
Selection and Characterization of a Bacillus Strain for Potential Application in Industrial Production of White Button Mushroom (Agaricus bisporus)

 , ,
, ,  , and
, and
Abstract
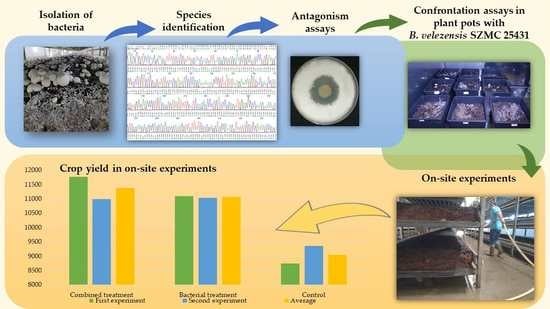
:
1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Bacillus Strains
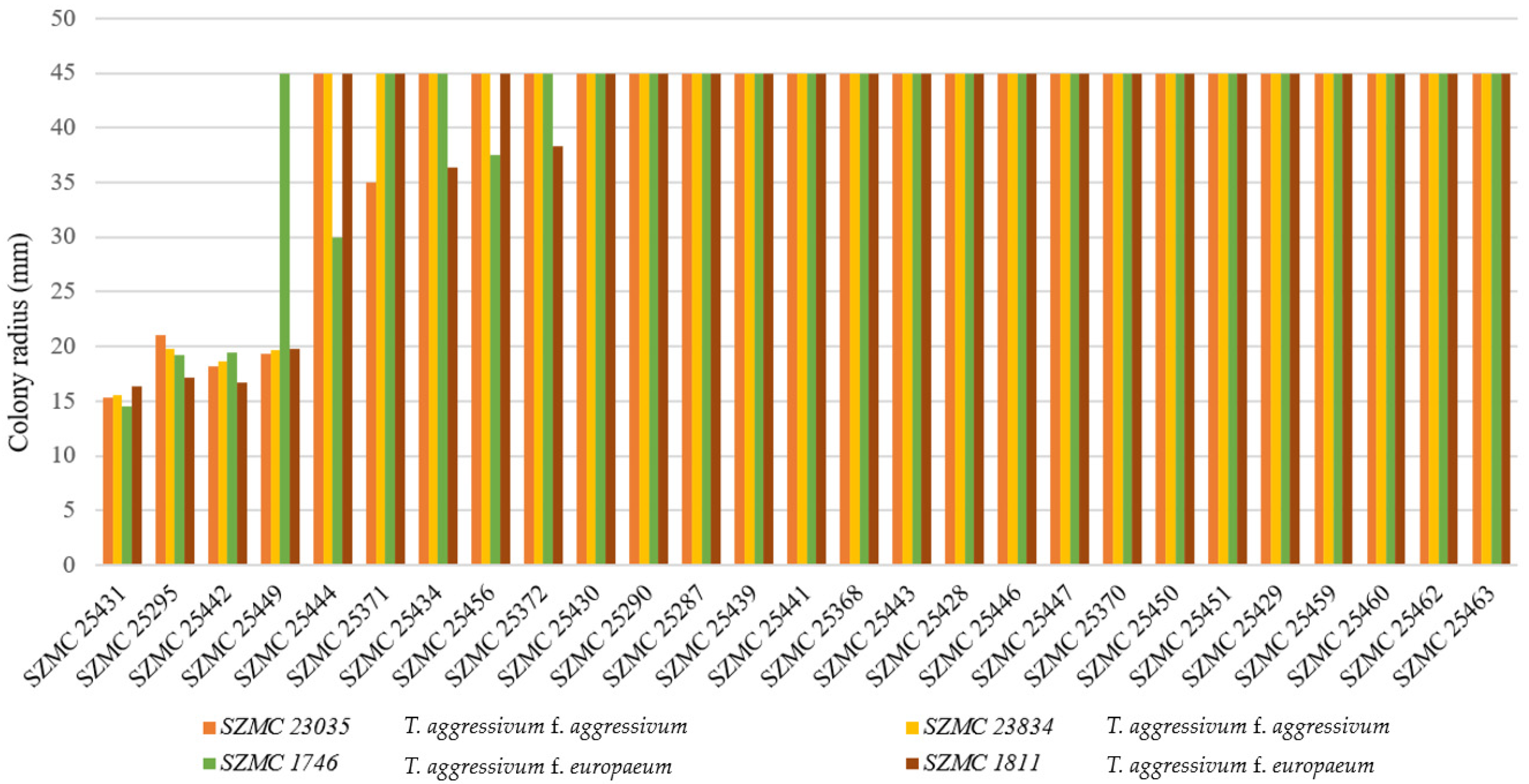
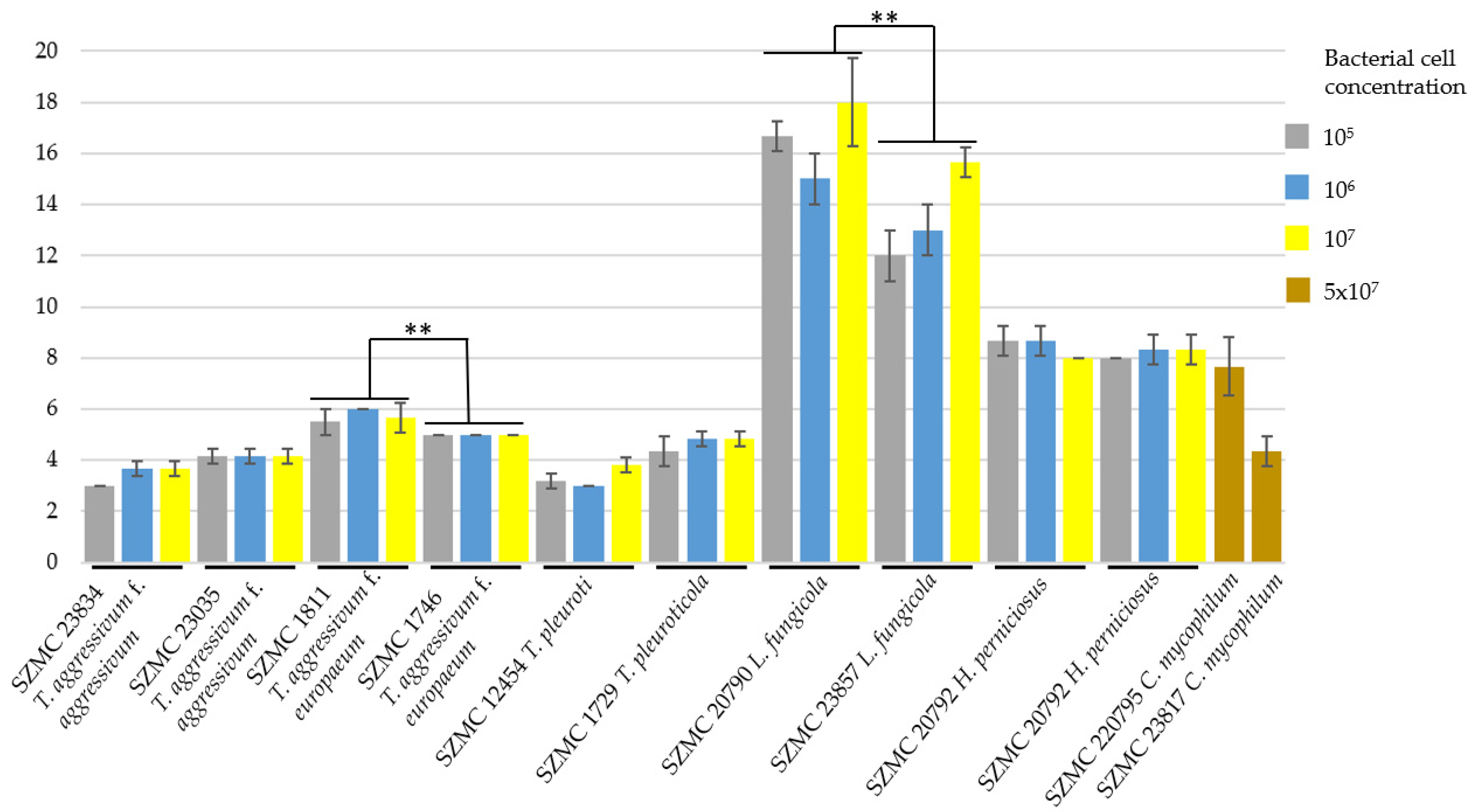
2.2. Confrontation Assays
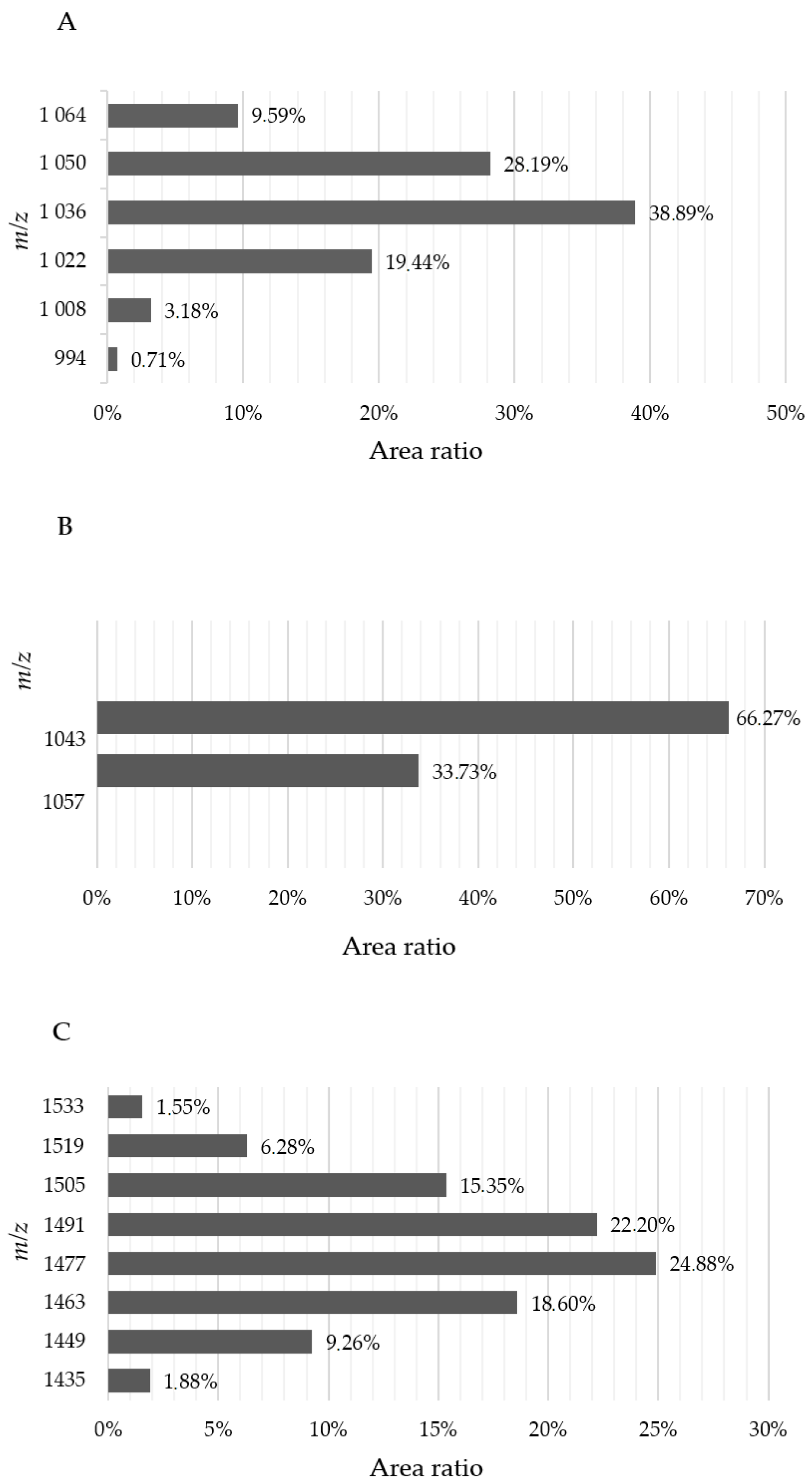
2.3. Extracellular Enzyme and Lipopeptide Production of B. velezensis SZMC 25431
2.4. Experiments in Plant Growth Chamber
2.5. Experiments in Mushroom Growing House
3. Results
3.1. Isolation and Identification of Bacillus Strains
3.2. Selection of Potential Biocontrol Strains
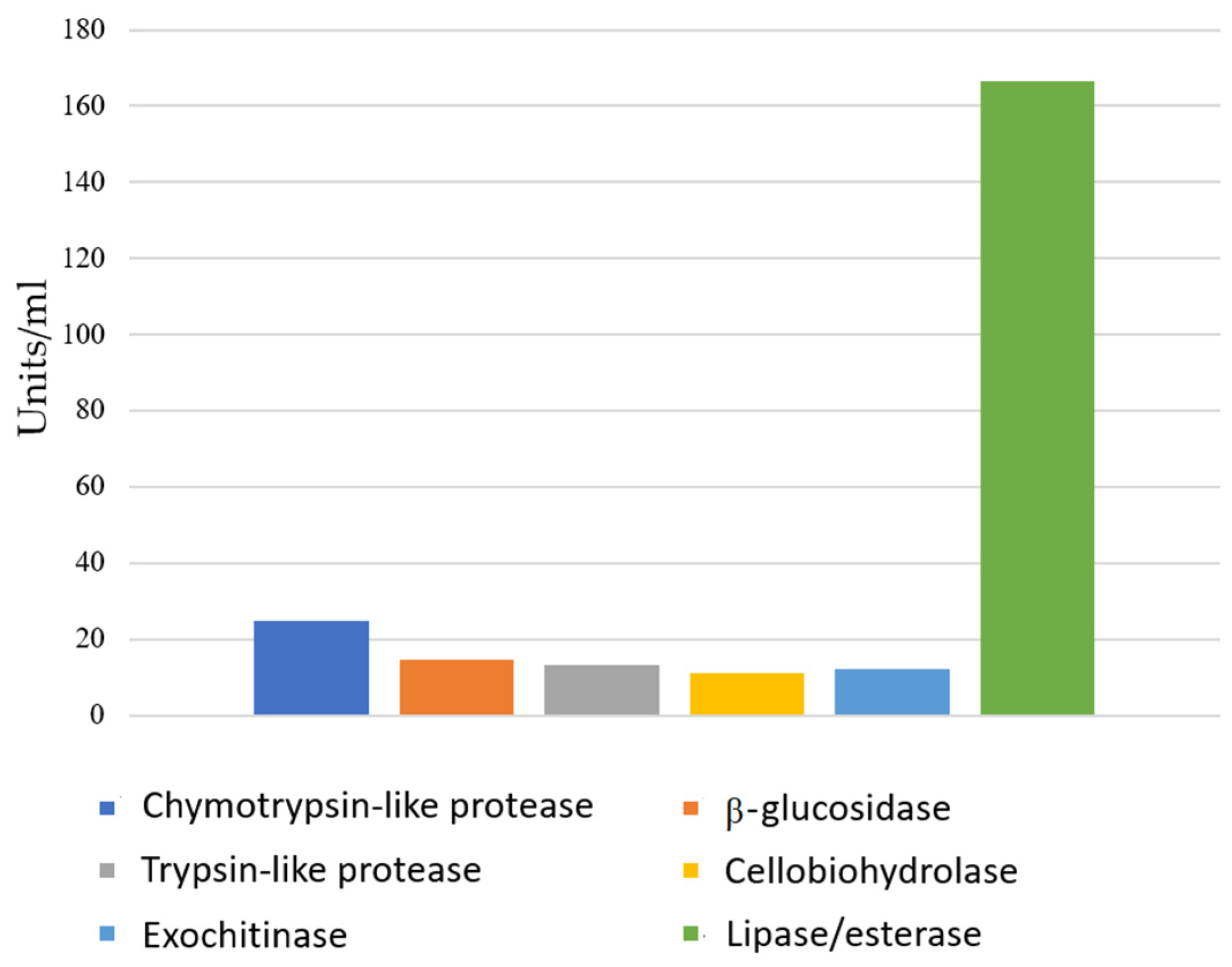
3.3. Enzyme Activities and Lipopeptide Production of B. velezensis SZMC 25431
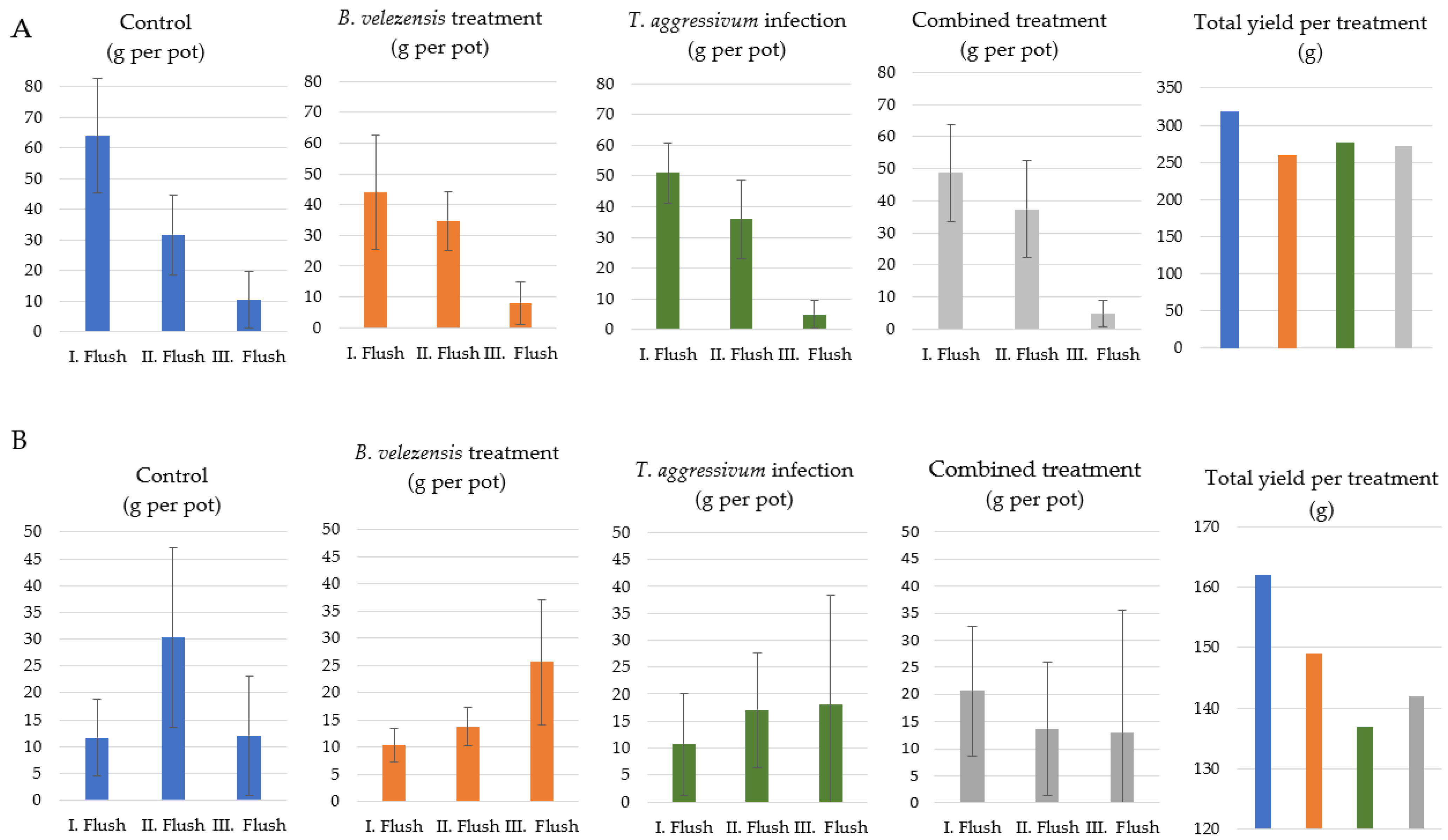
3.4. Green Mold Infection and Biocontrol Treatment in Plant Growth Chamber
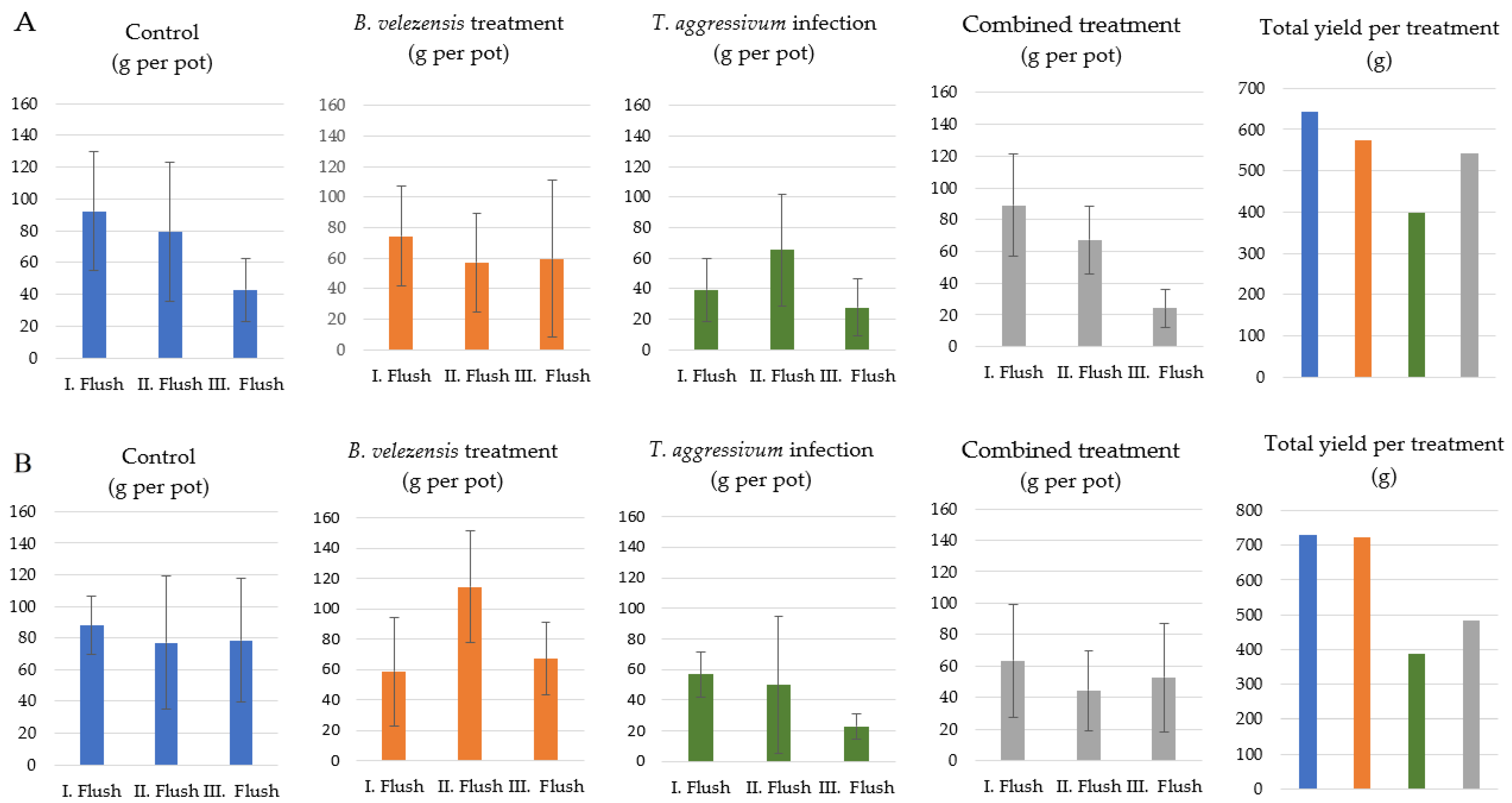
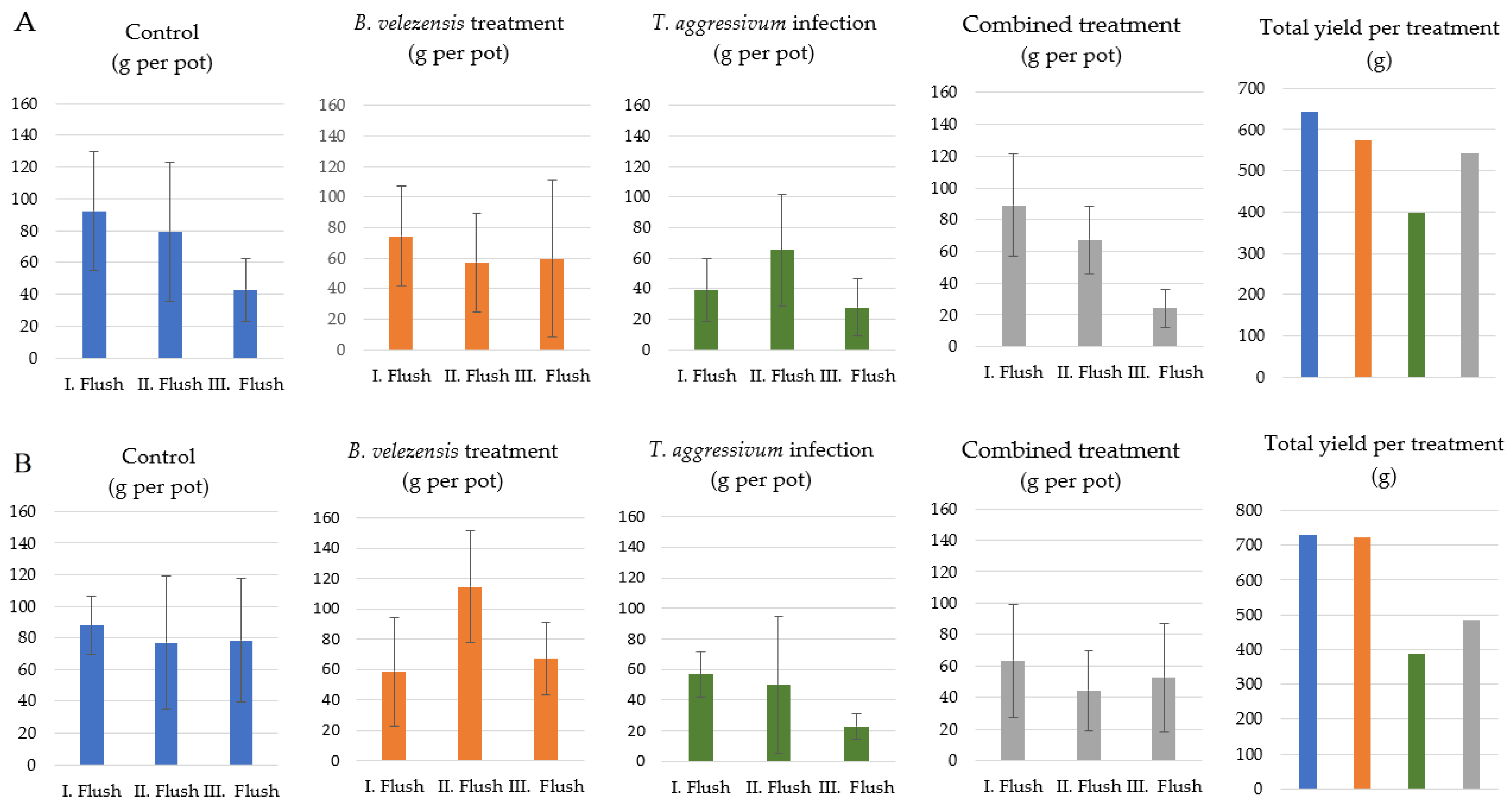
3.5. Influence of B. velezensis SZMC 25431 on the Yield of A. bisporus in Mushroom Growing Houses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hatvani, L.; Antal, Z.; Manczinger, L.; Szekeres, A.; Druzhinina, I.S.; Kubicek, C.P.; Nagy, A.; Nagy, E.; Vágvölgyi, C.; Kredics, L. Green mold disease of Agaricus bisporus and Pleurotus spp. are caused by related but phylogenetically different Trichoderma species. Phytopathology 2007, 97, 532–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Largeteau, M.L.; Savoie, J.M. Microbially induced diseases of Agaricus bisporus: Biochemical mechanisms and impact on commercial mushroom production. Appl. Microbiol. Biotechnol. 2010, 86, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Kredics, L.; García-Jimenez, L.; Naeimi, S.; Czifra, D.; Urbán, P.; Manczinger, L.; Vágvölgyi, C.; Hatvani, L. A challenge to mushroom growers: The green mould disease of cultivated champignons. In Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Mendez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2010; pp. 295–305. ISBN 978-84-614-6194-3. [Google Scholar]
