
Physiological and Biochemical Responses to Water Stress and Salinity of the Invasive Moth Plant, Araujia sericifera Brot., during Seed Germination and Vegetative Growth


 ,
, 

 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Seed Germination
2.2. Plant Growth and Stress Treatments
2.3. Substrate Analysis
2.4. Photosynthetic Pigments
2.5. Ion Content Measurements
2.6. Malondialdehyde, Total Phenolics, and Flavonoids Quantification
2.7. Antioxidant Enzyme Activities
2.8. Statistical Analyses
3. Results
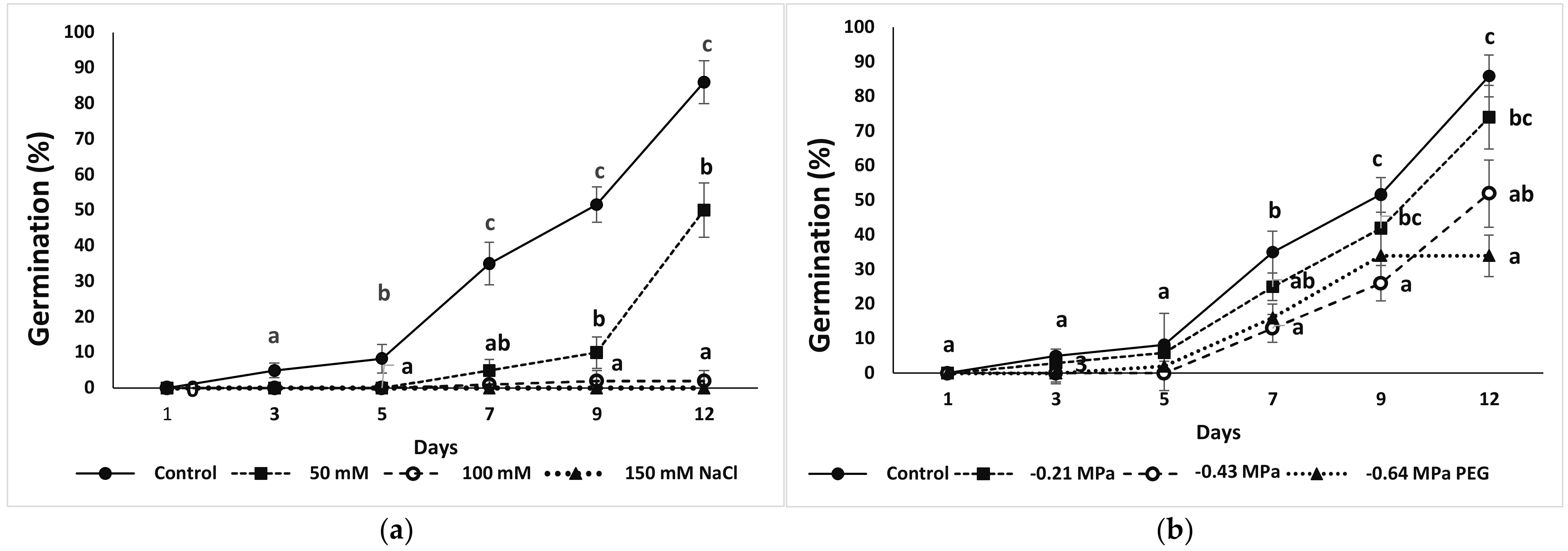
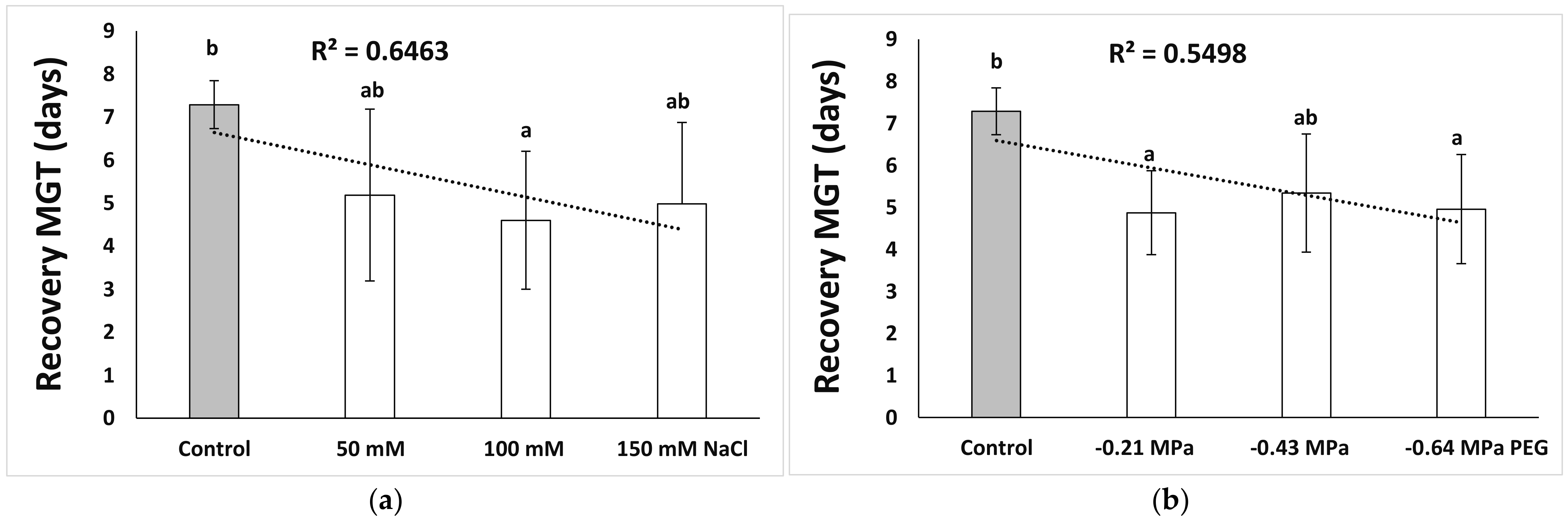
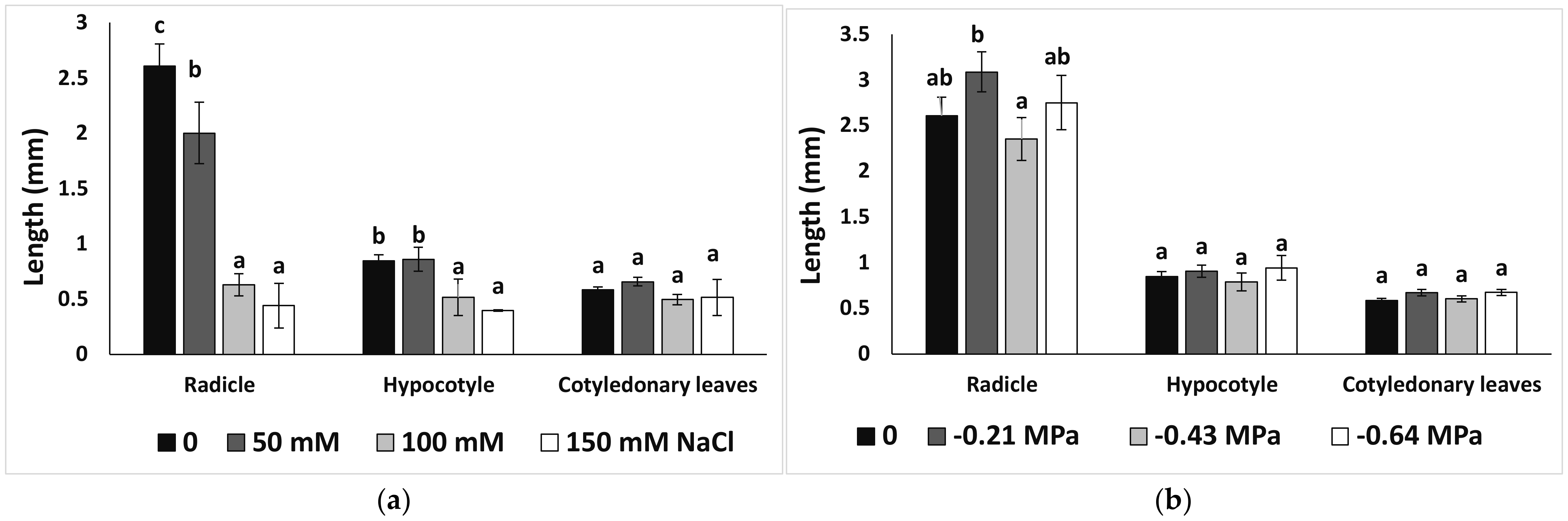
3.1. Seed Germination and Recovery of Germination
3.2. Substrate Analysis
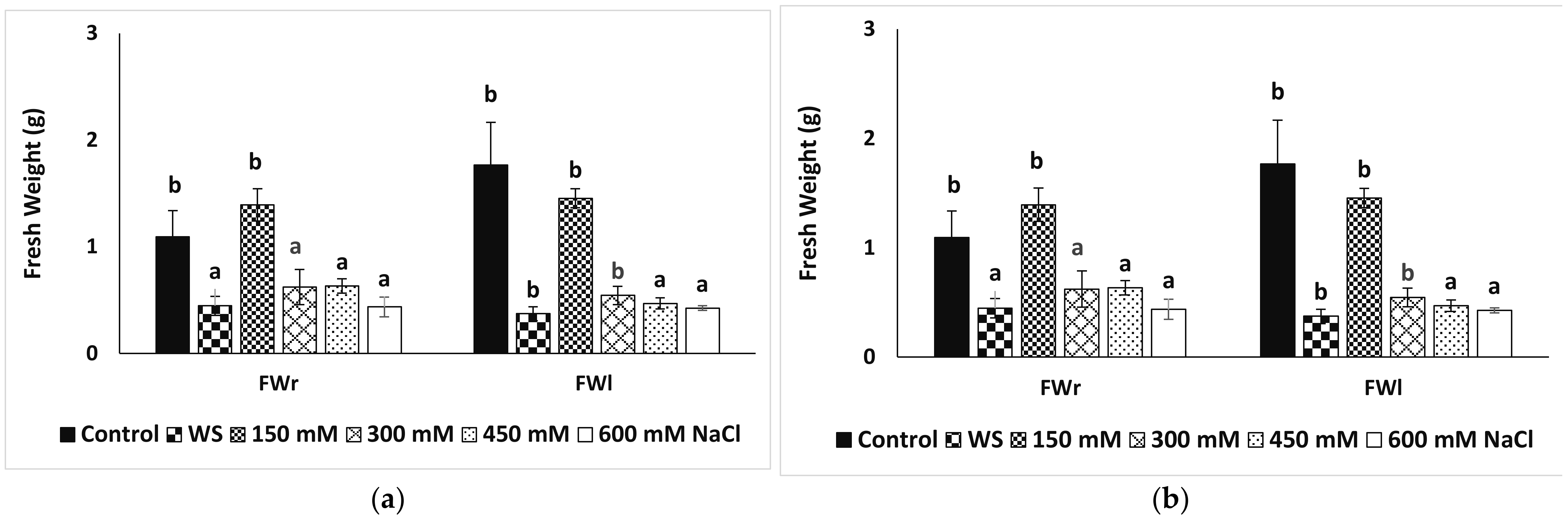
3.3. Growth Parameters and Photosyntetcic Pigments
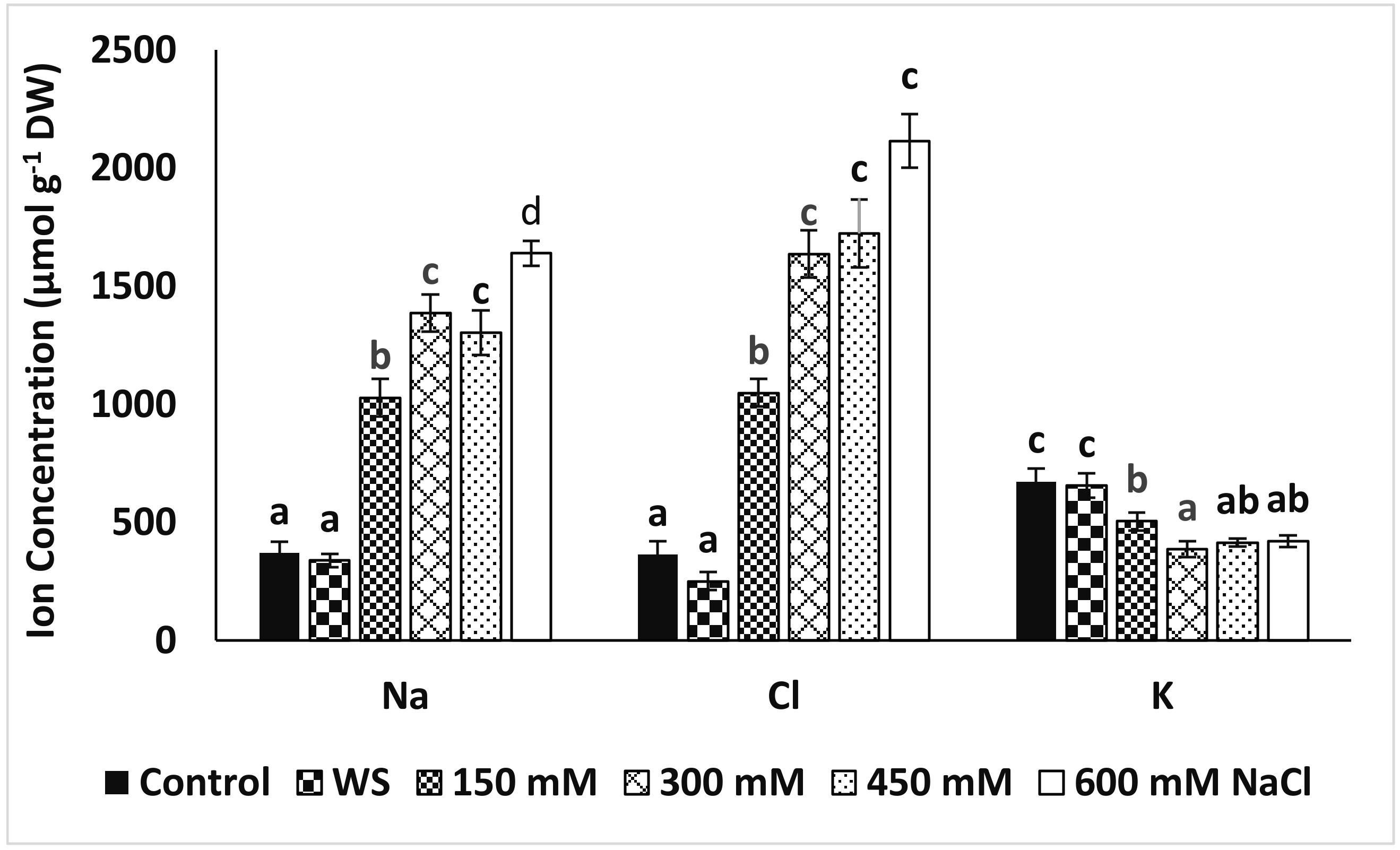
3.4. Ion Accumulation in Leaves
3.5. MDA, Non-Enzymatic Antioxidants, and Antioxidant Enzyme Activities
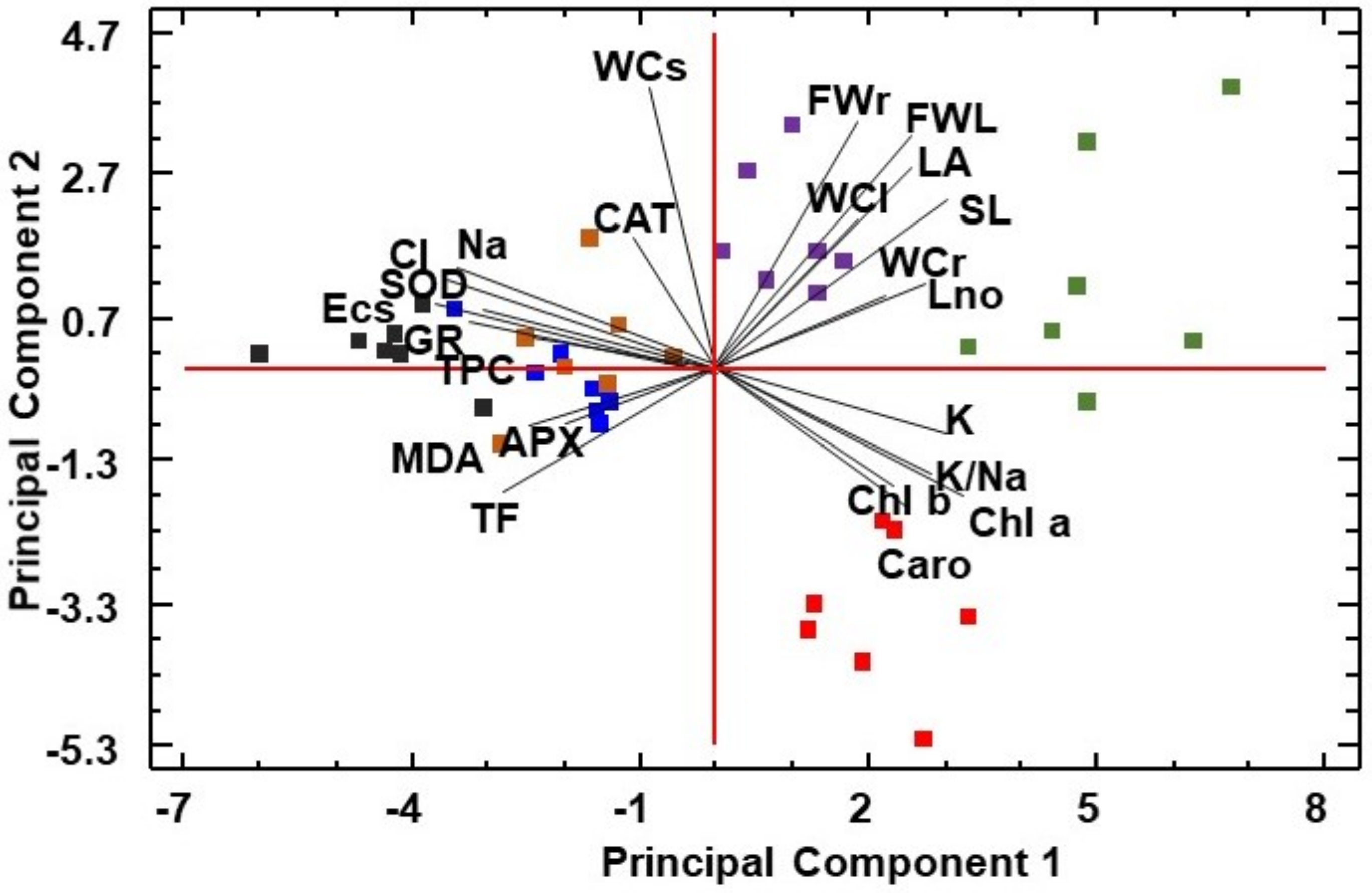
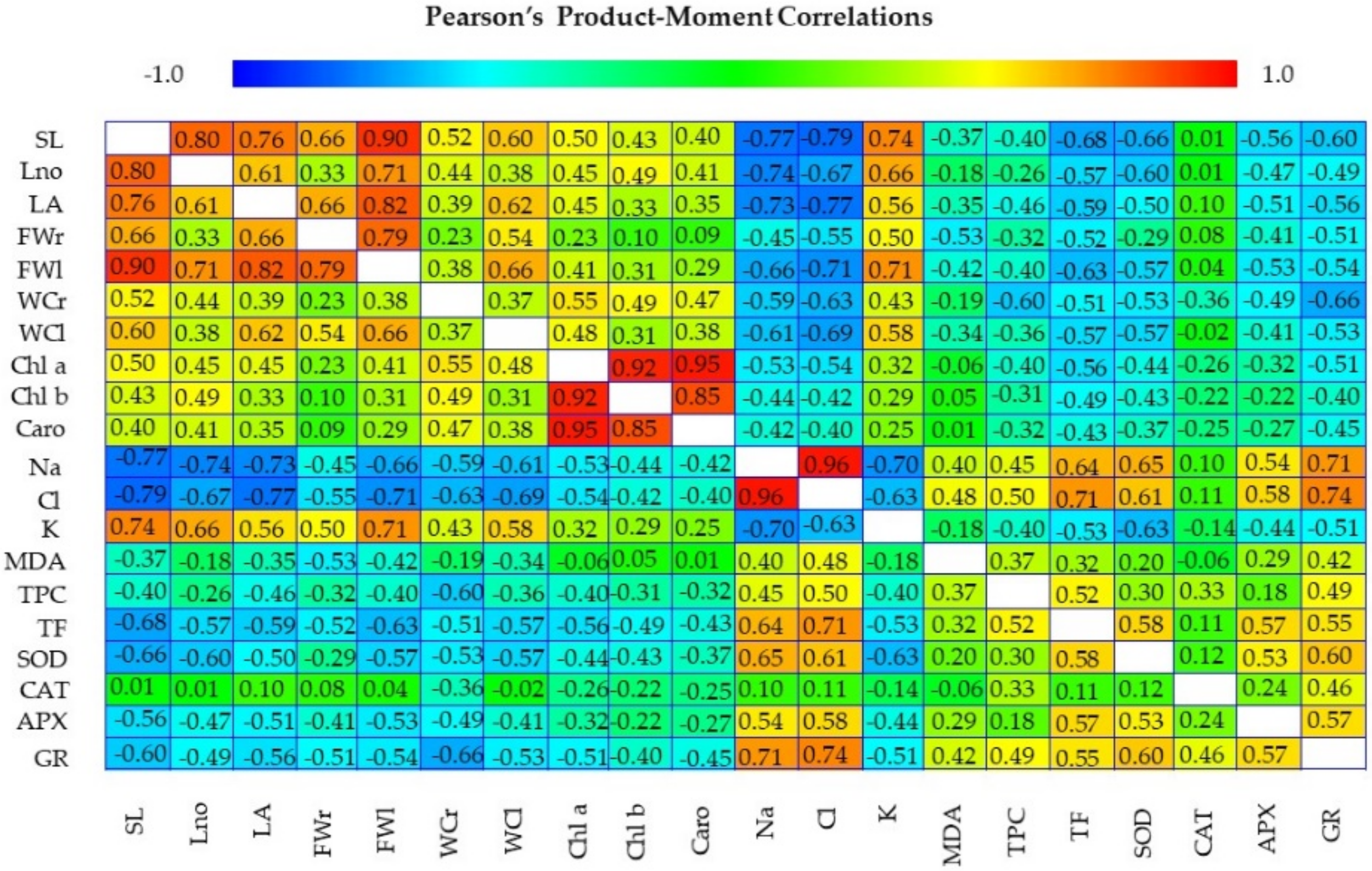
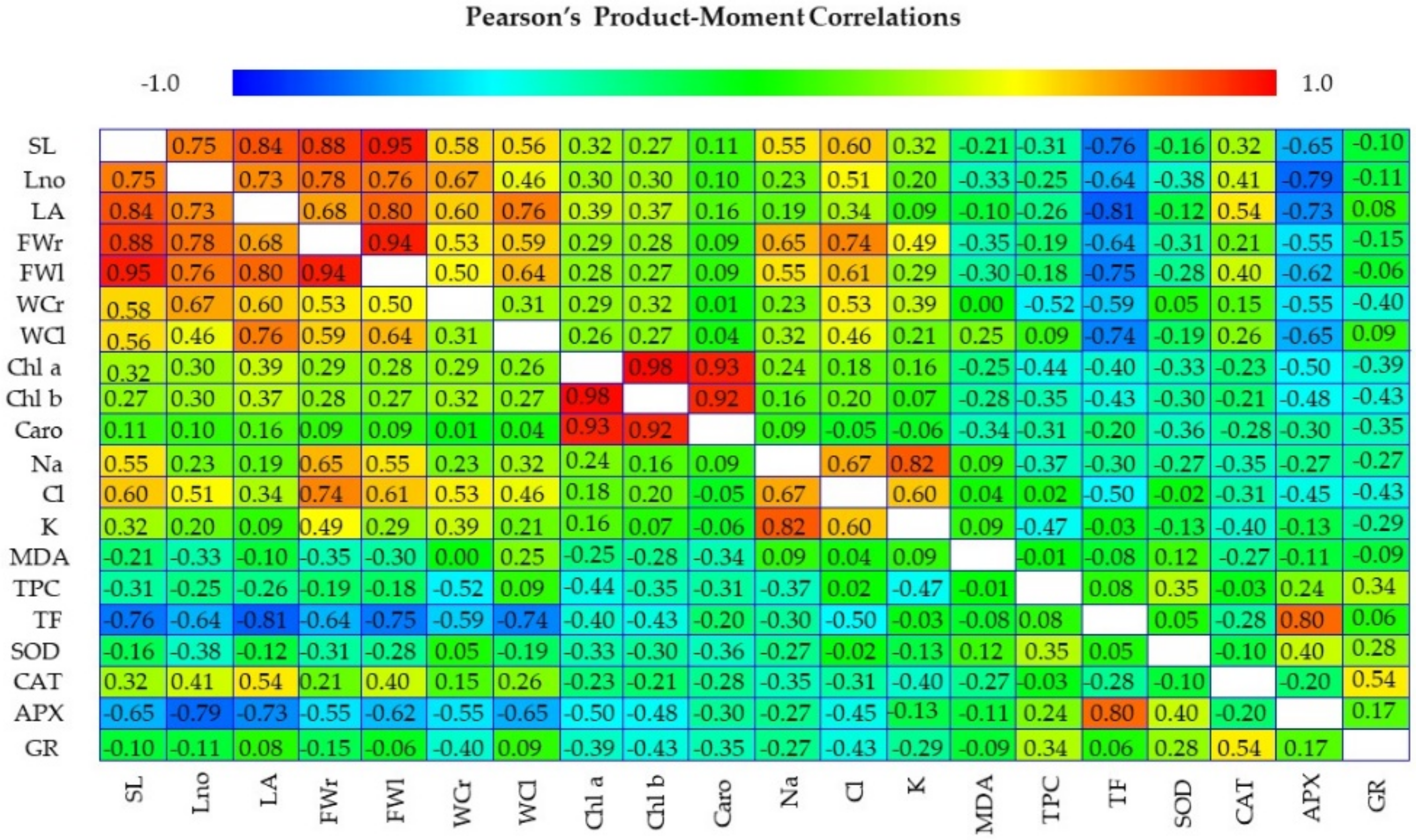
3.6. Multivariate Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bradley, B.A.; Wilcove, D.S.; Oppenheimer, M. Climate change increases risk of plant invasion in the Eastern United States. Biol. Invasions 2010, 12, 1855–1872. [Google Scholar] [CrossRef]
- Hulme, P.E. Climate change and biological invasions: Evidence, expectations, and response options. Biol. Rev. Camb. Philos. Soc. 2017, 3, 12971313. [Google Scholar] [CrossRef]
- Pimentel, D.; Lach, L.; Zuniga, R.; Morrison, D. Environmental and economic costs of nonindigenous species in the United States. BioScience 2000, 50, 53–65. [Google Scholar] [CrossRef]
- Pintó-Marijuan, M.; Munné-Bosch, S. Ecophysiology of invasive plants: Osmotic adjustment and antioxidants. Trends Plant Sci. 2013, 18, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Khurshid, N.; Nasim, G.; Bajwa, K.; Adkins, S. Growth responses of Parthenium hysterophorus L. growing under salt stress. Pak. J. Weed Sci. Res. 2012, 18, 51–64. [Google Scholar]
- Onen, H.; Farooq, S.; Gunal, H.; Ozaslan, C.; Erdem, H. Higher Tolerance to abiotic stresses and soil types may accelerate common raag weed (Ambrosia artemisiifolia) invasion. Weed Sci. 2017, 65, 115–127. [Google Scholar] [CrossRef]
- Tanveer, M.; Shah, A.N. An insight into salt stress tolerance mechanisms of Chenopodium album. Environ. Sci. Pollut. Res. Int. 2017, 24, 16531–16535. [Google Scholar] [CrossRef]
