Romania |
| n | 13 | 1 | 10 | 8 | 8 *1 | 6 *1 |
| Body length | 642.4 ± 42.4 (557.0–711.0) | 1000 | 896 (750–990) | 1060 (970–1150) | 700–870 | 600–710 |
| a | 36.6 ± 2.5 (34.0–43.0) | 40 | 39 (30–45) | 39 (34–43) | 33–41 | 28.1–39.4 |
| b | 6.5 ± 0.4 (6.0–7.2) | 6.6 | 6.4 (6.0–7.8) | 6.7 (6.1–7.6) | 5.7–6.9 | 4.6–6.5 |
| c | 4.8 ± 0.4 (4.4–5.5) | 6.5 | 5.2 (4.7–5.7) | 5.0 (4.7–5.2) | 4.6–5.6 | 4.0–4.7 |
| c′ | 13.0 ± 1.4 (11.1–15.2) | – | 11.2 (10.2–17.0) | 11.6 (9.8–13.2) | 9.3–11.9 | 10–15.6 |
| MB | 41.2 ± 1.3 (40.0–43.6) | – | 42 | 43.8 (42–46) | 42–44 | 44–54 |
| V | 62.0 ± 1.2 (59.2–64.4) | 64 | 60 (55–64) | 60 (57–62) | 59–65 | 53–61.6 |
| Lip height | 3.1 ± 0.1 (3.0–3.5) | – | 1.7–2.0 | 3.5 | – | – |
| Lip width | 6.2 ± 0.3 (6.0–7.0) | – | – | 7.5–8.0 | – | – |
| Stylet length | 12.1 ± 0.8 (10.0–13.0) | 13 | 13 (12–14) | 12.4 (12–13) | 12.0–13.0 | 12–14.5 |
| Anterior end to excretory pore | 81.0 ± 3.1 (75.0–86.0) | – | – | – | – | – |
| Pharynx length | 98.2 ± 3.8 (91.0–103.0) | – | – | – | – | – |
| Maximum body width | 17.6 ± 1.8 (15.0–21.0) | – | – | – | – | – |
| Vulva body width | 16.6 ± 1.0 (15.0–18.0) | – | – | – | – | – |
| Post-uterine sac (PUS) length | 11.0 ± 1.6 (9.0–14.0) | – | – | – | – | – |
| Distance from vulva to anus | 110.2 ± 13.1 (98.0–139.0) | – | – | – | – | – |
| Distance from vulva to tail terminus | 245.3 ± 18.7 (210.0–279.0) | – | – | – | – | – |
| Anal body width | 10.5 ± 1.1 (9.0–13.0) | – | – | – | – | – |
| Tail length | 135.1 ± 16.5 (112.0–159.0) | – | 172 (154–194) | 213 (189–242) | 140–178 | – |
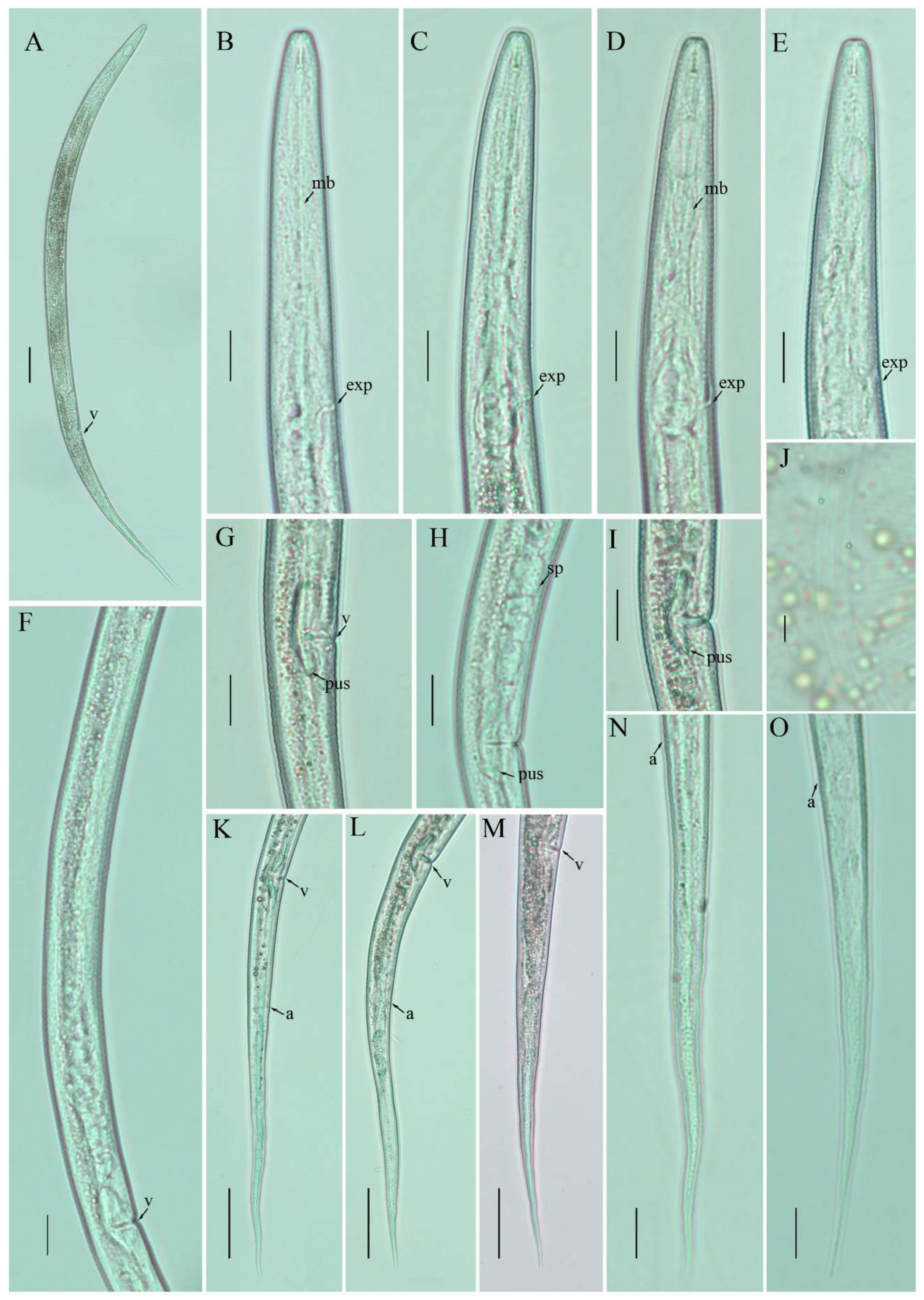
3.2. Description of Filenchus hazenensis

| Characteristics | This Study | Wu [16] | Brzeski [44] |
|---|---|---|---|
| Locality | Alberta, Canada | Nunavut, Canada | Poland |
| n | 17 | 4 | 5 |
| Body length | 655.6 ± 34.4 (547.0–701.0) | 1020–1090 | 877–1003 |