- Kredics, L.; Naeimi, S.; Hatvani, L.; Vágvölgyi, C.; Cai, F.; Druzhinina, I.S.; Manczinger, L. ‘The Good, the Bad and the Ugly’ in the shades of green: The genus Trichoderma in the spotlight. Indian Phytopathol. 2021, 74, 403–411. [Google Scholar] [CrossRef]
- Samuels, G.; Dodd, S.; Castlebury, L.; Petrini, O. Trichoderma species associated with the green mold epidemic of commercially grown Agaricus bisporus. Mycologia 2002, 94, 146–170. [Google Scholar] [CrossRef]
- Grogan, H.M.; Noble, R.; Gaze, R.H.; Fletcher, J.T. Control of Trichoderma harzianum—A weed mould of mushroom cultivation. In Proceedings of the Brighton Crop Protection Conference, Brighton, UK, 18–21 November 1996; Volume 1, pp. 337–342, ISBN 0-948404-99-X. [Google Scholar]
- Hatvani, L.; Kredics, L.; Allaga, H.; Manczinger, L.; Vágvölgyi, C.; Kuti, K.; Geösel, A. First report of Trichoderma aggressivum f. aggressivum green mold on Agaricus bisporus in Europe. Plant Dis. 2017, 101, 1052–1056. [Google Scholar] [CrossRef] [Green Version]
- Clarke, J.; Grogan, H.; Fitzpatrick, D.; Kavanagh, K. Analysis of the effect of Bacillus velezensis culture filtrate on the growth and proteome of Cladobotryum mycophilum. Fungal Biol. 2022, 126, 11–19. [Google Scholar] [CrossRef]
- White, P.F.; Smith, J. Pests. In Mushroom Pest and Disease Control: A Colour Handbook; Fletcher, J., Gaze, R., Eds.; Manson Publishing: London, UK, 2007; pp. 140–159. ISBN 978-1-84076-083-4. [Google Scholar]
- Carrasco, J.; Navarro, M.J.; Gea, F.J. Cobweb, a serious pathology in mushroom crops: A review. Spanish J. Agricult. 2017, 15, e10R01. [Google Scholar] [CrossRef] [Green Version]