- Yuan, X.; Wen, B. Seed germination response to high temperature and water stress in three invasive Asteraceae weeds from Xishuangbanna, SW China. PLoS ONE 2018, 13, e0191710. [Google Scholar] [CrossRef]
- Zeng, F.; Shabala, S.; Maksimović, J.D.; Maksimović, V.; Bonales-Alatorre, E.; Shabala, L.; Yu, M.; Zhang, G.; Živanović, B.D. Revealing mechanisms of salinity tissue tolerance in succulent halophytes: A case study for Carpobrotus rossi. Plant Cell Environ. 2018, 41, 2654–2667. [Google Scholar] [CrossRef]
- Santini, R.; De Lima, J.P.; Gratão, P.L.; Monteiro Camargo, A.F. Evaluation of growth and oxidative stress as indicative of salinity tolerance by the invasive tropical aquatic macrophyte tanner grass. Hydrobiologia 2022. [Google Scholar] [CrossRef]
- Türkan, I.; Demiral, T. Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 2009, 67, 2–9. [Google Scholar] [CrossRef]
- Koyro, H.W.; Ahmad, P.; Gessisler, N. Abiotic stress responses in plants: An Overview. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change 2012; Ahmad, P., Prsasad, M.N.V., Eds.; Springer Science+Business Media, LLC: New York, NY, USA, 2012; pp. 1–28. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Huang, S.; Van Aken, O.; Schwarzländer, M.; Belt, K.; Millar, A.H. The roles of mitochondrial reactive oxygen species in cellular signaling and stress response in plants. Plant Physiol. 2016, 171, 1551–1559. [Google Scholar] [CrossRef]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPPC). Summary for Policymakers. In Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Edenhofer, O., Pichs-Madruga, R., Sokona, Y., Farahani, E., Kadner, S., Seyboth, K., Adler, A., Baum, I., Brunner, S., Eickemeier, P., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2015. [Google Scholar]
- Coombs, G.; Peter, C.I. The invasive ‘mothcatcher’ (Araujia sericifera Brot.; Asclepiadoideae) co-opts native honeybees as its primary pollinator in South Africa. AoB Plants 2010, 2010, plq021. [Google Scholar] [CrossRef]
- Sans Elorza, M.; Dana Sánchez, E.D.; Sobrino Vesperinas, E. (Eds.) Plantas alóctonas invasoras en España; Dirección General para la Biodiversidad: Madrid, Spain, 2004; p. 384. [Google Scholar]
- Gaig, P.; Gázquez, V.; Lombardero, M.; Botey, E.; García-Ortega, P. Moth plant (Araujia sericifera) allergy. Allergy 2005, 60, 1092–1093. [Google Scholar] [CrossRef]