| a | 34.2 ± 3.2 (29.4–39.3) | 38–39 | 36.2 (33–39) |
| b | 6.0 ± 0.3 (5.0–6.3) | 6.6–6.8 | 6.5 (6.1–7.4) |
| c | 4.5 ± 0.2 (4.0–5.0) | 4.4–4.7 | 5.0 (4.6–6.3) |
| c′ | 13.0 ± 1.5 (10.2–15.0) | – | 11.5 (8.4–13.3) |
| MB | 42.0 ± 1.5 (39.5–45.0) | – | 43.6 (42–45) |
| V | 61.4 ± 1.4 (56.7–62.4) | 60–62 | 61.1 (57–60) |
| Lip height | 4.4 ± 0.2 (4.0–5.0) | – | – |
| Lip width | 6.5 ± 0.3 (6.0–7.0) | – | 8–9 |
| Stylet length | 14.0 ± 0.6 (13.0–15.0) | 14.6–15.5 | 14.5 (14–15) |
| Anterior end to excretory pore | 89.0 ± 4.2 (81.0–95.0) | 119–125 | – |
| Pharynx length | 109.4 ± 3.1 (104.0–114.0) | 149–165 | 143 (134–152) |
| Maximum body width | 19.3 ± 1.6 (16.0–22.0) | – | – |
| Vulva body width | 17.1 ± 1.3 (15.0–20.0) | – | – |
| Post-uterine sac (PUS) length | 12.6 ± 1.5 (9.0–15.0) | 15–17 | |
| Distance from vulva to anus | 108.6 ± 6.0 (100.0–119.0) | – | – |
| Distance from vulva to tail terminus | 253.0 ± 9.9 (237.0–275.0) | – | – |
| Anal body width | 11.2 ± 1.1 (10.0–14.0) | – | – |
| Tail length | 144.2 ± 6.4 (133.0–155.0) | 224–232 | 189 (138–219) |
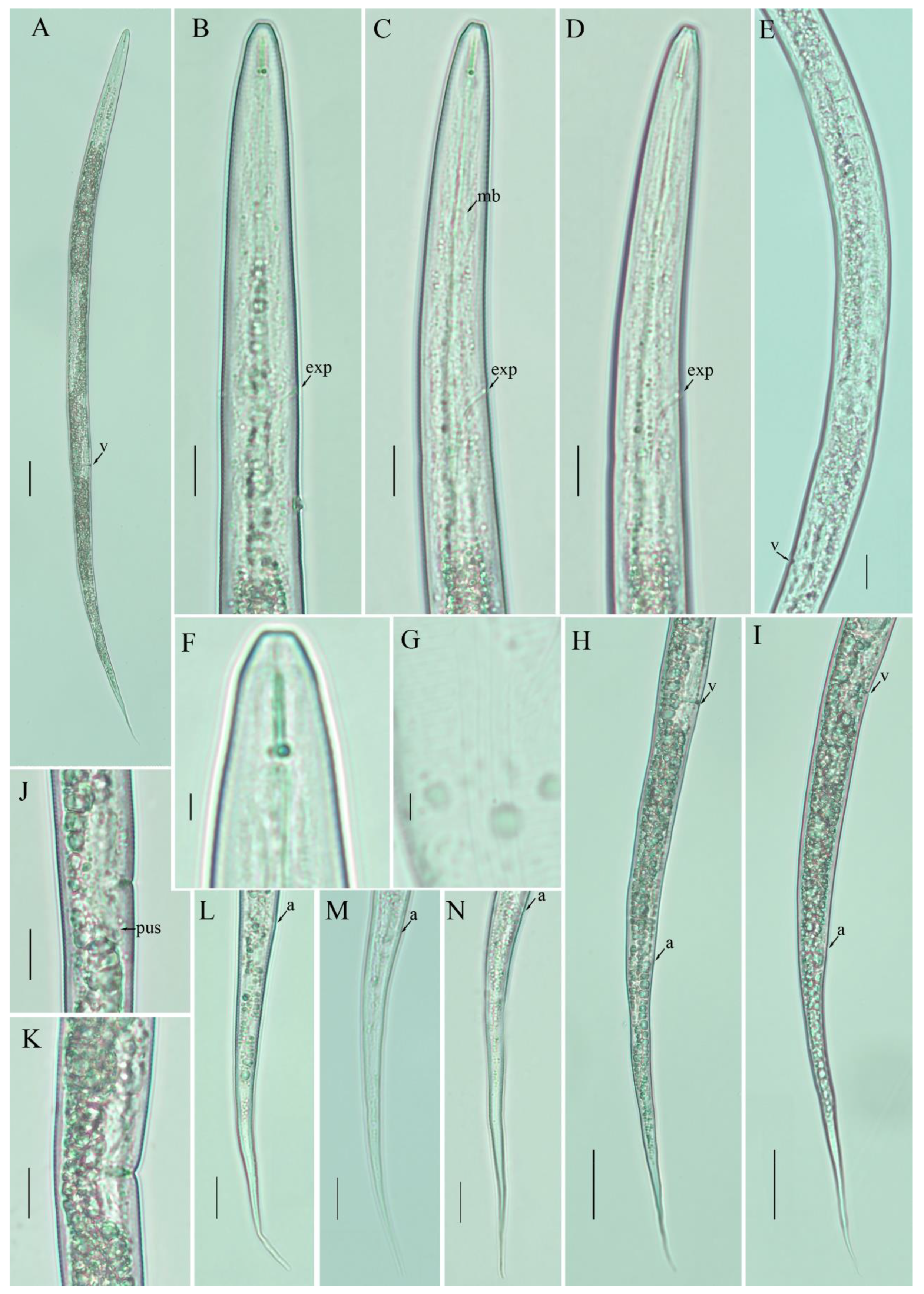
3.3. Description of Filenchus sheri

| Characteristics | This Study | Khan and Khan [21] | Karegar and Geraert [45] |
|---|---|---|---|
| Locality | Alberta, Canada | Afghanistan | Iran |
| n | 17 | 5 | 4 |
| Body length | 618.6 ± 28.0 (567.0–679.0) | 510 (460–550) | 515 (490–535) |
| a | 35.2 ± 3.0 (31.5–42.3) | 33 (31–38) | 35.3 (31.1–39.9) |
| b | 6.8 ± 0.3 (6.1–7.3) | 502 (5–6) | 5.6 (5.3–6.1) |
| c | 5.5 ± 0.3 (5.0–6.2) | 4.5 (4–5) | 3.8 (3.6–3.9) |
| c′ | 11.5 ± 1.1 (9.8–14.0) | – | 14.8 (13.6–15.8) |
| MB | 41.2 ± 3.1 (34.5–47.0) | 40–45 | 40.4 (38.9–43.2) |
| V | 69.0 ± 1.3 (66.0–71.0) | 60.2 (61–63) | 56.1 (56.1–60.7) |
| Lip height | 2.5 ± 0.2 (2.0–3.0) | – | – |
| Lip width | 5.8 ± 0.4 (5.0–6.5) | – | – |
| Stylet length | 7.7 ± 0.4 (7.0–8.5) | 7.6 (7–8) | – |