- Lakkireddy, K.; Khonsuntia, W.; Kües, U. Mycoparasite Hypomyces odoratus infests Agaricus xanthodermus fruiting bodies in nature. AMB Express 2020, 10, 141. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Baars, J.J.; Kalkhove, S.I.C.; Lugones, L.G.; Wösten, H.A.B.; Bakker, P.A.H.M. Lecanicillium fungicola: Causal agent of dry bubble disease in white-button mushroom. Mol. Plant. Pathol. 2010, 11, 585–595. [Google Scholar] [CrossRef]
- Soković, M.; Van Griensven, L.J.L.D. Antimicrobial activity of essential oils and their components against the three major pathogens of the cultivated button mushroom, Agaricus bisporus. Eur. J. Plant Pathol. 2006, 116, 211–224. [Google Scholar] [CrossRef]
- Mills, P.R.; Thomas, J.; Sergeant, M.R.; Costa, A.; Collopy, P.D.; Bailey, A.M.; Foster, G.D.; Challen, M.P. Interactions between Agaricus bisporus and the pathogen Verticillium fungicola. In Stress in Yeasts and Filamentous Fungi, 1st ed.; Avery, S.V., Stratford, M., van West, P., Eds.; Academic Press: London, UK, 2008; Volume 27, pp. 1–17. ISBN 978-0-12-374184-4. [Google Scholar]
- Kouser, S.; Shah, S.; Ahmed, M.; Shah, M.D.; Sheikh, P.A. Morphological characteristics of wet bubble disease (Mycogone perniciosa) isolated from button mushroom (Agaricus bisporus) and assessment of factors affecting disease development and spread. Afr. J. Microbiol. Res. 2015, 9, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Romaine, C.P.; Royse, D.J.; Schlagnhaufer, C. Superpathogenic Trichoderma resistant to topsin M found in Pennsylvania and Delaware. Mushr. News 2005, 53, 6–9. [Google Scholar]
- Pecchia, J.A.; Beyer, D.M. Pest management on US commercial mushroom farms. Outlooks Pest Manag. 2013, 24, 28–29. [Google Scholar] [CrossRef]
- Dhar, B.; Ahlawat, O.P.; Sharma, R.K.; Dubey, J.K.; Patiyal, S.K.; Thakur, M. Organic button mushroom (Agaricus bisporus) production, quality produce and pesticide residue analysis. In Mushroom Biology and Mushroom Products, Proceedings of the Sixth International Conference on Mushroom Biology and Mushroom Products, Bonn, Germany, 29 September–3 October 2008; Lelley, J.I., Buswell, J.A., Eds.; Gamu GmbH, Institut für Pilzforschung: Krefeld, Germany, 2008; pp. 203–211. [Google Scholar]
