
Use of Agro-Waste as a Source of Crop Nutrients in Intensive Horticulture System

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Experiment Design
2.2. Sampling
2.3. Statistical Analyses
3. Results and Discussion
3.1. Yield
3.2. Sap Analysis
3.3. Microorganisms
3.4. Enzymatic Activity
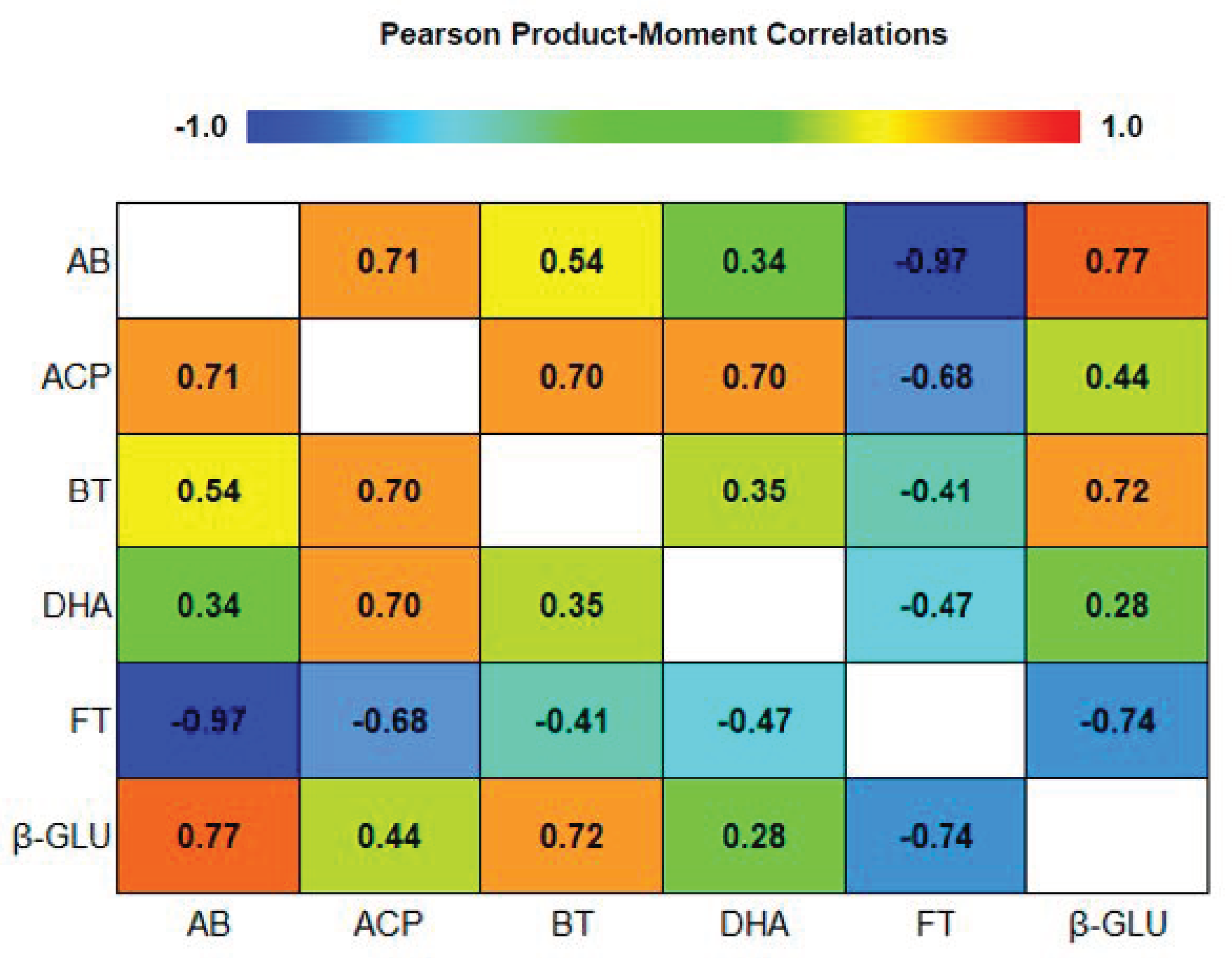
3.5. Pearson Product Moment Correlations among Soil Biological Proprieties
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Demirbas, A. Waste management, waste resource facilities and waste conversion processes. Energy Convers. Manag. 2011, 52, 1280–1287. [Google Scholar] [CrossRef]
- Aznar-Sánchez, J.A.; Velasco-Muñoz, J.F.; García-Arca, D.; López-Felices, B. Identification of opportunities for applying the circular economy to intensive agriculture in Almería (South-East Spain). Agronomy 2020, 10, 1499. [Google Scholar] [CrossRef]
- Lopez, M.J.; Masaguer, A.; Paredes, C.; Perez, L.; Muñoz, M.; Salas, M.C.; Hernandez, R. De resíduos a recursos. El camino hacia la Sostenibilidad. Red Española Compost. 2015, 91–121. [Google Scholar]
- Carricondo-Martínez, I.; Berti, F.; Salas-Sanjuán, M.C. Different Organic Fertilization Systems Modify Tomato Quality: An Opportunity for Circular Fertilization in Intensive Horticulture. Agronomy 2022, 12, 174. [Google Scholar] [CrossRef]
- Janssen, B.H.; Oenema, O. Global economics of nutrient cycling. Turk. J. Agric. For. 2008, 32, 165–176. [Google Scholar]
- Cambier, P.; Pot, V.; Mercier, V.; Michaud, A.; Benoit, P.; Revallier, A.; Houot, S. Impact of long-term organic residue recycling in agriculture on soil solution composition and trace metal leaching in soils. Sci. Total Environ. 2014, 499, 560–573. [Google Scholar] [CrossRef]
- Williams, H.; Colombi, T.; Keller, T. The influence of soil management on soil health: An on-farm study in southern Sweden. Geoderma 2020, 360, 114010. [Google Scholar] [CrossRef]
- Hernández, T.; Chocano, C.; Moreno, J.L.; García, C. Use of compost as an alternative to conventional inorganic fertilizers in intensive lettuce (Lactuca sativa L.) crops—Effects on soil and plant. Soil Tillage Res. 2016, 160, 14–22. [Google Scholar] [CrossRef]
- Carrera, L.M.; Buyer, J.S.; Vinyard, B.; Abdul-Baki, A.A.; Sikora, L.J.; Teasdale, J.R. Effects of cover crops, compost, and manure amendments on soil microbial community structure in tomato production systems. Appl. Soil Ecol. 2007, 37, 247–255. [Google Scholar] [CrossRef]
- Liang, B.; Zhao, W.; Yang, X.; Zhou, J. Fate of nitrogen-15 as influenced by soil and nutrient management history in a 19-year wheat–maize experiment. Field Crops Res. 2013, 144, 126–134. [Google Scholar] [CrossRef]
- Pant, A.P.; Radovich, T.J.; Hue, N.V.; Talcott, S.T.; Krenek, K.A. Vermicompost extracts influence growth, mineral nutrients, phytonutrients and antioxidant activity in pak choi (Brassica rapa cv. Bonsai, Chinensis group) grown under vermicompost and chemical fertilizer. J. Sci. Food Agric. 2009, 89, 2383–2392. [Google Scholar] [CrossRef]
