
Relatively Low Light Intensity Promotes Phosphorus Absorption and Enhances the Ethylene Signaling Component EIN3 in Maize, Wheat, and Oilseed Rape


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
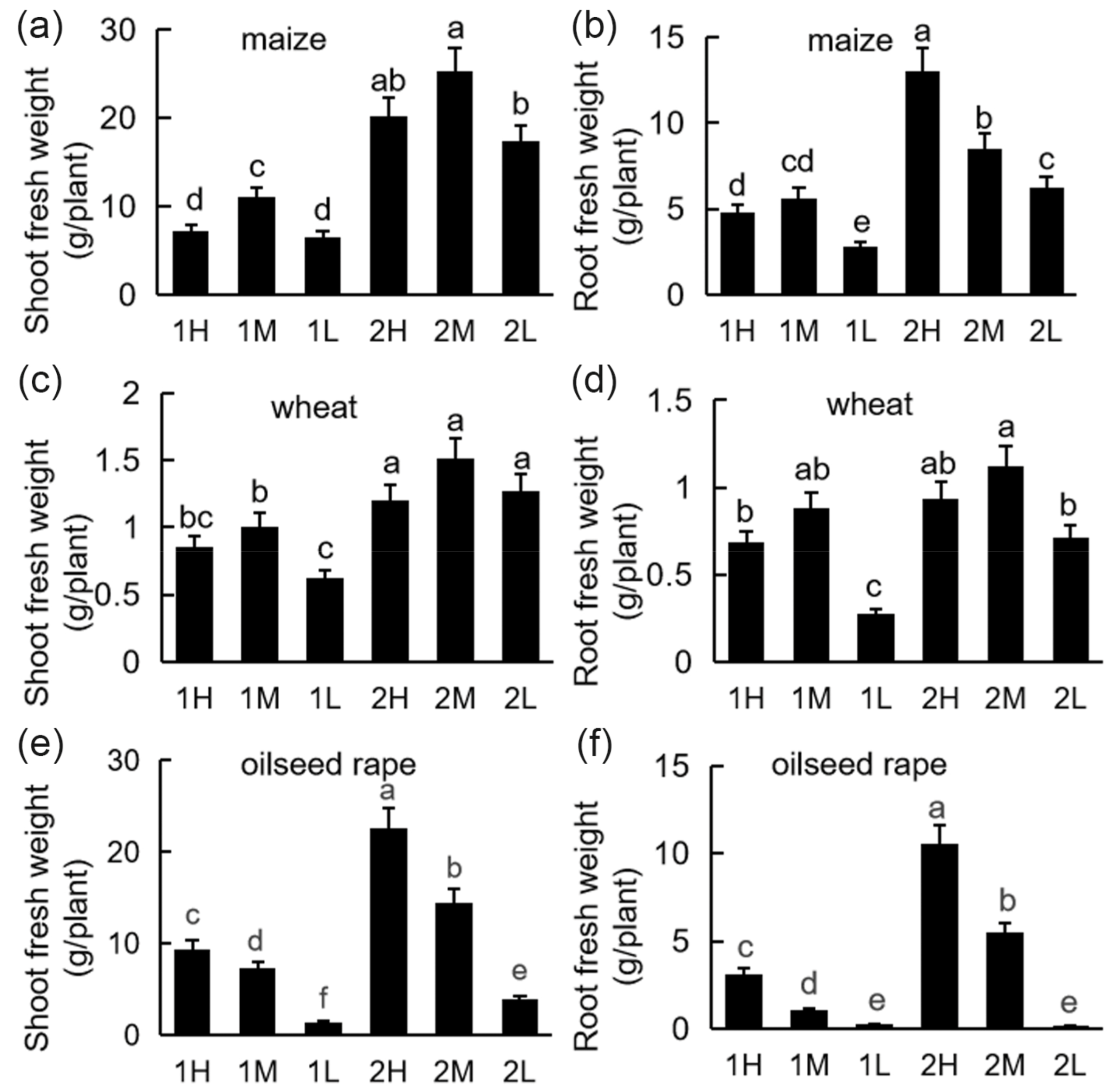
2.2. Plant Growth and Biomass
2.3. Determination of Photosystem Parameters
2.4. Determination of Starch and Sucrose Contents
2.5. Determination of Phosphorus Content
2.6. Acid Phosphatase Staining and Determination of Activity
2.7. Western Blotting of the EIN3 Protein
2.8. Determination of Ethylene Levels
2.9. Data Analysis
3. Results
3.1. Accumulation of Plant Biomass under Different Light Intensities
3.2. Differences in Photosynthetic Parameters under Different Light Intensities
3.3. Accumulation of Sucrose and Starch in Plants under Different Light Intensities
3.4. Phosphorus Contents of the Plants under Different Light Intensities
3.5. Secretion of APase from Plant Seedlings
3.6. EIN3 Protein Levels and Ethylene Levels in Plants under Different Light Intensities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: A review. Plant Soil. 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.M.; Mcneill, A.M.; Prigent, C.C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil. 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Ye, D.; Zhang, X.; Li, T.; Xu, J.; Chen, G. Phosphorus-acquisition characteristics and rhizosphere properties of wild barley in relation to genotypic differences as dependent on soil phosphorus availability. Plant Soil. 2018, 423, 503–516. [Google Scholar] [CrossRef]
- Holford, I.C.R.; Hird, C.; Lawrie, R. Effects of animal effluents on the phosphorus sorption characteristics of soils. Soil Res. 1997, 35, 365–373. [Google Scholar] [CrossRef]
- Fang, Z.Y.; Shao, C.; Meng, Y.J.; Ping, W.; Chen, M. Phosphate signaling in Arabidopsis and Oryza sativa. Plant Sci. 2009, 176, 170–180. [Google Scholar] [CrossRef]
- Hammond, J.P.; Broadley, M.R.; White, P.J. Genetic responses to phosphorus deficiency. Ann. Bot. 2004, 94, 323–332. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde, S.C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2010, 157, 423–447. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Wu, P.; Ling, H.; Xu, G.; Xu, F.; Zhang, Q. Plant nutriomics in China: An overview. Ann. Bot. 2006, 98, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Cordell, D.; Drangert, J.O.; White, S. The story of phosphorus: Global food security and food for thought. Global Environ. Change 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Zhu, J.; Kaeppler, S.M.; Lynch, J.P. Mapping of QTL controlling roots hair length in maize (Zea mays L.) under phosphorus deficiency. Plant Soil. 2005, 270, 299–310. [Google Scholar] [CrossRef]