- Hjeljord, L.; Tronsmo, A. Trichoderma and Gliocladium in biological control: An overview. In Trichoderma and Gliocladium, Enzymes, Biological Control and Commercial Applications, 1st ed.; Harman, G.E., Kubicek, C.P., Eds.; CRC Press: Boca Raton, FL, USA, 1998; Volume 2. [Google Scholar] [CrossRef]
- Grogan, H.M.; Gaze, R.H. Fungicide resistance among Cladobotryum spp.—Causal agents of cobweb disease of the edible mushroom Agaricus bisporus. Mycol. Res. 2000, 104, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Kosanović, D.; Potočnik, I.; Vukojević, J.; Stajić, M.; Rekanović, E.; Stepanović, M.; Todorović, B. Fungicide sensitivity of Trichoderma spp. from Agaricus bisporus farms in Serbia. J. Environ. Sci. Health Part B 2015, 50, 607–613. [Google Scholar] [CrossRef]
- Lewis, K.A.; Tzilivakis, J.; Warner, D.; Green, A. An international database for pesticide risk assessments and management. Hum. Ecol. Risk Assess. 2016, 22, 1050–1064. [Google Scholar] [CrossRef] [Green Version]
- European Commission, EFSA (European Food Safety Authority). Technical report on the outcome of the consultation with Member States and EFSA on the basic substance application for (sea) salt (sodium chloride) for use in plant protection as fungicide and insecticide. EFSA Supporting Publ. 2017, 14, 56. [Google Scholar] [CrossRef] [Green Version]
- Di Poi, C.; Costil, K.; Bouchart, V.; Halm-Lemeille, M.P. Toxicity assessment of five emerging pollutants, alone and in binary or ternary mixtures, towards three aquatic organisms. Environ. Sci. Pollut. Res. 2017, 25, 6122–6134. [Google Scholar] [CrossRef] [Green Version]
- Uwizeyimana, H.; Wang, M.; Chen, W.; Khan, K. The eco-toxic effects of pesticide and heavy metal mixtures towards earthworms in soil. Environ. Toxicol. Pharmacol. 2017, 55, 20–29. [Google Scholar] [CrossRef]