| Anterior end to excretory pore | 80.6 ± 1.4 (78.0–83.0) | – | 72.5 (66–77) |
| Pharynx length | 91.6 ± 3.4 (85.0–98.0) | – | – |
| Maximum body width | 17.7 ± 1.2 (15.0–19.0) | – | – |
| Vulva body width | 15.3 ± 0.7 (14.0–17.0 | – | – |
| Post-uterine sac (PUS) length | 9.5 ± 1.5 (7.5–13.0) | – | – |
| Distance from vulva to anus | 78.4 ± 6.4 (71.0–90.0) | – | – |
| Distance from vulva to tail terminus | 191.8 ± 9.0 (173.0–206.0) | – | – |
| Anal body width | 9.9 ± 0.5 (9.0–10.5) | – | – |
| Tail length | 113.4 ± 7.9 (98.0–126.0) | – | 136 (127–142) |
3.4. Description of Filenchus thornei
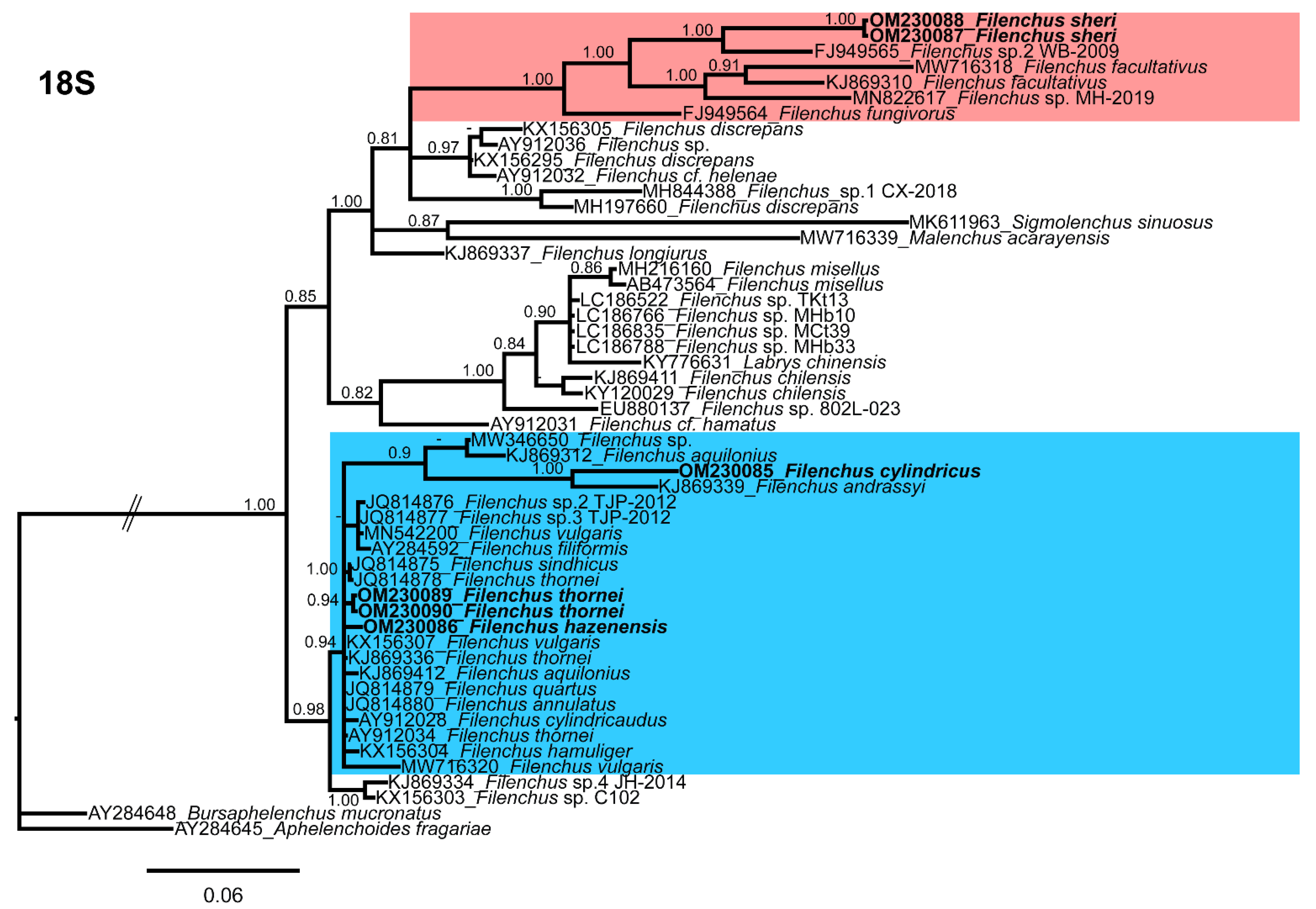
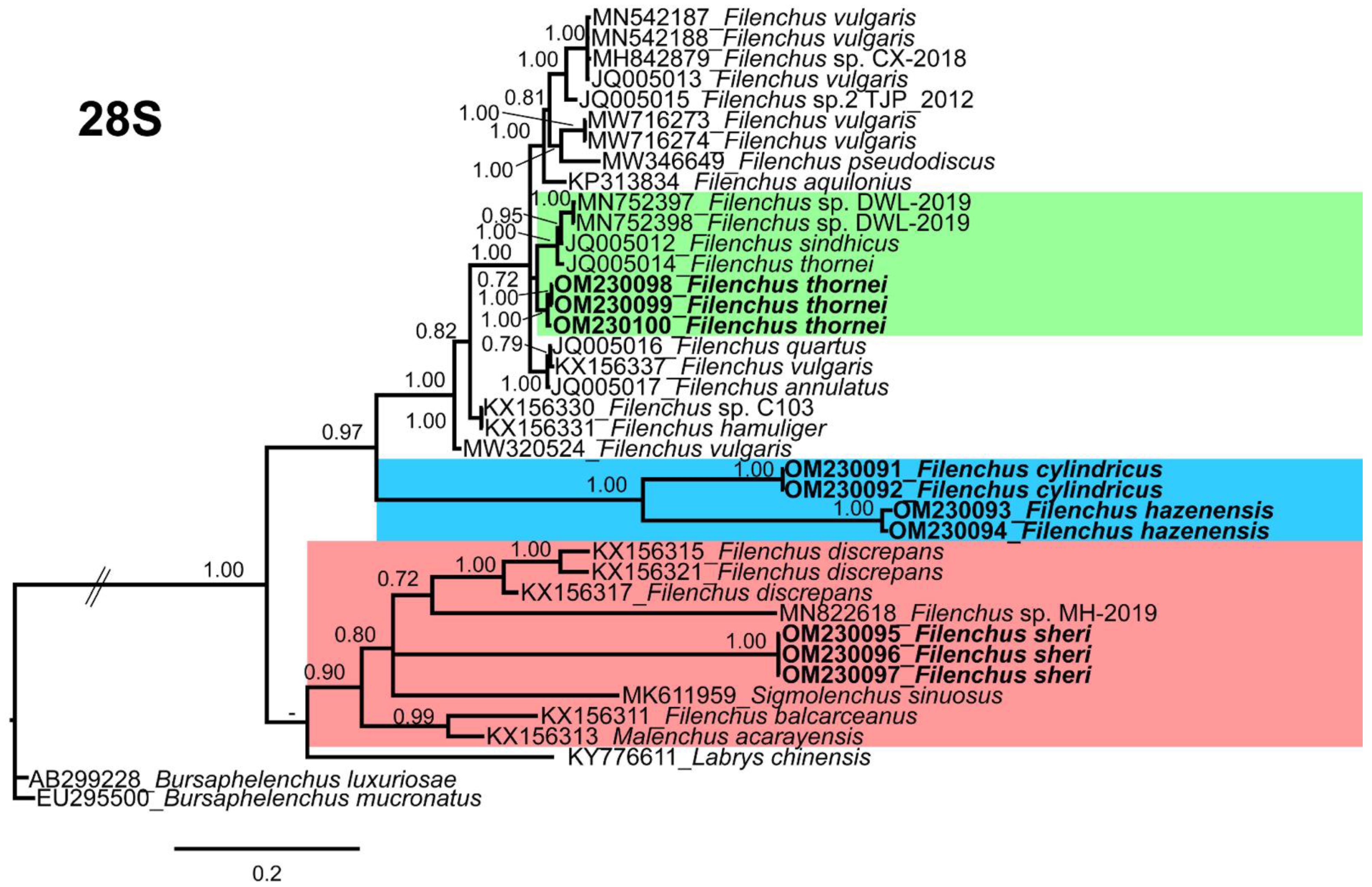
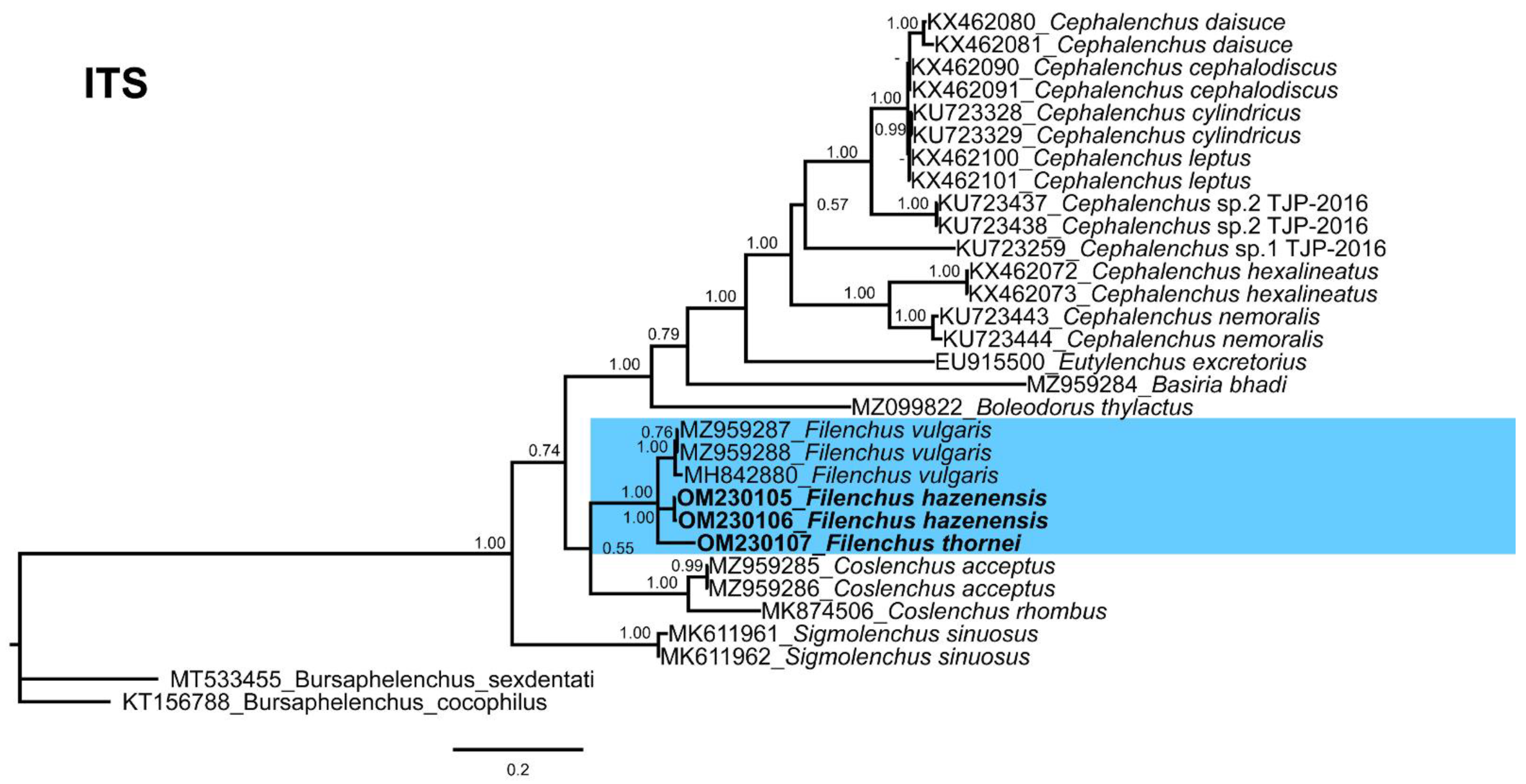
3.5. Molecular Characterization and Phylogenetic Relationships of Detected Filenchus Species with Related Filenchus Species

| Characteristics | This Study | Andrássy [1] | Raski and Geraert [6] | Castillo et al. [48] |
|---|---|---|---|---|
| Locality | Alberta, Canada | Bulgaria | Colorado, USA | Spain |
| n | 16 | 1 | 15 | 24 |
| Body length | 633.2 ± 37.1 (582.0–701.0) | 739 | 780 (710–870) | 734 (616–840) |
| a | 30.4 ± 2.6 (26.0–35.5) | 36.7 | 40 (37–46) | 37.5 (32.2–42.9) |
| b | 6.1 ± 0.3 (5.6–6.7) | 7.8 | 6.8 (6.4–8.2) | 6.5 (5.4–7.4) |
| c | 4.5 ± 0.4 (3.9–5.4) | 3.9 | 3.6 (3.4–4.1) | 4.0 (3.5–4.7) |
| c′ | 11.4 ± 1.4 (8.7–13.9) | – | – | 14.6 (12.1–17.5) |
| MB | 42.7 ± 3.3 (38.8–49.1) | 41 | 40 (38–42) | 40 (37–46) |
| V | 59.0 ± 2.5 (53.0–63.4) | 58.1 | 56 (53–60) | 59 (57–63) |
| Lip height | 3.2 ± 0.2 (3.0–4.0) | – | – | – |
| Lip width | 6.0 ± 0.3 (5.5–7.0) | – | – | – |
| Stylet length | 10.3 ± 0.8 (9.0–11.5) | 10.5 | 9.6 (9–11) | 11 (10–12) |
| Anterior end to excretory pore | 84.1 ± 3.5 (78.0–90.0) | – | 89 (81–97) | 86 (79–109) |
| Pharynx length | 103.0 ± 3.8 (96.0–112.0) | 95 | 115 (101–122) | 113 (99–127) |
