
Variability of Root System Size and Distribution among Coffea canephora Genotypes


 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Root Traits
2.3. Statistical Analyses
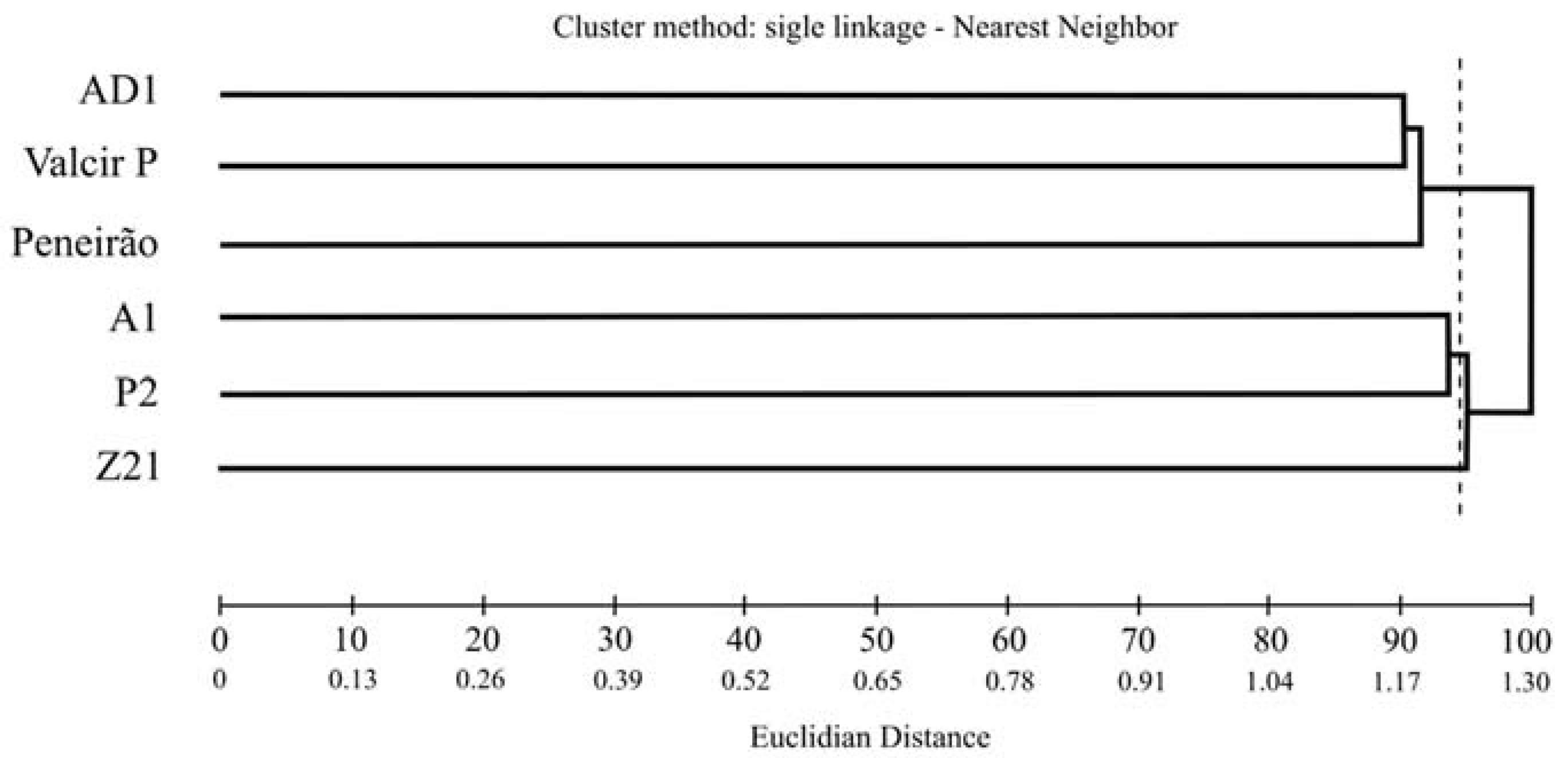
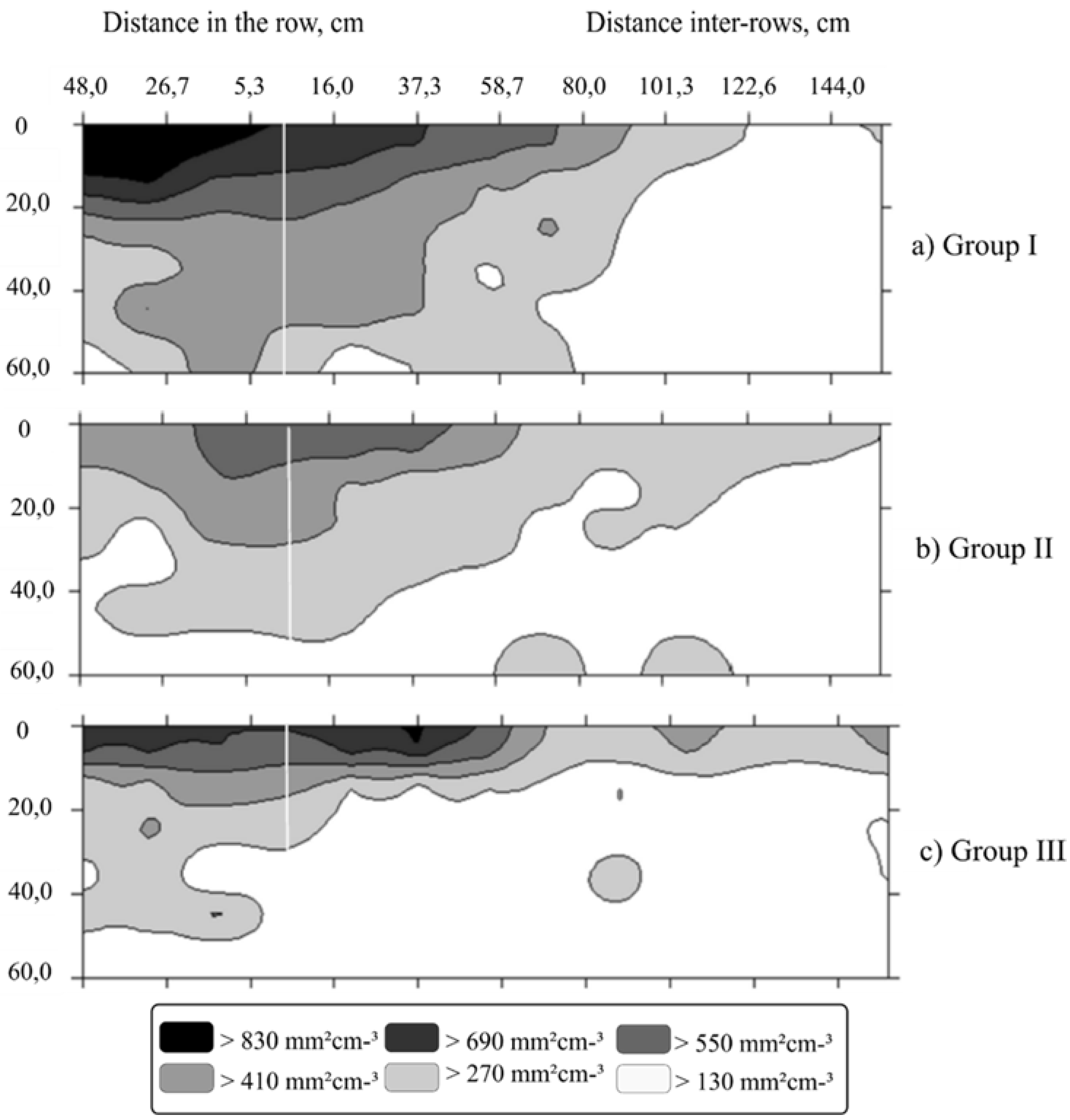
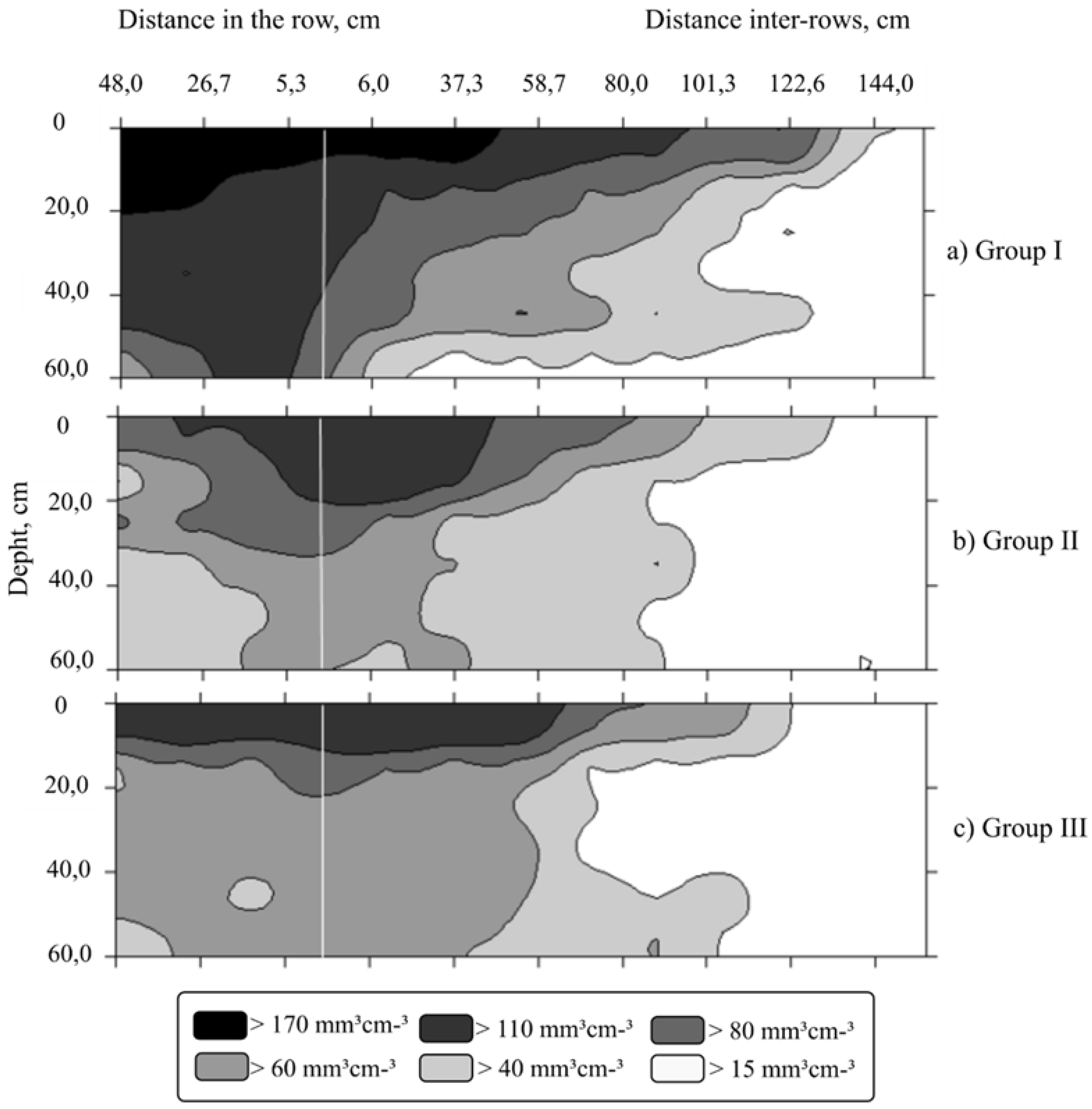
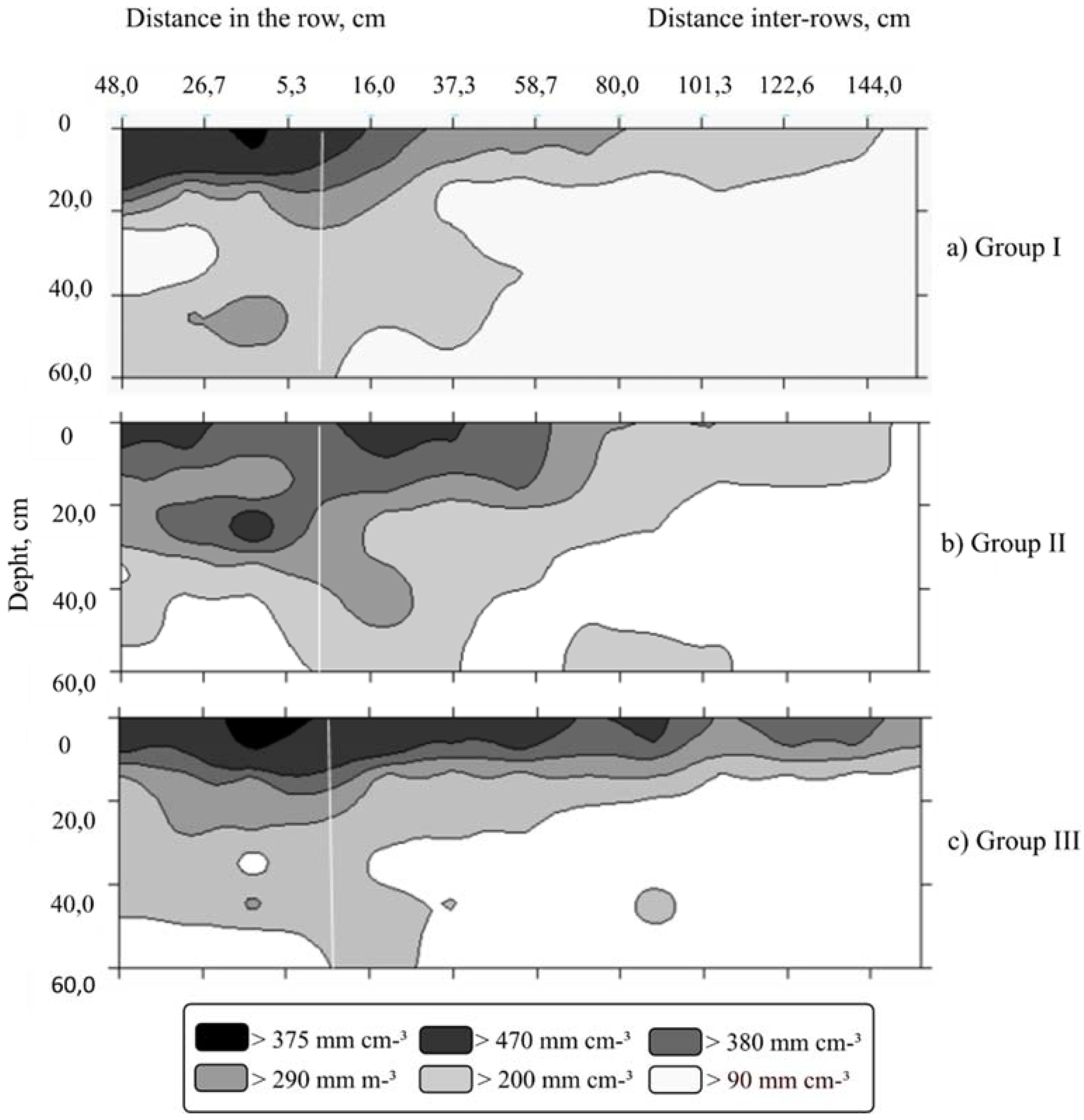
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ICO—International Coffee Organization. Global Coffee Trade. Available online: https://www.ico.org/documents/cy2021-22/cmr-1221-e.pdf (accessed on 21 January 2022).
- Conab—Companhia Nacional de Abastecimento. Acompanhamento de Safra Brasileira de Café, 6, 1–71. Available online: http://www.conab.gov.br (accessed on 10 March 2021).