- Nadeem, M.; Mollier, A.; Morel, C.; Shahid, M. Maize seedling phosphorus nutrition: Allocation of remobilized seed phosphorus reserves and external phosphorus uptake to seedling roots and shoots during early growth stages. Plant Soil. 2013, 371, 327–338. [Google Scholar] [CrossRef]
- Szabó-Nagy, A.; Oláh, Z.; Erdei, L. Phosphatase induction in roots of winter wheat during adaptation to phosphorus deficiency. Physiol. Plant. 2010, 70, 544–552. [Google Scholar] [CrossRef]
- Su, J.; Xiao, Y.; Ming, L.; Liu, Q.; Li, B.; Tong, Y.; Jia, J.; Li, Z. Mapping QTLs for phosphorus- deficiency tolerance at wheat seedling stage. Plant Soil. 2012, 281, 25–36. [Google Scholar] [CrossRef]
- Eckstein, A.; Ziba, P.; Gabry, H. Sugar and light effects on the condition of the photosynthetic apparatus of Arabidopsis thaliana cultured in vitro. J. Plant Growth Regul. 2012, 31, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Tang, X.; Vance, C.P.; White, P.J.; Zhang, F.; Shen, J. Interactions between light and phosphorus nutrition affect the phosphate-mining capacity of white lupin (Lupinus albus L.). J. Exp. Bot. 2014, 65, 2995–3003. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Li, H.; Shen, J.; Rengel, Z. Maize responds to low shoots P concentration by altering roots morphology rather than increasing roots exudation. Plant Soil. 2017, 416, 377–389. [Google Scholar] [CrossRef]
- Wang, L.; Dong, J.; Gao, Z.; Liu, D. The Arabidopsis gene hypersensitive to phosphate starvation 3 encodes Ethylene Overproduction 1. Plant Cell Physiol. 2012, 6, 1093–1105. [Google Scholar] [CrossRef] [Green Version]
- Nagarajan, V.; Smith, L.P. Ethylene’s role in phosphate starvation signaling: More than just a roots growth regulator. Plant Cell Physiol. 2012, 53, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Li, Z.; Qian, W.; Guo, W.; Gao, X.; Huang, L.; Wang, H.; Zhu, H.; Wu, J.W.; Wang, D. The Arabidopsis purple acid phosphatase AtPAP10 is predominantly associated with the root surface and plays an important role in plant tolerance to phosphate limitation. Plant Physiol. 2011, 157, 1283–1299. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xie, Y.; Wang, H.; Ma, X.; Yao, W.; Wang, H. Light and ethylene coordinately regulate the phosphate starvation response through transcriptional regulation of phosphate starvation response1. Plant Cell 2017, 29, 2269–2284. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Wang, J.; Zhang, Z.; Quan, R.; Zhang, H.; Deng, X.W.; Ma, L.; Huang, R. Ethylene promotes hypocotyl growth and HY5 degradation by enhancing the movement of COP1 to the nucleus in the light. PLoS Genet. 2013, 9, e1004025. [Google Scholar] [CrossRef]