| Maximum body width | 21.1 ± 2.1 (18.5–27.0) | – | – | – |
| Vulva body width | 18.9 ± 2.0 (15.5–25.0) | – | – | – |
| Post-uterine sac (PUS) length | 12.7 ± 1.5 (10.5–15.0) | – | – | – |
| Distance from vulva to anus | 120.6 ± 19.9 (102.0–178.0) | 123 | 132 (193–233) | 119 (93–146) |
| Distance from vulva to tail terminus | 262.9 ± 24.0 (238.0–317.0) | – | – | – |
| Anal body width | 12.5 ± 1.2 (10.0–14.0) | – | – | – |
| Tail length | 142.3 ± 11.9 (123.0–160.0) | 187 | 218 (193–233) | 182 (171–209) |
3.5.1. 18S Phylogeny
3.5.2. 28S Phylogeny
3.5.3. ITS Phylogeny
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrássy, I. Revision der Gattung Tylenchus Bastian, 1865 (Tylenchidae, Nematoda). Acta Zool. Hung 1954, 1, 5–42. [Google Scholar]
- Husain, S.I.; Khan, A. A new subfamily, a new subgenus and eight new species of nematodes from India belonging to superfamily Tylenchoidea. Proc. Helminthol. Soc. Wash. 1967, 34, 175–186. [Google Scholar]
- Wu, L.Y. Dactylotylenchus crassacuticulus, a new genus and new species (Tylenchinae: Nematoda). Can. J. Zool. 1968, 46, 831–834. [Google Scholar] [CrossRef]
- Brzeski, M. Lambertia uliginosa g. n. sp. n. (Nematoda, Nothotylenchidae). Nematol. Mediterr. 1977, 5, 51–55. [Google Scholar]
- Siddiqi, M.R. Seven new species in a new nematode subfamily Duosulciinae (Tylenchidae), with proposals for Duosulcius gen. n., Zanenchus gen. n. and Neomalenchus gen. n. Nematologica 1979, 25, 215–236. [Google Scholar] [CrossRef]
- Geraert, E.; Raski, D. Review of the genus Filenchus Andrassy, 1954 and descriptions of dix new species (Nemata: Tylenchidae). Nematologica 1986, 32, 265–311. [Google Scholar] [CrossRef] [Green Version]
- Siddiqi, M. Tylenchida: Parasites of Plants and Insects; CABI: Wallingford, UK, 2000; Volume 2, p. 800. [Google Scholar]