- Conagin, C.H.T.M.; Mendes, A.J.T. Pesquisas citológicas e genéticas em três espécies de Coffea: Auto-incompatibilidade em Coffea canephora Pierre ex Froehner. Bragantia 1961, 20, 788–804. [Google Scholar] [CrossRef]
- Moraes, M.S.; Teixeira, A.L.; Ramalho, A.R.; Espíndula, M.C.; Ferrão, M.A.G.; Rocha, R.B. Characterization of gametophytic self-incompatibility of superior clones of Coffea canephora. Genet. Mol. Res. 2018, 17, gmr16039876. [Google Scholar] [CrossRef]
- Rodrigues, W.P.; Martins, M.Q.; Fortunato, A.S.; Rodrigues, A.P.; Semedo, J.N.; Simões-Costa, M.C.; Pais, I.P.; Leitão, A.E.; Colwell, F.; Goulao, L.; et al. Long-Term Elevated Air [CO2] Strengthens Photosynthetic Functioning and Mitigates the Impact of Supra-Optimal Temperatures in Tropical Coffea arabica and C. canephora Species. GCB Bioenergy 2016, 22, 415–431. [Google Scholar] [CrossRef]
- Dubberstein, D.; Lidon, F.C.; Rodrigues, A.N.A.P.; Semedo, J.N.; Marques, I.; Rodrigues, W.P.; Gouveia, D.D.; Armengaud, J.; Semedo, M.C.; Martins, S.; et al. Resilient and Sensitive Key Points of the Photosynthetic Machinery of Coffea spp. to the Single and Superimposed Exposure to Severe Drought and Heat Stresses. Front. Plant. Sci. 2020, 11, 1049. [Google Scholar] [CrossRef] [PubMed]
- Semedo, J.N.; Rodrigues, A.P.; Lidon, F.C.; Pais, I.P.; Marques, I.; Gouveia, D.; Armengaud, J.; Martins, S.; Semedo, M.C.; Silva, M.J.; et al. Intrinsic Non-Stomatal Resilience to Drought of the Photosynthetic Apparatus in Coffea spp. Can Be Strengthened by Elevated Air CO2. Tree Physiol. 2021, 41, 708–727. [Google Scholar] [CrossRef] [PubMed]
- Marques, I.; Gouveia, D.; Gaillard, J.C.; Martins, S.; Semedo, M.C.; Lidon, F.C.; DaMatta, F.M.; Ribeiro-Barros, A.I.; Armengaud, J.; Ramalho, J.C. Next-Generation Proteomics Reveals a Greater Antioxidative Response to Drought in Coffea arabica Than in Coffea canephora. Agronomy 2020, 12, 148. [Google Scholar] [CrossRef]
- Ramalho, J.C.; DaMatta, F.M.; Rodrigues, A.P.; Scotti-Campos, P.; Pais, I.; Batista-Santos, P.; Partelli, F.L.; Ribeiro, A.; Lidon, F.C.; Leitão, A.E. Cold Impact and Acclimation Response of Coffea spp. Plants. Theor. Exp. Plant. Physiol. 2014, 26, 5–18. [Google Scholar] [CrossRef]
- Partelli, F.L.; Golynski, A.; Ferreira, A.; Martins, M.Q.; Mauri, A.L.; Ramalho, J.C.; Vieira, H.D. Andina-First clonal cultivar of high-altitude conilon coffee. Crop. Breed. Appl. Biotechnol. 2019, 19, 476–480. [Google Scholar] [CrossRef]
- Partelli, F.L.; Giles, J.A.D.; Oliosi, G.; Covre, A.M.; Ferreira, A.; Rodrigues, V.M. Tributun: A coffee cultivar developed in partnership with farmers. Crop. Breed. Appl. Biotechnol. 2020, 20. [Google Scholar] [CrossRef]
- Senra, J.F.B.; Ferrão, M.A.G.; Mendonça, R.F.; Fonseca, A.F.A.; Ferrão, R.G.; Volpi, P.S.; Verdin Filho, A.C.; Comério, M.; Silva, M.W. Genetic Variability of Access of the Active Germplasm Bank of Coffea canephora of Incaper in Southern Espírito Santo. J. Genet. Resour. 2020, 6, 172–184. [Google Scholar] [CrossRef]