- Fravel, D.R. Commercialization and implementation of biocontrol. Annu. Rev. Phytopathol. 2005, 43, 337–359. [Google Scholar] [CrossRef]
- Thakore, Y. The biopesticide market for global agricultural use. Ind. Biotech. 2006, 2, 194–208. [Google Scholar] [CrossRef]
- Cawoy, H.; Bettiol, W.; Fickers, P.; Ongena, M. Bacillus-based Biological Control of Plant Diseases. In Pesticides in the Modern World: Pesticides Use and Management; Stoytcheva, M., Ed.; InTechOpen: London, UK, 2011; pp. 273–302. [Google Scholar] [CrossRef] [Green Version]
- Stanojević, O.; Berić, T.; Potočnik, I.; Rekanović, E.; Stanković, S.; Milijasević-Marić, S. Biological control of green mould and dry bubble diseases of cultivated mushroom (Agaricus bisporus L.) by Bacillus spp. Crop Prot. 2019, 126, 104944. [Google Scholar] [CrossRef]
- Yánez-Mendizábal, V.; Viñas, I.; Usall, J.; Torres, R.; Solsona, C.; Teixidó, N. Production of the postharvest biocontrol agent Bacillus subtilis CPA-8 using low cost commercial products and by-products. Biol. Control 2012, 60, 280–289. [Google Scholar] [CrossRef]
- Cai, W.M.; Yao, H.Y.; Feng, W.L.; Jin, Q.L.; Liu, Y.Y.; Li, N.Y.; Zheng, Z. Microbial community structure of casing soil during mushroom growth. Pedosphere 2009, 19, 446–452. [Google Scholar] [CrossRef]
- Kim, W.G.; Weon, H.Y.; Seok, S.J.; Lee, K.H. In vitro antagonistic characteristics of bacilli isolates against Trichoderma spp. and three species of mushrooms. Mycobiology 2008, 36, 266–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandin, C.; Le Coq, D.; Deschamps, J.; Védie, R.; Rousseau, T.; Aymerich, S.; Briandet, R. Complete genome sequence of Bacillus velezensis QST713: A biocontrol agent that protects Agaricus bisporus crops against the green mould disease. J. Biotechnol. 2018, 278, 10–19. [Google Scholar] [CrossRef]