- Grobelak, A.; Napora, A.; Kacprzak, M. Using plant growth-promoting rhizobacteria (PGPR) to improve plant growth. Ecol. Eng. 2015, 84, 22–28. [Google Scholar] [CrossRef]
- Wang, L.; Kaur, M.; Zhang, P.; Li, J.; Xu, M. Effect of Different Agricultural Farming Practices on Microbial Biomass and Enzyme Activities of Celery Growing Field Soil. Int. J. Environ. Res. Public Health 2021, 18, 12862. [Google Scholar] [CrossRef]
- Bhunia, S.; Bhowmik, A.; Mallick, R.; Mukherjee, J. Agronomic Efficiency of Animal-Derived Organic Fertilizers and Their Effects on Biology and Fertility of Soil: A Review. Agronomy 2021, 11, 823. [Google Scholar] [CrossRef]
- Treonis, A.M.; Austin, E.E.; Buyer, J.S.; Maul, J.E.; Spicer, L.; Zasada, I.A. Effects of organic amendment and tillage on soil microorganisms and microfauna. Appl. Soil Ecol. 2010, 46, 103–110. [Google Scholar] [CrossRef]
- Wang, J.; Song, Y.; Ma, T.; Raza, W.; Li, J.; Howland, J.G.; Huang, Q.; Shen, Q. Impacts of inorganic and organic fertilization treatments on bacterial and fungal communities in a paddy soil. Appl. Soil Ecol. 2017, 112, 42–50. [Google Scholar] [CrossRef]
- Luo, Y.; van Veelen, H.P.J.; Chen, S.; Sechi, V.; ter Heijne, A.; Veeken, A.; Buisman, C.J.N.; Bezemer, T.M. Effects of sterilization and maturity of compost on soil bacterial and fungal communities and wheat growth. Geoderma 2022, 409, 115598. [Google Scholar] [CrossRef]
- Chen, Y.P.; Tsai, C.F.; Rekha, P.D.; Ghate, S.D.; Huang, H.Y.; Hsu, Y.H.; Liaw, L.L.; Young, C.C. Agricultural management practices influence the soil enzyme activity and bacterial community structure in tea plantations. Bot. Stud. 2021, 62, 8. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Lewu, F.B.; Mulidzi, R.; Ncube, B. The biological activities of β-glucosidase, phosphatase and urease as soil quality indicators: A review. J. Soil Sci. Plant Nutr. 2017, 17, 794–807. [Google Scholar] [CrossRef] [Green Version]
- Cordovil, C.M.D.S.; de Varennes, A.; Pinto, R.M.D.S.; Alves, T.F.; Mendes, P.; Sampaio, S.C. Decomposition rate and enzymatic activity of composted municipal waste and poultry manure in the soil in a biofuel crops field. J. Sci. Food Agric. 2017, 97, 2245–2255. [Google Scholar] [CrossRef]
- Ruiz, J.L.; Salas, M.C. Evaluation of organic substrates and microorganisms as bio-fertilization tool in container crop production. Agronomy 2019, 9, 705. [Google Scholar] [CrossRef] [Green Version]
- Vargas-García, M.C.; Suárez-Estrella, F.; López, M.J.; Moreno, J. Microbial population dynamics and enzyme activities in composting processes with different starting materials. Waste Manag. 2010, 30, 771–778. [Google Scholar] [CrossRef]
- Bergougnoux, V. The history of tomato: From domestication to biopharming. Biotechnol. Adv. 2014, 32, 70–189. [Google Scholar] [CrossRef]