- Partelli, F.L.; Covre, A.M.; Oliveira, M.G.; Alexandre, R.S.; Vitória, E.L.; Silva, M.B. Root system distribution and yield of “Conilon” coffee propagated by seeds or cuttings. Pesqui. Agropecu. Bras. 2014, 49, 349–355. [Google Scholar] [CrossRef]
- Covre, A.M.; Partelli, F.L.; Gontijo, I.; Zucoloto, M. Root system distribution of irrigated and nonirrigated conilon coffee. Pesqui. Agropecu. Bras. 2015, 50, 1006–1016. [Google Scholar] [CrossRef]
- Voss-Fels, K.P.; Snowdon, R.J.; Hickey, L.T. Designer Roots for Future Crops. Trends Plant. Sci. 2018, 23, 957–960. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.O.E.; Schmidt, R.; Valani, G.P.; Ferreira, A.; Ribeiro-Barros, A.I.; Partelli, F.L. Root Trait Variability in Coffea canephora Genotypes and Its Relation to Plant Height and Crop Yield. Agronomy 2020, 10, 1394. [Google Scholar] [CrossRef]
- Ronchi, C.P.; Sousa Júnior, J.M.; Almeida, W.L.; Souza, D.S.; Silva, N.O.; Oliveira, L.B.; Guerra, A.M.N.M.; Ferreira, P.A. Morfologia radicular de cultivares de café arábica submetidas a diferentes arranjos espaciais. Pesqui. Agropecu. Bras. 2015, 50, 187–195. [Google Scholar] [CrossRef]
- Tracy, S.R.; Nagel, K.A.; Postma, J.A.; Fassbender, H.; Wasson, A.; Watt, M. Crop Improvement from Phenotyping Roots: Highlights Reveal Expanding Opportunities. Trends Plant. Sci. 2020, 25, 105–118. [Google Scholar] [CrossRef]
- Partelli, F.L.; Cavalcanti, A.C.C.; Menegardo, C.; Covre, A.M.; Gontijo, I.; Braun, H. Spatial distribution of the root system of Conilon and Arabica coffee plants. Pesqui. Agropecu. Bras. 2020, 55, e01333. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.M.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Santos, H.G.; Jacomine, P.K.T.; Anjos, L.H.C.; Oliveira, V.Á.; Lumbreras, J.F.; Coelho, M.R.; Almeida, J.A.; Araújo Filho, J.C.; Oliveira, J.B.; Cunha, T.J.F. Brazilian Soil Classification System, 5th ed.; Embrapa: Brasília, Brazil, 2018. [Google Scholar]
- Paye, H.S.; Partelli, F.L.; Martins, A.G.; Siebeneichler, E.A. Recomendação de adubação e calagem. In Café conilon: Conhecimento Para Superar Desafios; Partelli, F.L., Espíndula, M.C., Eds.; Caufes: Alegre, Brazil, 2019; pp. 75–97. [Google Scholar]
- Partelli, F.L.; Covre, A.M.; Oliosi, G.; Covre, D.T. Monte Pascoal: First clonal Conilon coffee cultivar for Southern Bahia-Brazil. Funct. Plant. Breed. J. 2021, 3, 107–112. [Google Scholar] [CrossRef]
- Santos, W.J.R.; Silva, B.M.; Oliveira, G.C.; Volpato, M.M.L.; Lima, J.M.; Curi, N.; Marques, J.J. Soil moisture in the root zone and its relation to plant vigor assessed by remote sensing at management scale. Geoderma 2014, 222, 91–95. [Google Scholar] [CrossRef]
- Jorge, L.A.C.; Silva, D.J.C. SAFIRA: Manual de Utilização; Embrapa Instrumentação Agropecuária: São Carlos, Brazil, 2010. [Google Scholar]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistica Computing: Viena, Austria, 2020; Available online: https://www.R-project.org (accessed on 20 February 2022).