- Forster, P.I.; Bruyns, P.V. Clarification of synonymy for the common moth-vine Araujia sericifera (Asclepiadaceae). Taxon 1992, 41, 746–749. [Google Scholar] [CrossRef]
- Zuloaga, F.; Morrone, O.; Belgrano, M. Catálogo de las Plantas Vasculares del Cono Sur (Argentina, sur de Brasil, Chile, Paraguay y Uruguay); Missouri Botanical Garden: Sant Louis, MI, USA, 2008; Volumes 1–3. [Google Scholar]
- D’Errico, G.; Crescenzi, A.; Landi, S. First report of the southern root-knot nematode Meloidogyne incognita on the invasive weed Araujia sericifera in Italy. Plant Dis. 2014, 98, 15931593. [Google Scholar] [CrossRef]
- Santa Cruz, J.; Cordero, S. First record of Araujia sericifera (Apocynaceae: Asclepiadoideae) for Chile, a new alien climbing species from South America. B Soc. Argent. Bot. 2018, 53, 313–317. [Google Scholar] [CrossRef]
- Gómez de Barreda, A. Araujia sencifera Brot., mala hierba trepadora en los agrios españoles. Levante Agric. 1976, 205, 13–15. [Google Scholar]
- Gómez de Barreda Ferraz, D.; De Luca, V.; Osca, J.M.; Verdeguer, M.; Muñoz, M. Control químico de la mala hierba invasora Araujia sericifera Brot. ITEA Inf. Tec. Econ. Agrar. 2020, 116, 212–226. [Google Scholar] [CrossRef]
- Ward, B.G.; Henzell, R.F.; Holland, P.T.; Spiers, A.G. Non-sprays methods to control invasive weeds in urban areas. In Proceedings of the 52nd New Zealand Plant Protection Conference, Auckland, New Zealand, 10–12 August 1999; pp. 1–5. [Google Scholar]
- Oddo, M. Effects of Different Weed Control Practices on Soil Quality in Mediterranean Crops. Ph.D. Dissertation, Universitat Politècnica de València, Valencia, Spain, 2 October 2017. [Google Scholar]
- Ben-Gal, A.; Neori-Borochov, H.; Yermiyahu, U.; Shani, U. Is osmotic potential a more appropriate property than electrical conductivity for evaluating whole plant response to salinity? Environ. Exp. Bot. 2009, 65, 232–237. [Google Scholar] [CrossRef]
- Ellis, R.A.; Roberts, E.H. The quantification of aging and survival in orthodox seeds. Seed Sci. Technol. 1981, 9, 373–409. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil; California Agriculture Experiment Station, College of Agriculture, University of California: Berkeley, CA, USA, 1950; Volume Circular 347, pp. 1–32. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. T 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Weimberg, R. Solute adjustments in leaves of two species of wheat at two different stages of growth in response to salinity. Physiol. Plantarum 1987, 70, 381–388. [Google Scholar] [CrossRef]