- Geraert, E. The Tylenchidae of the World: Identification of the Family Tylenchidae (Nematoda); Academia Press: Gent, Belgium, 2008; pp. 246–356. [Google Scholar]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Bert, W.; Okada, H.; Tavernier, I.; Borgonie, G.; Houthoofd, W. Morphological, morphometrical and molecular characterisation of Filenchus fungivorus n. sp., a fungivorous nematode from Japan in a most likely polyphyletic genus (Nematoda: Tylenchina). Nematology 2010, 12, 235–246. [Google Scholar] [CrossRef]
- Sawahata, T.; Nakamura, H.; Okada, H.; Sasaki, A.; Kanematsu, S. Nonlethal ectoparasitism of the mycophagous nematode Filenchus discrepans (Nematoda: Tylenchidae). Nematology 2012, 14, 159–164. [Google Scholar] [CrossRef]
- Okada, H.; Harada, H.; Kadota, I. Fungal-feeding habits of six nematode isolates in the genus Filenchus. Soil Biol. Biochem. 2005, 37, 1113–1120. [Google Scholar] [CrossRef]
- Okada, H.; Kadota, I. Host status of 10 fungal isolates for two nematode species, Filenchus misellus and Aphelenchus avenae. Soil Biol. Biochem. 2003, 35, 1601–1607. [Google Scholar] [CrossRef]
- Okada, H.; Tsukiboshi, T.; Kadota, I. Mycetophagy in Filenchus misellus (Andrássy, 1958) Lownsbery & Lownsbery, 1985 (Nematoda: Tylenchidae), with notes on its morphology. Nematology 2002, 4, 795–801. [Google Scholar]
- Munawar, M.; Yevtushenko, D.P.; Palomares-Rius, J.E.; Castillo, P. Species diversity of pin nematodes (Paratylenchus spp.) from potato growing regions of southern Alberta, Canada. Plants 2021, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.Y. Five new species of Tylenchus Bastian, 1865 (Nematoda: Tylenchidae) from the Canadian high Arctic. Can. J. Zool. 1969, 47, 1005–1010. [Google Scholar] [CrossRef]
- Brzeski, M.W. Nematodes of Tylenchina in Poland and Temperate Europe; Muzeum i Instytut Zoologii Polska Adademia Nauk: Warszawa, Poland, 1998. [Google Scholar]