- Stepanova, A.N.; Alonso, J.M. Cutting out the middle man in light-hormone interactions. Dev. Cell 2016, 39, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.W.; Shi, H.; Xue, C.; Ning, W.; Xing, W.D. Ethylene-orchestrated circuitry coordinates a seedling’s response to soil cover and etiolated growth. Proc. Natl. Acad. Sci. USA 2014, 111, 3913–3920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Shen, X.; Liu, R.; Xue, C.; Wei, N.; Deng, X.W.; Zhong, S.W. The red light receptor phytochrome B directly enhances substrate-e3 ligase interactions to attenuate ethylene responses. Dev. Cell 2016, 39, 597–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Ecker, J.R. Plant responses to ethylene gas are mediated by SCF (EBF1/EBF2)—Dependent proteolysis of EIN3 transcription factor. Cell 2003, 115, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Potuschak, T.; Lechner, E.; Parmentier, Y.; Yanagisawa, S.; Grava, S.; Koncz, C.; Genschik, P. EIN3-dependent regulation of plant ethylene hormone signaling by two arabidopsis F box proteins: EBF1 and EBF2. Cell 2003, 115, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Solano, R.; Stepanova, A.; Chao, Q.M.; Ecker, J.R. Nuclear events in ethylene signaling: A transcriptional cascade mediated by ETHYLENE- INSENSITIVE3 and ETHYLENE- RESPONSEFACTOR1. Genes Dev. 1998, 12, 3703–3714. [Google Scholar] [CrossRef] [Green Version]
- Ames, B.N. Assay of inorganic phosphate, total phosphate and phosphatase. Method Enzymol. 1966, 8, 114–118. [Google Scholar]
- Gibson, D.M.; Christen, A.A.; Mullaney, E.J. Direct screening for acid phosphatase production on bcip-agar plates. Biotechnol. Tech. 1988, 2, 63–68. [Google Scholar] [CrossRef]
- Yuan, W.; Zhou, J.; Tong, J.; Zhuo, W.; Qian, W. ALBA protein complex reads genic R-loops to maintain genome stability in Arabidopsis. Sci. Adv. 2019, 55, eaav9040. [Google Scholar] [CrossRef] [Green Version]
- Vogel, J.P.; Woeste, K.E.; Theologis, A.; Kieber, J.J. Recessive and dominant mutations in the ethylene biosynthetic gene ACS5 of Arabidopsis confer cytokinin insensitivity and ethylene overproduction, respectively. Proc. Natl. Acad. Sci. USA 1998, 95, 4766–4771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, R.; Kumar, D.; Sultana, A.; Hazra, S.; Bhattacharyya, D.; Chattopadhyay, S. Glutathione regulates 1-aminocyclopropane-1-carboxylate synthase transcription via WRKY33 and 1-aminocyclopropane-1-carboxylate oxidase by modulating messenger RNA stability to induce ethylene synthesis during stress. Plant Physiol. 2015, 169, 2963–2981. [Google Scholar]
- Lei, M.; Zhu, C.; Liu, Y.; Karthikeyan, A.S.; Bressan, R.A.; Raghothama, K.G.; Liu, D. Ethylene signalling is involved in regulation of phosphate starvation-induced gene expression and production of acid phosphatases and anthocyanin in Arabidopsis. New Phytol. 2011, 189, 1084–1095. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.G.; Liu, Y.D.; Zhang, B.C.; Zhao, Y.T.; Wang, X.J.; Zhou, Y.H.; Raghothama, K.G. Genetic and genomic evidence that sucrose is a global regulator of plant responses to phosphate starvation in Arabidopsis. Plant Physiol. 2011, 156, 1116–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.W.; Feng, L.Y.; Wang, J.H.; Fu, Y.F.; Yuan, S. Two-factor ANOVA of SSH and RNA-seq analysis reveal development-associated Pi-starvation genes in oilseed rape. Planta 2019, 4, 1073–1088. [Google Scholar] [CrossRef] [PubMed]