- Hodges, D.M.; Delong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Blainski, A.; Lopes, G.C.; Palazzodemello, J.C. Application and analysis of the Folin Ciocalteu method for the determination of the total phenolic content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beyer, W.F., Jr.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–888. [Google Scholar]
- Conell, J.P.; Mullet, J.E. Pea chloroplast glutathione reductase: Purification and characterization. Plant Physiol. 1986, 82, 351–356. [Google Scholar] [CrossRef]
- Campoy, J.G.; Lema, M.; Fenollosa, E.; Munné-Bosch, S.; Retuerto, R. Functional responses to climate change may increase invasive potential of Carpobrotus edulis. Am. J. Bot. 2021, 108, 1902–1916. [Google Scholar] [CrossRef]
- Walther, G.-R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pysek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef]
- Mason, R.A.B.; Cooke, J.; Moles, A.T.; Leishman, M.R. Reproductive output of invasive versus native plants. Glob. Ecol. Biogeogr. 2008, 17, 633–640. [Google Scholar] [CrossRef]
- Vivian-Smith, G.; Panetta, F.D. Seedling recruitment, seed persistence and aspects of dispersal ecology of the invasive moth vine, Araujia sericifera (Asclepiadaceae). Aust. J. Bot. 2005, 53, 225–230. [Google Scholar] [CrossRef]
- Flowers, T.J.; Hajibagheri, M.A.; Clipson, N.J.W. Halophytes. Q. Rev. Biol. 1986, 61, 313–337. [Google Scholar] [CrossRef]
- Ungar, I.A. Seed germination and seed-bank ecology in halophytes. In Seed Development and Seed Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 599–628. [Google Scholar]
- Gul, B.; Ansari, R.; Flowers, T.J.; Khan, M.A. Germination strategies of halophyte seeds under salinity. Environ. Exp. Bot. 2013, 92, 4–18. [Google Scholar] [CrossRef]
- Alhaddad, F.A.; Abu-Dieyeh, M.H.; ElAzazi, E.-S.M.; Ahmed, T.A. Salt tolerance of selected halophytes at the two initial growth stages for future management options. Sci. Rep. 2021, 11, 10194. [Google Scholar] [CrossRef] [PubMed]
- Vicente, O.; Boscaiu, M.; Naranjo, M.; Estrelles, E.; Bellés, J.M.; Soriano, P. Responses to salt stress in the halophyte Plantago crassifolia (Plantaginaceae). J. Arid Environ. 2004, 58, 463481. [Google Scholar] [CrossRef]
- Yupsanis, T.; Moustakas, M.; Eleftherrious, P.; Damalanidou, K. Protein phosporylation dephosphorylation in alfalfa seeds germinating under salt stress. J. Plant Physiol. 1994, 143, 234–240. [Google Scholar] [CrossRef]