- Niblack, T.L.; Bernard, E.C. Plant-parasitic nematode communities in dogwood, maple, and peach nurseries in Tennessee. J. Nematol. 1985, 17, 132–139. [Google Scholar] [PubMed]
- Thorne, G.; Malek, R.B. Nematodes of the northern Great Plains. Part I. Tylenchida (Nemata: Secernentea). Tech. Bull. S. Dak. Agric. Exp. Stn. 1968, 1, 72–76. [Google Scholar]
- Andrássy, I. Evolution as a Basis for the Systematization of Nematodes; Pitman Publishing: Budapest, Hungary, 1976; pp. 1–288. [Google Scholar]
- Khan, M.; Khan, S. Two new and a known species of Tylenchus Bastian (Nematoda: Tylenchinae) from Afghanistan. Nematol. Mediterr. 1978, 6, 213–221. [Google Scholar]
- Siddiqi, M.; Siddiqi, M.R. Tylenchida Parasites of Plants and Insects; CABI: Slough, UK, 1986; p. 646. [Google Scholar]
- Andrássy, I. Freilebende nematoden aus Angola I. Einige moosbewohnende Nematoden. Publicaçōes Cult. Da cia. de Diam. de Angola 1963, 66, 55–80. [Google Scholar]
- Jenkins, W. A rapid centrifugal-flotation technique for separating nematodes from soil. Plant Dis. Report. 1964, 48, 692. [Google Scholar]
- Seinhorst, J. On the Killing, fixation and transferring to glycerin of nematodes. Nematologica 1962, 8, 29–32. [Google Scholar] [CrossRef] [Green Version]
- De Grisse, A.T. Redescription ou modification de quelques techniques utilisees dans l’etude des nematodes phytoparasitaires. Meded. Rijksfakulteit Landbowwetenschappen Gent. 1969, 34, 351–369. [Google Scholar]
- Maria, M.; Powers, T.; Tian, Z.; Zheng, J. Distribution and description of criconematids from Hangzhou, Zhejiang Province. China. J. Nematol. 2018, 50, 183–206. [Google Scholar]
- Holterman, M.; Van der Wurff, A.; van den Elsen, S.; Van Megen, H.; Bongers, T.; Holovachov, O.; Bakker, J.; Helder, J. Phylum-Wide analysis of SSU rDNA reveals deep phylogenetic relationships among nematodes and accelerated evolution toward crown clades. Mol. Biol. Evol. 2006, 23, 1792–1800. [Google Scholar] [CrossRef]