- Potočnik, I.; Todorović, B.; Rekanović, E.; Luković, J.; Paunović, D.; Milijašević-Marčić, S. Impact of Bacillus subtilis QST713 mushroom grain spawn treatment on yield and green mould control. Pestic. Phytomed. 2018, 33, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Potočnik, I.; Rekanović, E.; Todorović, B.; Luković, J.; Paunović, D.; Stanojević, O.; Milijašević-Marčić, S. The effects of casing soil treatment with Bacillus subtilis Ch-13 biofungicide on green mould control and mushroom yield. Pest. Phytomed. 2019, 34, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Anastassiadou, M.; Arena, M.; Auteri, D.; Brancato, A.; Bura, L.; Carrasco Cabrera, L.; Chaideftou, E.; Chiusolo, A.; Crivellente, F.; De Lentdecker, C.; et al. Peer review of the pesticide risk assessment of the active substance Bacillus amyloliquefaciens strain QST 713 (formerly Bacillus subtilis strain QST 713). EFSA J. 2021, 19, 6381–6401. [Google Scholar] [CrossRef]
- Aydoğdu, M.; Sülü, S.M.; Kurbetli, İ.; Sülü, G. In vitro and in vivo biological control of the green mold using different bacteria in button mushroom cultivation. Egypt. J. Biol. Pest Cont. 2021, 31, 1–11. [Google Scholar] [CrossRef]
- Bhat, M.A.; Simon, S.; Munshi, N.A.; Bhat, Z.A. In vitro efficacy of casing and compost isolated bacterial inoculants against Verticillium fungicola (Preuss) Hassebrauk and Agaricus bisporus (Lange) Imbach. J. Biol. Control 2010, 24, 137–141. [Google Scholar] [CrossRef]
- Drancourt, M.; Bollet, C.; Carlioz, A.; Martelin, R.; Gayral, J.P.; Raoult, D. 16S ribosomal DNA sequence analysis of a large collection of environmental and clinical unidentifiable bacterial isolates. J. Clin. Microbiol. 2000, 38, 3623–3630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.-H.; Park, B.-S.; Kim, H.-Y.; Lee, K.-H.; Kim, K.S. Antagonistic and plant growth-promoting effects of Bacillus velezensis BS1 isolated from rhizosphere soil in a pepper field. Plant Pathol. J. 2021, 37, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Ki, J.S.; Zhang, W.; Qian, P.Y. Discovery of marine Bacillus species by 16S rRNA and rpoB comparisons and their usefulness for species identification. J. Microbiol. Meth. 2009, 77, 48–57. [Google Scholar] [CrossRef]