- Vieira, S.R.; Hatfield, J.L.; Nielsen, D.R.; Biggar, J.W. Geostatistical theory and application to variability of some agronomical properties. Hilgardia 1983, 51, 1–75. [Google Scholar] [CrossRef]
- Robertson, G.P. GS+: “Geostatistics for the Environmental Sciences”; Gamma Design Software: Plainwell, MI, USA, 2008; Available online: https://geostatistics.com/files/GSPlusUserGuide.pdf (accessed on 20 February 2022).
- Amado, T.J.C.; Pontelli, C.B.; Santi, A.L.; Viana, J.H.M.; Sulzbach, L.A.S. Variabilidade espacial e temporal da produtividade de culturas sob sistema de plantio direto. Pesqui. Agropecu. Bras. 2007, 42, 1101–1110. [Google Scholar] [CrossRef]
- Mojena, R. Hierarchical grouping methods and stopping rules: An evaluation. Comput. J. 1977, 20, 359–363. [Google Scholar] [CrossRef]
- Dubberstein, D.; Partelli, F.L.; Guilhen, J.H.S.; Rodrigues, W.P.; Ramalho, J.C.; Ribeiro-Barros, A. Biometric traits as a tool for the identification and breeding of Coffea canephora genotypes. Genet. Mol. Res. 2020, 19, GMR18541. [Google Scholar] [CrossRef]
- Partelli, F.L.; Oliosi, G.; Dalazen, J.R.; Silva, C.A.; Vieira, H.D.; Espindula, M.C. Proportion of ripe fruit weight and volume to green coffee: Differences in 43 genotypes of Coffea canephora. Agron. J. 2021, 113, 1050–1057. [Google Scholar] [CrossRef]
- Salviano, A.A.C.; Vieira, S.R.; Sparovek, G. Variabilidade espacial de atributos de solo e de Crotalaria juncea L. em área severamente erodida. Rev. Bras. Cienc. Solo 1998, 22, 115–122. [Google Scholar] [CrossRef]
- Faraco, M.A.; Uribe-Opazo, M.A.; Silva, E.A.A.; Johann, J.A.; Borssoi, J.A. Seleção de modelos de variabilidade espacial para elaboração de mapas temáticos de atributos físicos do solo e produtividade da soja. Rev. Bras. Cienc. Solo 2018, 32, 463–476. [Google Scholar] [CrossRef]
- Rao, I.M.; Miles, J.W.; Beebe, S.E.; Horst, W.J. Root adaptations to soils with low fertility and aluminium toxicity. Ann. Bot. 2016, 118, 593–605. [Google Scholar] [CrossRef]
- Nunes, A.L.P.; Cortez, G.L.S.; Zaro, G.C.; Zorzenoni, T.O.; Melo, T.R.; Figueiredo, A.; Aquino, G.S.; Medina, C.C.; Ralisch, R.; Caramori, P.H.; et al. Soil morphostructural characterization and coffee root distribution under agroforestry system with Hevea Brasiliensis. Sci. Agric. 2021, 78, 1–12. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Attributes | Soil Layers (cm) | |||||
|---|---|---|---|---|---|---|
| 0–10 | 10–20 | 20–30 | 30–40 | 40–50 | 50–60 | |
| K (mg dm−3) | 110 | 95 | 74 | 57 | 52 | 46 |
| S (mg dm−3) | 15 | 11 | 29 | 15 | 15 | 17 |
| Ca (cmol dm−3) | 3.8 | 3.4 | 1.9 | 1.0 | 0.7 | 0.6 |
| Mg (cmol dm−3) | 1.0 | 0.9 | 0.4 | 0.3 | 0.1 | 0.1 |
| Al (cmol dm−3) | 0.0 | 0.0 | 0.3 | 0.7 | 0.8 | 0.8 |
| H + Al (cmol dm−3) | 1.6 | 1.8 | 2.4 | 2.9 | 3.1 | 3.1 |
| pH (H2O) | 6.6 | 6.5 | 5.3 | 4.8 | 4.8 | 4.8 |
| Mat. Org. (dag dm−3) | 2.1 | 1.7 | 1.1 | 0.8 | 0.7 | 0.5 |