- Hohl, M.; Schopfer, P. Water relations of growing maize coleoptiles. Comparison between mannitol and polyethylene glycol 6000 as external osmotica for adjusting turgor pressure. Plant Physiol. 1991, 95, 716–722. [Google Scholar] [CrossRef]
- Soriano, P.; Moruno, F.; Boscaiu, M.; Vicente, O.; Hurtado, A.; Llinares, J.V.; Estrelles, E. Is salinity the main ecologic factor that shapes the distribution of two endemic Mediterranean plant species of the genus Gypsophila? Plant Soil 2014, 384, 363–379. [Google Scholar] [CrossRef]
- Cicevan, R.; Al Hassan, M.; Sestras, A.; Boscaiu, M.; Zaharia, A.; Vicente Meana, Ó.; Sestras, R. Comparative analysis of osmotic and ionic stress effects on seed germination in Tagetes (Asteraceae) cultivars. Propag. Ornam. Plants 2015, 15, 63–72. [Google Scholar]
- Katembe, W.J.; Ungar, I.A.; Mitchell, J.P. Effect of salinity on germination and seedling growth of two Atriplex species (Chenopodiaceae). Ann. Bot. 1998, 82, 167–175. [Google Scholar] [CrossRef]
- Borsai, O.; Al Hassan, M.; Boscaiu, M.; Sestras, R.E.; Vicente, O. Effects of salt and drought stress on seed germination and seedling growth in Portulaca. Rom. Biotechnol. Lett. 2018, 23, 13340–13349. [Google Scholar]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Keiffer, C.H.; Ungar, I.A. The effect of extended exposure to hypersaline conditions on the germination of five inland halophyte species. Am. J. Bot. 1997, 84, 104–111. [Google Scholar] [CrossRef]
- Nisar, F.; Gul, B.; Ajmal Khan, M.; Hameed, A. Germination and recovery responses of heteromorphic seeds of two co-occurring Arthrocnemum species to salinity, temperature and light. S. Afr. J. Bot. 2019, 121, 143–151. [Google Scholar] [CrossRef]
- Lambers, H.; Poorter, H. Inherent variation in growth rate between higher plants: A search for physiological causes and ecological consequences. Adv. Ecol. Res. 1992, 23, 188–261. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef]
- Al Hassan, M.; Pacurar, A.; López-Gresa, M.P.; Donat-Torres, M.P.; Llinares, J.V.; Boscaiu, M.; Vicente, O. Effects of salt stress on three ecologically distinct Plantago species. PLoS ONE 2016, 11, e0160236. [Google Scholar] [CrossRef]
- Greenway, H.; Munns, R. Mechanisms of salt tolerance in non-halophytes. Annu. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in Glycophytes. Front Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Morais, M.C.; Panuccio, M.R.; Muscolo, A.; Freitas, H. Salt tolerance traits increase the invasive success of Acacia longifolia in Portuguese coastal dunes. Plant Physiol. Biochem. 2012, 55, 60–65. [Google Scholar] [CrossRef]