- NCBI Blast, Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. 2004. Available online: https://www.blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 29 December 2021).
- Kartali, T.; Shahab, D.; Nyilasi, I.; Hatvani, L.; Kredics, L.; Vágvölgyi, C.; Papp, T. Double-stranded RNA elements in Lecanicil-lium and Mycogene strains infecting Agaricus bisporus. Mikol. Közl. Clusiana 2017, 56, 103–105. [Google Scholar]
- Kedves, O.; Kocsubé, S.; Bata, T.; Andersson, M.A.; Salo, J.M.; Mikkola, R.; Salonen, H.; Szűcs, A.; Kedves, A.; Kónya, Z.; et al. Chaetomium and Chaetomium-like species from European indoor environments include Dichotomopilus finlandicus sp. nov. Pathogens 2021, 10, 1133. [Google Scholar] [CrossRef] [PubMed]
- Allaga, H.; Büchner, R.; Hatvani, L.; Szekeres, A.; Vágvölgyi, C.; Kredics, L.; Manczinger, L. Adaptation of a plant growth chamber for the experimental cultivation of champignons (Agaricus bisporus). In Proceedings of the 21st Danube-Kris-Mures-Tisza (DKMT) Euroregional Conference on Environment and Health, Novi Sad, Serbia, 6–8 June 2019; Škrbić, B., Ed.; University of Novi Sad, Faculty of Technology: Novi Sad, Serbia, 2019; pp. 51–59. [Google Scholar]
- Zied, D.C.; Minhoni, M.T.A.; Pardo-Gonzalez, J.E.; Pardo-Gimenez, A. A study of compost added to a casing technique in Agaricus bisporus cultivation from phase III bulk compost. Hort. Sci. 2010, 45, 1649–1653. [Google Scholar] [CrossRef] [Green Version]
- Pertot, I.; Alabouvette, C.; Esteve, H.E.; França, S. Mini-Paper—The Use of Microbial Biocontrol Agents against Soil-Borne Diseases; Eip-Agri: Brussels, Belgium, 2015; Available online: http://ec.europa.eu/eip/agriculture/sites/agrieip/files/8_eip_sbd_mp_biocontrol_final.pdf (accessed on 9 February 2022).