- Faostat. Available online: http://www.fao.org/faostat/es (accessed on 13 December 2021).
- Cadahia, C. La savia como índice de fertilización. Cultivos agroenergéticos, hortícolas, frutales y ornamentales. Mundi-Prensa Madr. 2008, 35–68. [Google Scholar]
- Wilson, P.W.; Knight, S.C. Experiments in Bacterial Physiology; Burgess: Minneapolis, MN, USA, 1952. [Google Scholar]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 4, 301–307. [Google Scholar] [CrossRef]
- Sánchez, N.A.; Sánchez, J.A.; Salas, M.C.; Arantzazu, M.B.; Delgado, M.J. Medium-term influence of organic fertilization on the quality and yield of a celery crop. Agronomy 2020, 10, 1418. [Google Scholar]
- Bilalis, D.; Krokida, M.; Roussis, I.; Papastylianou, P.; Travlos, I.; Cheimona, N.; Dede, A. Effects of organic and inorganic fertilization on yield and quality of processing tomato (Lycopersicon esculentum Mill.). Folia Hort. 2018, 30, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Riahi, A.; Hdider, C.; Sanaa, M.; Tarchoun, N.; Kheder, M.B.; Guezal, I. Effect of conventional and organic productions systems on the yield and quality of field tomato cultivars grown in Tunisia. J. Sci. Food Agric. 2009, 89, 2275–2282. [Google Scholar] [CrossRef]
- Arancon, N.Q.; Edwards, C.A.; Bierman, P.; Welch, C.; Metzger, J.D. Influences of vermicomposts on field strawberries: Effects on growth and yields. Bioresour. Technol. 2004, 93, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Zhao, F.; Zhang, G.; Zhang, Y.; Yang, L. Vermicompost improves tomato yield and quality and the biochemical properties of soils with different tomato planting history in a greenhouse study. Front. Plant Sci. 2017, 8, 1978. [Google Scholar] [CrossRef] [Green Version]
- Murmu, K.; Ghosh, B.C.; Swain, D.K. Yield and quality of tomato grown under organic and conventional nutrient management. Arch. Agron. Soil Sci. 2013, 59, 1311–1321. [Google Scholar] [CrossRef]
- Hale, L.; Curtis, D.; Azeem, M.; Montgomery, J.; Crowley, D.E.; McGiffen, M.E., Jr. Influence of compost and biochar on soil biological properties under turfgrass supplied deficit irrigation. Appl. Soil Ecol. 2021, 168, 104134. [Google Scholar] [CrossRef]
- Fu, H.; Zhang, G.; Zhang, F.; Sun, Z.; Geng, G.; Li, T. Effects of continuous tomato monoculture on soil microbial properties and enzyme activities in a solar greenhouse. Sustainability 2017, 9, 317. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.G.; Wu, F.Z. Changes in soil chemical characters and enzyme activities during continuous monocropping of cucumber (Cucumis sativus). Pak. J. Bot. 2015, 47, 691–697. [Google Scholar]
- Guerra, P.A.M.; Sanjúan, M.C.S.; López, M.J. Evaluation of physicochemical properties and enzymatic activity of organic substrates during four crop cycles in soilless containers. Food Sci. Nutr. 2018, 6, 2066–2078. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Production Cycle | Transplant | Last Harvest |