- De Ley, P.; Felix, M.-A.; Frisse, L.; Nadler, S.; Sternberg, P.; Thomas, W.K. Molecular and morphological characterisation of two reproductively isolated species with mirror-image anatomy (Nematoda: Cephalobidae). Nematology 1999, 1, 591–612. [Google Scholar] [CrossRef]
- Ferris, V.; Ferris, J.; Faghihi, J. Variation in spacer ribosomal DNA in some cyst-forming species of plant parasitic nematodes. Fundam. Appl. Nematol. 1993, 16, 177–184. [Google Scholar]
- Curran, J.; Driver, F.; Ballard, J.; Milner, R. Phylogeny of Metarhizium: Analysis of ribosomal DNA sequence data. Mycol. Res. 1994, 98, 547–552. [Google Scholar] [CrossRef]
- Mortazavi, P.; Heydari, F.; Abolafia, J.; Castillo, P.; Pedram, M. Morphological and molecular characterization of Filenchus pseudodiscus n. sp. from east Golestan province, north Iran; with an updated phylogeny of Malenchus Andrássy, 1968 (Tylenchomorpha: Tylenchidae). J. Nematol. 2021, 53, e2021-069. [Google Scholar] [CrossRef] [PubMed]
- Munawar, M.; Yevtushenko, D.P.; Castillo, P. First report of three Tylenchidae taxa from southern Alberta, Canada. Horticulturae 2021, 7, 449. [Google Scholar] [CrossRef]
- Qing, X.; Bert, W. 3D printing in zoological systematics: Integrative taxonomy of Labrys chinensis gen. nov., sp. nov. (Nematoda: Tylenchomorpha). J. Zool. Syst. Evol. Res. 2017, 56, 35–47. [Google Scholar] [CrossRef]
- Qing, X.; Bert, W. Family Tylenchidae (Nematoda): An overview and perspectives. Org. Divers. Evol. 2019, 19, 391–408. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T. In BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Castresana, J. Selection of conserved clocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v1. 4.2, A Graphical Viewer of Phylogenetic Trees. Available online: http://tree.bio.ed.ac.uk/software/figtree/rightanglebracket (accessed on 30 December 2021).