- Pandin, C.; Darsonval, M.; Mayeur, C.; Le Coq, D.; Aymerich, S.; Romain, B. Biofilm formation and synthesis of antimicrobial compounds by the biocontrol agent Bacillus velezensis QST713 in an Agaricus bisporus compost micromodel. Appl. Environ. Microbiol. 2019, 85, e00327-19. [Google Scholar] [CrossRef] [Green Version]
- Milijašević-Marčić, S.; Stepanović, S.; Todorović, B.; Duduk, B.; Stepanović, J.; Rekanović, E.; Potočnik, I. Biological control of green mould on Agaricus bisporus by a native Bacillus subtilis strain from mushroom compost. Eur. J. Plant. Pathol. 2017, 148, 509–519. [Google Scholar] [CrossRef]
- Liu, C.; Sheng, J.; Chen, L.; Zheng, Y.; Lee, D.Y.W.; Yang, Y.; Xu, M.; Shen, L. Biocontrol activity of Bacillus subtilis isolated from Agaricus bisporus mushroom compost against pathogenic fungi. J. Agricul. Food Chem. 2015, 63, 6009–6018. [Google Scholar] [CrossRef]
- Velázquez-Cedeño, M.; Farnet, A.M.; Mata, G.; Savoie, J.M. Role of Bacillus spp. in antagonism between Pleurotus ostreatus and Trichoderma harzianum in heat-treated wheat-straw substrates. Bioresour. Technol. 2008, 99, 6966–6973. [Google Scholar] [CrossRef] [PubMed]
- Devi, S.; Kiesewalter, H.T.; Kovács, R.; Frisvad, J.C.; Weber, T.; Larsen, T.O.; Kovács, Á.T.; Ding, L. Depiction of secondary metabolites and antifungal activity of Bacillus velezensis DTU001. Synth. Syst. Biotechnol. 2019, 4, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.Q.; Li, Q.L.; Hu, J.C. The complete genome sequence of Bacillus velezensis 9912D reveals its biocontrol mechanism as a novel commercial biological fungicide agent. J. Biotechnol. 2017, 247, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Bóka, B.; Manczinger, L.; Kecskeméti, A.; Chandrasekaran, M.; Kadaikunnan, S.; Alharbi, N.S.; Vágvölgyi, C.; Szekeres, A. Ion trap mass spectrometry of surfactins produced by Bacillus subtilis SZMC 6179J reveals novel fragmentation features of cyclic lipopeptides. Rapid Commun. Mass Spectrom. 2016, 30, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Kecskeméti, A.; Bartal, A.; Bóka, B.; Kredics, L.; Manczinger, L.; Shine, K.; Alharby, N.S.; Khaled, J.M.; Varga, M.; Vágvölgyi, C.; et al. High-frequency occurrence of surfactin monomethyl isoforms in the ferment broth of a Bacillus subtilis strain revealed by ion trap mass spectrometry. Molecules 2018, 23, 2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Li, X.; Li, X.; Yu, H.; Shen, Z. Identification of lipopeptide isoforms by MALDI-TOF-MS/MS based on the simultaneous purification of iturin, fengycin, and surfactin by RP-HPLC. Anal. Bioanal. Chem. 2015, 407, 2529–2542. [Google Scholar] [CrossRef]
- Calvo, O.C.; Quaglia, G.; Mohiley, A.; Cesarini, M.; Fangmeier, A. Assessing potential aquatic toxicity of airport runoff using physicochemical parameters and Lemna gibba and Aliivibrio fischeri bioassays. Environ. Sci. Pollut. Res. 2020, 27, 40604–40617. [Google Scholar] [CrossRef]
- Yan, Y.; Ziemek, J.; Schetelig, M.F. CRISPR/Cas9 mediated disruption of the white gene leads to pigmentation deficiency and copulation failure in Drosophila suzukii. J. Insect Physiol. 2020, 126, 104091. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | SZMC Number | Concentration (Conidia mL−1) |
|---|---|---|
| Trichoderma aggressivum f. aggressivum | 23834 | 5 × 105 |
| T. aggressivum f. aggressivum | 23035 | 5 × 105 |
| T. aggressivum f. europaeum | 1811 | 5 × 105 |
| T. aggressivum f. europaeum | 1746 | 5 × 105 |
| T. pleuroti | 12454 | 5 × 105 |
| T. pleuroticola | 1729 | 5 × 105 |
| Lecanicillium fungicola | 20790 | 2 × 105 |
| L. fungicola * | 23857 | 2 × 105 |
| Cladobotryum mycophilum | 20795 | 106 |
| C. mycophilum | 23817 | 106 |
| Hypomyces perniciosus | 20792 | 5 × 105 |
| H. perniciosus | 20793 | 5 × 105 |
| Experiment | Concentration * of B. velezensis SZMC 25431 Suspension | Concentration * of B. velezensis SZMC 25431 Cells g−1 of Casing | Concentration * of T. aggressivum f. aggressivum SZMC 23834 Suspension | Concentration * of T. aggressivum f. aggressivum SZMC 23834 Conidia g−1 of Casing |
|---|---|---|---|---|
| 1. | 107 | 106 | 106 | 105 |
| 2. | 106 | 105 | 107 | 106 |
| 3. | 105 | 104 | 107 | 106 |
| 4. | 105 | 104 | 107 | 106 |
| First Flush | Second Flush | Total Yield | Difference Compared to Fungicide Treatment | |||||
|---|---|---|---|---|---|---|---|---|
| I. | (kg) | (kg/m2) | (kg) | (kg/m2) | (kg) | (kg/m2) | (kg) | (%) |
| Combined treatment | 6480 | 13.5 | 5280 | 11 | 11760 | 24.5 | +3024 | +34.6 |
| Bacterial treatment | 6096 | 12.7 | 4992 | 10.4 | 11088 | 23.1 | +2352 | +26 |
| Fungicide treatment | 5856 | 12.2 | 2880 | 6 | 8736 | 18.26 | - | - |
| II. | ||||||||
| Combined treatment | 6096 | 12.7 | 4896 | 10.2 | 10992 | 22.9 | +1632 | +17.4 |
| Bacterial treatment | 5904 | 12.3 | 5184 | 10.8 | 11040 | 23.0 | +1680 | +17.9 |
| Fungicide treatment | N/A * | N/A | N/A | N/A | 9360 | 19.5 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Büchner, R.; Vörös, M.; Allaga, H.; Varga, A.; Bartal, A.; Szekeres, A.; Varga, S.; Bajzát, J.; Bakos-Barczi, N.; Misz, A.; et al. Selection and Characterization of a Bacillus Strain for Potential Application in Industrial Production of White Button Mushroom (Agaricus bisporus). Agronomy 2022, 12, 467. https://doi.org/10.3390/agronomy12020467
Büchner R, Vörös M, Allaga H, Varga A, Bartal A, Szekeres A, Varga S, Bajzát J, Bakos-Barczi N, Misz A, et al. Selection and Characterization of a Bacillus Strain for Potential Application in Industrial Production of White Button Mushroom (Agaricus bisporus). Agronomy. 2022; 12(2):467. https://doi.org/10.3390/agronomy12020467
Chicago/Turabian StyleBüchner, Rita, Mónika Vörös, Henrietta Allaga, András Varga, Attila Bartal, András Szekeres, Sarolta Varga, Judit Bajzát, Nóra Bakos-Barczi, András Misz, and et al. 2022. "Selection and Characterization of a Bacillus Strain for Potential Application in Industrial Production of White Button Mushroom (Agaricus bisporus)" Agronomy 12, no. 2: 467. https://doi.org/10.3390/agronomy12020467
APA StyleBüchner, R., Vörös, M., Allaga, H., Varga, A., Bartal, A., Szekeres, A., Varga, S., Bajzát, J., Bakos-Barczi, N., Misz, A., Csutorás, C., Hatvani, L., Vágvölgyi, C., & Kredics, L. (2022). Selection and Characterization of a Bacillus Strain for Potential Application in Industrial Production of White Button Mushroom (Agaricus bisporus). Agronomy, 12(2), 467. https://doi.org/10.3390/agronomy12020467