- Kaur, T.; Bhat, J.A.; Raina, A.; Koul, S.; Vyas, D. Glutathione regulates enzymatic antioxidant defense with differential thiol content in perennial pepperweed and helps adapting to extreme environment. Acta Physiol. Plant. 2013, 35, 2501–2511. [Google Scholar] [CrossRef]
- Oliveira, M.T.; Matzek, V.; Dias Medeiros, C.; Rivas, R.; Falcão, H.M.; Santos, M.G. Stress tolerance and ecophysiological ability of an invader and a native species in a seasonally dry tropical forest. PLoS ONE 2014, 9, e105514. [Google Scholar] [CrossRef]
- Mamik, S.; Sharma, A.D. Protective role of boiling stable antioxidant enzymes in invasive alien species of Lantana exposed to natural abiotic stress like conditions. Russ. J. Biol. Invasions 2017, 8, 75–86. [Google Scholar] [CrossRef]
- Hussain, M.I.; González, L.; Souto, C.; Reigosa, M.J. Ecophysiological responses of three native herbs to phytotoxic potential of invasive Acacia melanoxylon R. Br. Agroforest. Syst. 2011, 83, 149–166. [Google Scholar] [CrossRef]
- Güçlütürk, I.; Detsi, A.; Weiss, E.K.; Ioannou, E.; Roussis, V.; Kefalas, P. Evaluation of anti-oxidant activity and identification of major polyphenolics of the invasive weed Oxalis pes-caprae. Phytochem. Anal. 2012, 23, 642–646. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Kapusta, P.; Stanek, M.; Frąc, M.; Oszust, K.; Woch, M.W.; Zubek, S. Invasive plant Reynoutria japonica produces large amounts of phenolic compounds and reduces the biomass but not activity of soil microbial communities. Sci. Total Environ. 2021, 767, 145439. [Google Scholar] [CrossRef]
- Bello, R.; Barrachina, M.; Esplugues, J.; López de Medrano-Villar, M.J.; Primo-Yúfera, E. Analgesic activity and effects on isolated smooth muscle of different fractions of hexane extract from Araujia sericifera Brot. Phytother. Res. 1996, 10, 337–339. [Google Scholar] [CrossRef]
- Bucciarelli, A.; Lofiego, A.; Bensack, A.; Rihl, C.; Skliar, M. Gastroprotective activity of Araujia sericifera Brot. var. hortorum (E. Fourn.), a South American species of medicinal use. Rev. Asoc. Med. Bahía Blanca 2017, 27, 30–36. [Google Scholar]