|---|---|---|
| Tomato cycle 0 (TC0) * | 2 February 2017 | 6 July 2017 |
| Tomato cycle 1 (TC1) | 22 August 2017 | 30 January 2018 |
| Tomato cycle 2 (TC2) | 6 February 2018 | 10 July 2018 |
| Tomato cycle 3 (TC3) | 22 August 2018 | 25 February 2019 |
| Treatments | TC1 | TC2 | TC3 | ||
|---|---|---|---|---|---|
| FertControl | 4.86 | 7.51 | a | 5.44 | a |
| OrgControl | 5.38 | 6.35 | b | 4.53 | c |
| CropRes | 5.32 | 7.23 | a | 4.74 | bc |
| Comp | 5.21 | 7.46 | a | 5.24 | ab |
| VermiComp | 5.31 | 6.88 | ab | 5.28 | ab |
| ns | * | * | |||
| Cycle | DAT | FertControl | OrgControl | CropRes | Comp | Vermicomp |
|---|---|---|---|---|---|---|
| N-NO3− | ||||||
| TC2 | 111 | 1729 b | 1327 c | 1189 c | 1288 c | 1994 a |
| 126 | 1428 a | 1374 b | 874 c | 1205 bc | 1016 bc | |
| 141 | 1705 a | 1352 a | 1808 a | 1831 a | 1339 a | |
| Mean | 1621 a | 1351 a | 1290 a | 1441 a | 1450 a | |
| TC3 | 101 | 1696 a | 1033 b | 1321 ab | 1721 a | 1362 ab |
| 118 | 1918 b | 2286 ab | 2359 b | 2858 a | 1965 b | |
| 151 | 2093 a | 1938 a | 1438 b | 2053 a | 2057 a | |
| Mean | 1902 a | 1752 a | 1706 a | 2211 a | 1795 a | |
| K+ | ||||||
| TC2 | 111 | 3925 b | 3732 b | 3926 b | 3918 b | 4959 a |
| 126 | 3147 a | 2935 a | 3188 a | 3031 a | 3031 a | |
| 141 | 4117 a | 2586 c | 3663 b | 3395 b | 4066 a | |
| Mean | 3730 ab | 3048 b | 3592 ab | 3448 ab | 4019 a | |
| TC3 | 101 | 3843 c | 5185 a | 4439 bc | 5160 a | 4555 b |
| 118 | 4279 c | 4814 b | 4999 b | 5426 a | 4438 c | |
| 151 | 3425 c | 3592 ab | 3951 a | 3626 ab | 3504 b | |
| Mean | 3849 b | 4530 a | 4451 a | 4737 a | 4166 ab | |
| BT | FT | AB | ||||
|---|---|---|---|---|---|---|
| FertControl | 5.68 | b | 3.90 | a | 3.00 | b |
| OrgControl | 5.66 | b | 2.45 | b | 7.31 | a |
| CropRes | 7.48 | ab | 2.82 | b | 7.37 | a |
| Comp | 7.60 | a | 2.52 | b | 7.48 | a |
| VermiComp | 6.36 | ab | 2.70 | b | 7.43 | a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carricondo-Martínez, I.; Falcone, D.; Berti, F.; Orsini, F.; Salas-Sanjuan, M.d.C. Use of Agro-Waste as a Source of Crop Nutrients in Intensive Horticulture System. Agronomy 2022, 12, 447. https://doi.org/10.3390/agronomy12020447
Carricondo-Martínez I, Falcone D, Berti F, Orsini F, Salas-Sanjuan MdC. Use of Agro-Waste as a Source of Crop Nutrients in Intensive Horticulture System. Agronomy. 2022; 12(2):447. https://doi.org/10.3390/agronomy12020447
Chicago/Turabian StyleCarricondo-Martínez, Isidoro, Domenico Falcone, Francesca Berti, Francesco Orsini, and María del Carmen Salas-Sanjuan. 2022. "Use of Agro-Waste as a Source of Crop Nutrients in Intensive Horticulture System" Agronomy 12, no. 2: 447. https://doi.org/10.3390/agronomy12020447
APA StyleCarricondo-Martínez, I., Falcone, D., Berti, F., Orsini, F., & Salas-Sanjuan, M. d. C. (2022). Use of Agro-Waste as a Source of Crop Nutrients in Intensive Horticulture System. Agronomy, 12(2), 447. https://doi.org/10.3390/agronomy12020447