- Elmiligy, I.A. Two New Species of Tylenchidae, Basiroides nortoni n. sp. and Tylenchus hageneri n. sp. (Nematoda: Tylenchida). J. Nematol. 1971, 3, 108–112. [Google Scholar] [PubMed]
- Geraert, E.; Zeidan, A. The genera Filenchus Andrassy, 1954, Sakia Khan, 1964, Boleodorus Thorne, 1941 and Basiria Siddiqi, 1959 (Nemata: Tylenchida) from Sudan. Nematologica 1991, 37, 185–212. [Google Scholar] [CrossRef]
- Dobrin, I.; Geraert, E. Some tylenchida from Romania. Biol. Jaarb. 1994, 62, 121–133. [Google Scholar]
- Brzeski, M. Redescription of some species of the genus Filenchus Andrássy, 1954 (Nematoda, Tylenchidae). Miscel. Zoològica 1997, 20, 45–64. [Google Scholar]
- Karegar, A.; Geraert, E. The genus Filenchus Andrássy, 1954 (Nemata: Tylenchidae) from Iran. Belg. J. Zool. 1995, 125, 363–382. [Google Scholar]
- Andrássy, I. Fauna Paraguayensis. 2. Nematoden aus den Galeriewäldern des Acaray-Flusses. Opusc. Zool. Bp. 1968, 8, 167–312. [Google Scholar]
- Bongers, T. De Nematoden Van Nederland: Een Identificatietabel Voor de in Nederland Aangetroffen Zoetwater-En Bodembewonende Nematoden; Stichting Uitgeverij Koninklijke Nederlandse Natuurhistorische Vereniging: Zeist, The Netherlands, 1988. [Google Scholar]
- Castillo, P.; Gomez-Barcina, A.; Gonzalez, M.A. Nematodes of the family Tylenchidae Orley, 1880 (Nematoda: Tylenchida), in Cazorla Mountains, Spain. Zool. Baetica. 1991, 2, 137–161. [Google Scholar]
- Atighi, M.R.; Pourjam, E.; Pereira, T.J.; Okhovvat, S.M.; Alizada, B.A.; Mundo-Ocampo, M.; Baldwin, J.G. Redescription of Filenchus annulatus (Siddiqui & Khan, 1983) Siddiqi, 1986 based on specimens from Iran with contributions to the molecular phylogeny of the Tylenchidae. Nematology 2013, 15, 129–141. [Google Scholar]
- Szczygiel, A. A new genus and four new species of the subfamily Tylenchinae de Man, 1876 (Nematoda: Tylenchidae) from Poland. Opusc. Zool. 1969, 9, 159–170. [Google Scholar]
- Andrássy, I. The genera and species of the family Tylenchidae Örley, 1880 (Nematoda). The genus Tylenchus Bastian, 1865. Acta Zool. Hung. 1979, 25, 1–33. [Google Scholar]
- Lownsbery, J.W.; Lownsbery, B.F. Plant-parasitic nematodes associated with forest trees in California. Hilgardia 1985, 53, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.; Geraert, E. Tylenchidae from Buenos Aires, Argentina. Nematologica 1996, 42, 42–61. [Google Scholar] [CrossRef]
- Andrássy, I. A Magyarországról eddig kimutatott szabadon élő fonálférgek (Nematoda) jegyzéke. Állattani Közlemények 1972, 59, 161–171. [Google Scholar]
- Gharahkhani, A.; Pourjam, E.; Abolafia, J.; Castillo, P.; Pedram, M. Sigmolenchus sinuosus n. gen., n. sp. (Tylenchidae: Ecphyadophorinae), a new member of the family. Nematology 2020, 22, 985–997. [Google Scholar] [CrossRef]
- Maqbool, M.; Shahina, F. Description of Filenchus sindhicus sp.n. and observations on three known species (Nematoda: Tylenchidae) on banana in Pakistan. Nematologica 1994, 40, 69–77. [Google Scholar] [CrossRef]
- Brzeski, M. On the taxonomic status of Tylenchus filiformis Butschli, 1873, and description of T. vulgaris sp. n. (Nematoda, Tylenchidae). Bulletin de 1′Academic Polonaise des Sciences. Cl. U. Ser. Des Sci. Biol. 1963, 11, 531–535. [Google Scholar]
- Siddiqi, M.R. The unusual position of the phasmids in Coslenchus costatus (De Man, 1921) gen. n., comb. n. and other Tylenchidae (Nematoda: Tylenchida). Nematologica 1978, 24, 449–455. [Google Scholar] [CrossRef]
- Forge, T.A.; Larney, F.J.; Kawchuk, L.M.; Pearson, D.C.; Koch, C.; Blackshaw, R.E. Crop rotation effects on Pratylenchus neglectus populations in the root zone of irrigated potatoes in southern Alberta. Can. J. Plant Pathol. 2015, 37, 363–368. [Google Scholar] [CrossRef]
- Mahran, A.; Tenuta, M.; Shinners-Carenelly, T.; Mundo-Ocampo, M.; Daayf, F. Prevalence and species identification of Pratylenchus spp. in Manitoba potato fields and host suitability of ‘Russet Burbank’. Can. J. Plant Pathol. 2010, 32, 272–282. [Google Scholar] [CrossRef]
- Munawar, M.; Yevtushenko, D.P.; Castillo, P. Integrative taxonomy, distribution, and host associations of Geocenamus brevidens and Quinisulcius capitatus from southern Alberta, Canada. J. Nematol. 2021, 53, 1–13. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munawar, M.; Castillo, P.; Yevtushenko, D.P. Description of Filenchus Species from Agroecosystem of Southern Alberta, Canada. Agronomy 2022, 12, 690. https://doi.org/10.3390/agronomy12030690
Munawar M, Castillo P, Yevtushenko DP. Description of Filenchus Species from Agroecosystem of Southern Alberta, Canada. Agronomy. 2022; 12(3):690. https://doi.org/10.3390/agronomy12030690
Chicago/Turabian StyleMunawar, Maria, Pablo Castillo, and Dmytro P. Yevtushenko. 2022. "Description of Filenchus Species from Agroecosystem of Southern Alberta, Canada" Agronomy 12, no. 3: 690. https://doi.org/10.3390/agronomy12030690
APA StyleMunawar, M., Castillo, P., & Yevtushenko, D. P. (2022). Description of Filenchus Species from Agroecosystem of Southern Alberta, Canada. Agronomy, 12(3), 690. https://doi.org/10.3390/agronomy12030690