- Palomino-Schätzlein, M.; Montaño, M.C.; Escrig, P.V.; Boira, H.; Corma, A.; Pineda-Lucena, A.; Primo, J.; Cabedo, N. Identification of bioactive compounds in polar and nonpolar extracts of Araujia sericifera. Planta Med. Int. Open 2017, 4, e93–e103. [Google Scholar] [CrossRef][Green Version]
- Di Santo Meztler, P.; Fait, M.E.; Foresti, M.L.; Morcelle, S.R. Biocatalytic characterization of a naturally immobilized lipase found in Araujia sericifera Brot. (Apocynaceae) latex. Catal. Sci. Technol. 2014, 4, 1386–1394. [Google Scholar] [CrossRef]
- Landi, N.; Ragucci, S.; Letizia, F.; Fuggi, A.; Russo, R.; Pedone, P.V.; Di Maro, A. A haem-peroxidase from the seeds of Araujia sericifera: Characterization and use as bio-tool to remove phenol from aqueous solutions. Biocatal. Agric. Biotechnol. 2019, 20, 101215. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 372, 1331–1341. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS homeostasis in halophytes in the context of salinity stress tolerance. J. Exp. Bot. 2014, 65, 1241–1257. [Google Scholar] [CrossRef] [PubMed]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35 (Suppl. 1), 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate peroxidase and catalase activities and their genetic regulation in plants subjected to drought and salinity stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Control | Water Stress | 150 mM NaCl | 300 mM NaCl | 450 mM NaCl | 600 mM NaCl |
|---|---|---|---|---|---|---|
| Moisture | 68.48 ± 0.54 c | 9.79 ± 0.46 a | 67.97 ± 2.80 bc | 64.62 ± 3.12 bc | 66.37 ± 5.10 bc | 63.40 ± 1.58 b |
| EC1:5 | 1.07 ± 0.07 a | 0.54 ± 0.04 a | 9.45 ± 0.19 b | 19.96 ± 0.57 c | 20.27 ± 0.38 d | 26.6 ± 0.37 d |
| Parameter | Control | Water Stress | 150 mM NaCl | 300 mM NaCl | 450 mM NaCl | 600 mM NaCl |
|---|---|---|---|---|---|---|
| SL (cm) | 7.73 ± 0.87 c | 3.66 ± 0.25 a | 5.27 ± 0.53 b | 3.47 ± 0.24 a | 3.61 ± 0.20 a | 3.12 ± 0.24 a |
| Lno (units) | 6.86 ± 0.48 c | 4.42 ± 0.63 b | 4.29 ± 0.22 b | 3.00 ± 0.52 a | 4.14 ± 0.31 b | 4.00 ± 0.00 ab |
| LA (cm2) | 23.46 ± 2.39 b | 7.47 ± 1.13 a | 22.81 ± 1.99 b | 8.64 ± 1.68 a | 8.74 ± 2.52 a | 6.17 ± 1.36 a |
| Chl a (μg g−1DW) | 44.76 ± 3.48 d | 40.53 ± 4.90 cd | 30.83 ± 3.19 abc | 33.98 ± 3.71 bc | 28.08 ± 5.93 ab | 20.13 ± 3.77 a |
| Chl b (μg g−1DW) | 7.31 ± 0.91 c | 6.48 ± 1.23 bc | 4.62 ± 0.71 bc | 5.13 ± 0.76 ab | 4.65 ± 0.89 ab | 3.85 ± 0.74 a |
| Caro (μg g−1DW) | 4.58 ± 0.38 c | 4.50 ± 0.46 c | 2.90 ± 0.27 ab | 3.64 ± 0.45 bc | 3.18 ± 0.69 ab | 2.19 ± 0.51 a |
| Parameter | Control | Water Stress | 150 mM NaCl | 300 mM NaCl | 450 mM NaCl | 600 mM NaCl |