- Groots, C.; Marcelis, L.; Boogaard, R.; Lambers, H. Growth and dry-mass partitioning in tomato as affected by phosphorus nutrition and light. Plant Cell Environ. 2010, 24, 1309–1317. [Google Scholar] [CrossRef]
- Gent, M.P.N. Carbohydrate level and growth of tomato plants: The effect of irradiance and temperature. Plant Physiol. 1986, 81, 1075–1079. [Google Scholar] [CrossRef] [Green Version]
- Fredeen, A.L.; Rao, I.M.; Terry, N. Influence of phosphorus nutrition on growth and carbon partitioning in Glycine max. Plant Physiol. 1989, 89, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Sims, D.A.; Seemann, J.R.; Luo, Y. The significance of differences in the mechanisms of photosynthetic acclimation to light, nitrogen and CO2 for return on investment in leaves. Funct. Ecol. 1998, 12, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Wang, L.; Li, S.; Gao, Y.; Yong, L.; Zhao, D. Interactions between light and phosphorus nutrition affect the p uptake capacity of maize and soybean seedling in a low light area. Front. Plant Sci. 2019, 10, 183–198. [Google Scholar] [CrossRef]
- Kang, J.; Yu, H.; Tian, C.; Zhou, W.; Li, C.; Jiao, Y.; Liu, D. Suppression of photosynthetic gene expression in roots is required for sustained root growth under phosphate deficiency. Plant Physiol. 2014, 165, 1156–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, F.; Zhao, Q.; Ji, Y.; Li, W.; Jiang, Z.; Yu, X.; Zhang, C.; Han, Y.; He, W.; Liu, Y. Ethylene-induced stabilization of ETHYLENE INSENSITIVE3 and EIN3-LIKE1 is mediated by proteasomal degradation of EIN3 binding F-box 1 and 2 that requires EIN2 in Arabidopsis. Plant Cell 2010, 22, 2384–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, C.; Chang, C. Advances in ethylene signalling: Protein complexes at the endoplasmic reticulum membrane. AoB Plants 2012, 2012, pls031. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ma, M.; Feng, Y.; Li, H.; Wang, Y.; Ma, Y.; Li, M.; An, F.; Guo, H. EIN2-directed translational regulation of ethylene signaling in Arabidopsis. Cell 2015, 163, 670–683. [Google Scholar] [CrossRef] [Green Version]
- Merchante, C.; Brumos, J.; Yun, J.; Hu, Q.; Spencer, K.R.; Enriquez, P.; Binder, B.M.; Heber, S.; Stepanova, A.N.; Alonso, J.M. Gene-specifific translation regulation mediated by the hormone- signaling molecule EIN2. Cell 2015, 163, 684–697. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Shen, Z.; Huang, S.S.; Schmitz, R.J.; Urich, M.A.; Briggs, S.P.; Ecker, J.R. Processing and subcellular traffificking of ER-tethered EIN2 control response to ethylene gas. Science 2012, 338, 390–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, X.; Zhang, C.; Ji, Y.; Zhao, Q.; He, W.; An, F.; Jiang, L.; Guo, H. Activation of ethylene signaling is mediated by nuclear translocation of the cleaved EIN2 carboxyl terminus. Cell Res. 2012, 22, 1613–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, S.; Zhao, M.; Shi, T.; Shi, H.; An, F.Y.; Zhao, Q.; Guo, H.W. EIN3/EIL1 cooperate with PIF1 to prevent photo-oxidation and to promote greening of Arabidopsis seedlings. Proc. Natl. Acad. Sci. USA 2009, 106, 21431–21436. [Google Scholar] [CrossRef] [Green Version]
- Lau, O.S.; Deng, X.W. The photomorphogenic repressors COP1 and DET1: 20 years later. Trends Plant Sci. 2012, 17, 584–593. [Google Scholar] [CrossRef]
- Von Arnim, A.; Deng, X.W. Light control of seedling development. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 215–243. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.A.; Bianchetti, R.E.; Freschi, L. Shedding light on ethylene metabolism in higher plants. Front. Plant Sci. 2014, 5, 665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grodzinski, B.; Boesel, I.; Horton, R.F. Light stimulation of ethylene release from leaves of Gomphrena globosa L. Plant Physiol. 1983, 71, 588–593. [Google Scholar] [CrossRef] [Green Version]
- Vandenbussche, F.; Vriezen, W.H.; Smalle, J.; Laarhoven, L.J.; Harren, F.J.; Van Der Straeten, D. Ethylene and auxin control the Arabidopsis response to decreased light intensity. Plant Physiol. 2003, 133, 517–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenabeele, S.; Van Der Kelen, K.; Dat, J.; Gadjev, I.; Boonefaes, T.; Morsa, S.; Rottiers, P.; Slooten, L.; Van Montagu, M.; Zabeau, M.; et al. A comprehensive analysis of hydrogen peroxide-induced gene expression in tobacco. Proc. Natl. Acad. Sci. USA 2003, 100, 16113–16118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, T.; Sato, F. The function of ETHYLENE RESPONSE FACTOR genes in the light-induced anthocyanin production of Arabidopsis thaliana leaves. Plant Biotechnol. 2018, 35, 87–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Kumar, S.; Mohapatra, T. Interaction between macro- and micro-nutrients in plants. Front. Plant Sci. 2021, 12, 665583. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, R. Nitrogen and phosphorus signaling and transport during Legume-Rhizobium symbiosis. Front. Plant Sci. 2021, 12, 683601. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Net Photosynthetic Rate (μmol m−2 s−1) | Stomatal Conductance (mol m−2 s−1) | Intercellular CO2 Concentration (μmol mol−1) | Transpiration Rate (mmol m−2 s−1) | ||
|---|---|---|---|---|---|
| Maize | 1H | 7.96 ± 0.04 bc | 0.15 ± 0.01 a | 316.06 ± 6.16 b | 2.12 ± 0.01 a |
| 1M | 6.56 ± 0.10 d | 0.11 ± 0.01 ab | 312.69 ± 1.88 b | 1.46 ± 0.02 b | |