| Fe (mg dm−3) | 140 | 138 | 126 | 94 | 88 | 87 |
| Zn (mg dm−3) | 10.2 | 4.5 | 2.9 | 1.1 | 0.6 | 0.5 |
| Cu (mg dm−3) | 3.4 | 4.3 | 3.0 | 1.9 | 1.2 | 1.0 |
| Mn (mg dm−3) | 207 | 174 | 104 | 46 | 44 | 40 |
| B (mg dm−3) | 0.81 | 0.83 | 0.58 | 0.55 | 0.56 | 0.61 |
| Na (mg dm−3) | 11.0 | 37.0 | 8.0 | 6.0 | 5.0 | 4.0 |
| Particle size fractions | ||||||
| Sand (g kg−1) | 434 | 352 | 188 | 368 | 366 | 376 |
| Silt (g kg−1) | 86 | 168 | 212 | 32 | 74 | 124 |
| Clay (g kg−1) | 480 | 480 | 600 | 600 | 560 | 500 |
| Parameters | Root Surface Area (mm2 cm−3) | Root Volume (mm3 cm−3) | Root Length Density (mm cm−3) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Group I | Group II | Group III | Group I | Group II | Group III | Group I | Group II | Group III | |
| SDI | 0.23 | 20.81 | 17.98 | 0.050 | 0.069 | 13.03 | 0.08 | 13.23 | 0.041 |
| R2 | 0.908 | 0.922 | 0.925 | 0.92 | 0.974 | 0.989 | 0.996 | 0.99 | 0.992 |
| RSS | 1.67 × 108 | 1.01 × 107 | 3.77 × 107 | 547,404 | 38,734.0 | 36,542 | 16,115.0 | 1,822,960 | 2,189,057 |
| RCV | 1.009 | 1.058 | 1.073 | 0.96 | 0.924 | 1.057 | 1.106 | 1.016 | 0.966 |
| Parameters | Root Surface Area (mm2 cm−3) | Root Volume (mm3 cm−3) | Root Length Density (mm cm−3) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Group I | Group II | Group III | Group I | Group II | Group III | Group I | Group II | Group III | |
| Model | Spherical | Spherical | Spherical | Spherical | Spherical | Gaussian | Spherical | Spherical | Spherical |
| Co | 100.0 | 3540.0 | 6420.0 | 1.0 | 1.0 | 222 | 10.0 | 2650.0 | 10.0 |
| Co + C | 42,621.0 | 17,010.0 | 35,700.0 | 1981.39 | 1430.0 | 1703.0 | 11,670.0 | 20,030.0 | 24,180.0 |
| Range (m) | 93.7 | 121.3 | 93.30 | 89.60 | 134.50 | 139.94 | 55.80 | 180.10 | 68.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidt, R.; Silva, L.O.E.; Ferreira, A.; Gontijo, I.; Guimarães, R.J.; Ramalho, J.C.; Partelli, F.L. Variability of Root System Size and Distribution among Coffea canephora Genotypes. Agronomy 2022, 12, 647. https://doi.org/10.3390/agronomy12030647
Schmidt R, Silva LOE, Ferreira A, Gontijo I, Guimarães RJ, Ramalho JC, Partelli FL. Variability of Root System Size and Distribution among Coffea canephora Genotypes. Agronomy. 2022; 12(3):647. https://doi.org/10.3390/agronomy12030647
Chicago/Turabian StyleSchmidt, Raquel, Larícia Olária Emerick Silva, Adesio Ferreira, Ivoney Gontijo, Rubens José Guimarães, José C. Ramalho, and Fábio Luiz Partelli. 2022. "Variability of Root System Size and Distribution among Coffea canephora Genotypes" Agronomy 12, no. 3: 647. https://doi.org/10.3390/agronomy12030647
APA StyleSchmidt, R., Silva, L. O. E., Ferreira, A., Gontijo, I., Guimarães, R. J., Ramalho, J. C., & Partelli, F. L. (2022). Variability of Root System Size and Distribution among Coffea canephora Genotypes. Agronomy, 12(3), 647. https://doi.org/10.3390/agronomy12030647