|---|---|---|---|---|---|---|
| MDA (nmol g−1DW) | 39.59 ± 10.72 a | 89.58 ± 3.79 b | 88.11 ± 4.27 b | 87.31 ± 8.37 b | 110.41 ± 13.84 bc | 123.87 ± 6.43 c |
| TPC (mg eq. GA g−1 DW) | 40.91 ± 1.50 a | 43.04 ± 3.20 ab | 44.74 ± 1.69 ab | 47.53 ± 4.18 ab | 49.44 ± 2.72 b | 58.52 ± 5.74 c |
| TF (mg eq. C g−1 DW) | 40.81 ± 4.65 a | 97.28 ± 5.68 c | 71.77 ± 9.02 b | 87.08 ± 7.78 bc | 100.05 ± 14.73 cd | 122.29 ± 6.96 d |
| SOD (Units mg−1 protein) | 9.17 ± 2.49 a | 10.20 ± 2.16 a | 22.66 ± 6.32 b | 26.87 ± 4.23 bc | 31.45 ± 5.66 bc | 37.19 ± 5.28 c |
| CAT (Units mg−1 protein) | 3.33 ± 2.37 a | 1.48 ± 0.87 a | 2.86 ± 1.79 a | 3.51 ± 2.12 a | 2.11 ± 0.98 a | 7.01 ± 2.03 a |
| APX (Units mg−1 protein) | 0.15 ± 0.01 a | 1.14 ± 0.15 b | 0.88 ± 0.13 ab | 1.58 ± 0.24 bc | 1.66 ± 0.33 bc | 2.36 ± 0.93 c |
| GR (Units mg−1 protein) | 47.46 ± 19.25 a | 57.48 ± 9.17 a | 88.14 ± 12.96 ab | 122.99 ± 5.33 bc | 126.27 ± 23.19 bc | 185.91 ± 26.56 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellache, M.; Moltó, N.; Benfekih, L.A.; Torres-Pagan, N.; Mir, R.; Verdeguer, M.; Boscaiu, M.; Vicente, O. Physiological and Biochemical Responses to Water Stress and Salinity of the Invasive Moth Plant, Araujia sericifera Brot., during Seed Germination and Vegetative Growth. Agronomy 2022, 12, 361. https://doi.org/10.3390/agronomy12020361
Bellache M, Moltó N, Benfekih LA, Torres-Pagan N, Mir R, Verdeguer M, Boscaiu M, Vicente O. Physiological and Biochemical Responses to Water Stress and Salinity of the Invasive Moth Plant, Araujia sericifera Brot., during Seed Germination and Vegetative Growth. Agronomy. 2022; 12(2):361. https://doi.org/10.3390/agronomy12020361
Chicago/Turabian StyleBellache, Manel, Nadia Moltó, Leila Allal Benfekih, Natalia Torres-Pagan, Ricardo Mir, Mercedes Verdeguer, Monica Boscaiu, and Oscar Vicente. 2022. "Physiological and Biochemical Responses to Water Stress and Salinity of the Invasive Moth Plant, Araujia sericifera Brot., during Seed Germination and Vegetative Growth" Agronomy 12, no. 2: 361. https://doi.org/10.3390/agronomy12020361
APA StyleBellache, M., Moltó, N., Benfekih, L. A., Torres-Pagan, N., Mir, R., Verdeguer, M., Boscaiu, M., & Vicente, O. (2022). Physiological and Biochemical Responses to Water Stress and Salinity of the Invasive Moth Plant, Araujia sericifera Brot., during Seed Germination and Vegetative Growth. Agronomy, 12(2), 361. https://doi.org/10.3390/agronomy12020361