| 1L | 5.56 ± 0.19 e | 0.12 ± 0.05 a | 303.86 ± 2.93 b | 1.55 ± 0.60 b | |
| 2H | 9.59 ± 0.09 a | 0.06 ± 0.01 b | 154.41 ± 2.91 d | 0.82 ± 0.01 c | |
| 2M | 7.68 ± 0.05 c | 0.07 ± 0.01 b | 218.91 ± 1.73 c | 0.76 ± 0.01 c | |
| 2L | 8.05 ± 0.02 bc | 0.07 ± 0.01 b | 212.03 ± 0.12 c | 0.78 ± 0.01 c | |
| Net Photosynthetic Rate (μmol m−2 s−1) | Stomatal Conductance (mol m−2 s−1) | Intercellular CO2 Concentration (μmol mol−1) | Transpiration Rate (mmol m−2 s−1) | ||
|---|---|---|---|---|---|
| wheat | 1H | 3.52 ± 0.06 e | 0.05 ± 0.01 d | 157.06 ± 1.95 f | 1.25 ± 0.01 d |
| 1M | 6.85 ± 0.00 d | 0.14 ± 0.01 b | 188.95 ± 0.08 e | 3.17 ± 0.01 b | |
| 1L | 6.74 ± 0.07 d | 0.14 ± 0.01 b | 188.28 ± 0.23 e | 3.11 ± 0.02 c | |
| 2H | 3.37 ± 0.09 e | 0.05 ± 0.01 d | 281.69 ± 1.04 a | 0.48 ± 0.01 g | |
| 2M | 8.54 ± 0.02 a | 0.09 ± 0.01 c | 259.44 ± 0.32 b | 1.11 ± 0.01 e | |
| 2L | 7.79 ± 0.08 b | 0.09 ± 0.01 c | 250.16 ± 3.80 c | 0.87 ± 0.01 f | |
| Net Photosynthetic Rate (μmol m−2 s−1) | Stomatal Conductance (mol m−2 s−1) | Intercellular CO2 Concentration (μmol mol−1) | Transpiration Rate (mmol m−2 s−1) | ||
|---|---|---|---|---|---|
| oilseed rape | 1H | 6.38 ± 0.20 d | 0.11 ± 0.03 b | 313.53 ± 2.27 b | 1.47 ± 0.04 b |
| 1M | 8.76 ± 0.05 b | 0.10 ± 0.02 c | 264.54 ± 0.36 e | 1.15 ± 0.03 c | |
| 1L | 4.42 ± 0.08 f | 0.07 ± 0.02 e | 303.28 ± 1.37 c | 0.93 ± 0.02 e | |
| 2H | 4.86 ± 0.63 ef | 0.11 ± 0.03 a | 324.53 ± 0.29 a | 1.54 ± 0.07 a | |
| 2M | 9.72 ± 0.01 a | 0.11 ± 0.01 b | 246.49 ± 0.56 f | 1.05 ± 0.05 d | |
| 2L | 5.49 ± 0.17 e | 0.09 ± 0.01 d | 304.93 ± 2.59 c | 1.48 ± 0.06 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.-Y.; Cai, X.; Wan, Y.-T.; Fu, Y.-F.; Yang, X.-Y.; Zhang, Z.-W.; Yuan, S. Relatively Low Light Intensity Promotes Phosphorus Absorption and Enhances the Ethylene Signaling Component EIN3 in Maize, Wheat, and Oilseed Rape. Agronomy 2022, 12, 427. https://doi.org/10.3390/agronomy12020427
Zhang M-Y, Cai X, Wan Y-T, Fu Y-F, Yang X-Y, Zhang Z-W, Yuan S. Relatively Low Light Intensity Promotes Phosphorus Absorption and Enhances the Ethylene Signaling Component EIN3 in Maize, Wheat, and Oilseed Rape. Agronomy. 2022; 12(2):427. https://doi.org/10.3390/agronomy12020427
Chicago/Turabian StyleZhang, Meng-Yao, Xin Cai, Yu-Ting Wan, Yu-Fan Fu, Xin-Yue Yang, Zhong-Wei Zhang, and Shu Yuan. 2022. "Relatively Low Light Intensity Promotes Phosphorus Absorption and Enhances the Ethylene Signaling Component EIN3 in Maize, Wheat, and Oilseed Rape" Agronomy 12, no. 2: 427. https://doi.org/10.3390/agronomy12020427
APA StyleZhang, M.-Y., Cai, X., Wan, Y.-T., Fu, Y.-F., Yang, X.-Y., Zhang, Z.-W., & Yuan, S. (2022). Relatively Low Light Intensity Promotes Phosphorus Absorption and Enhances the Ethylene Signaling Component EIN3 in Maize, Wheat, and Oilseed Rape. Agronomy, 12(2), 427. https://doi.org/10.3390/agronomy12020427