


Starch Bio-Synthetic Pathway Genes Contribute to Resistant Starch Content Differentiation in Bread Wheat
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Evaluation of RS Content
2.3. Statistical Analysis
2.4. Genotyping of All Varieties and Quality Control
2.5. GWAS for RS Genetic Loci
2.6. KASP Primer Design and Genotype Detection
2.7. Candidate Gene Prediction
3. Results
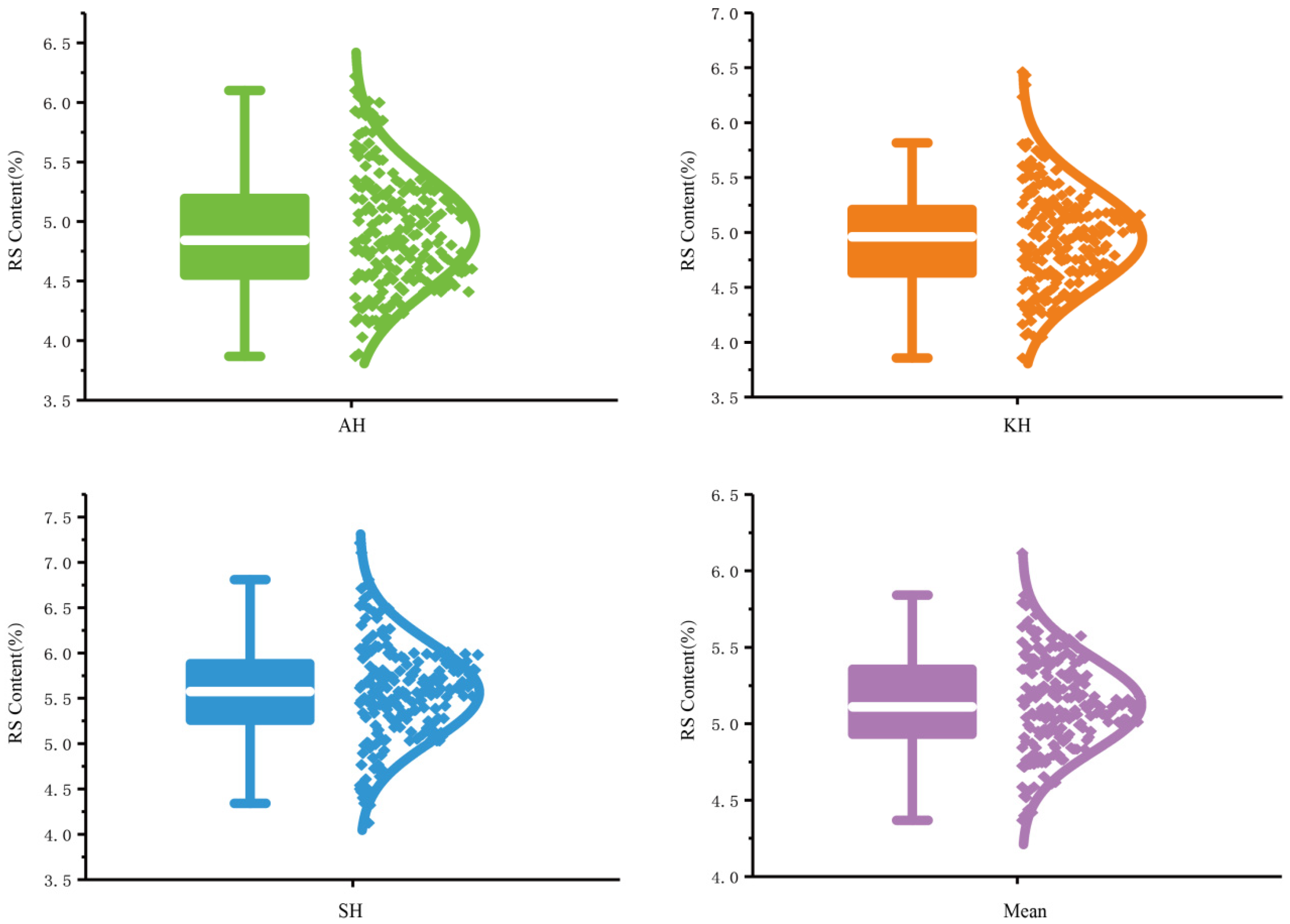
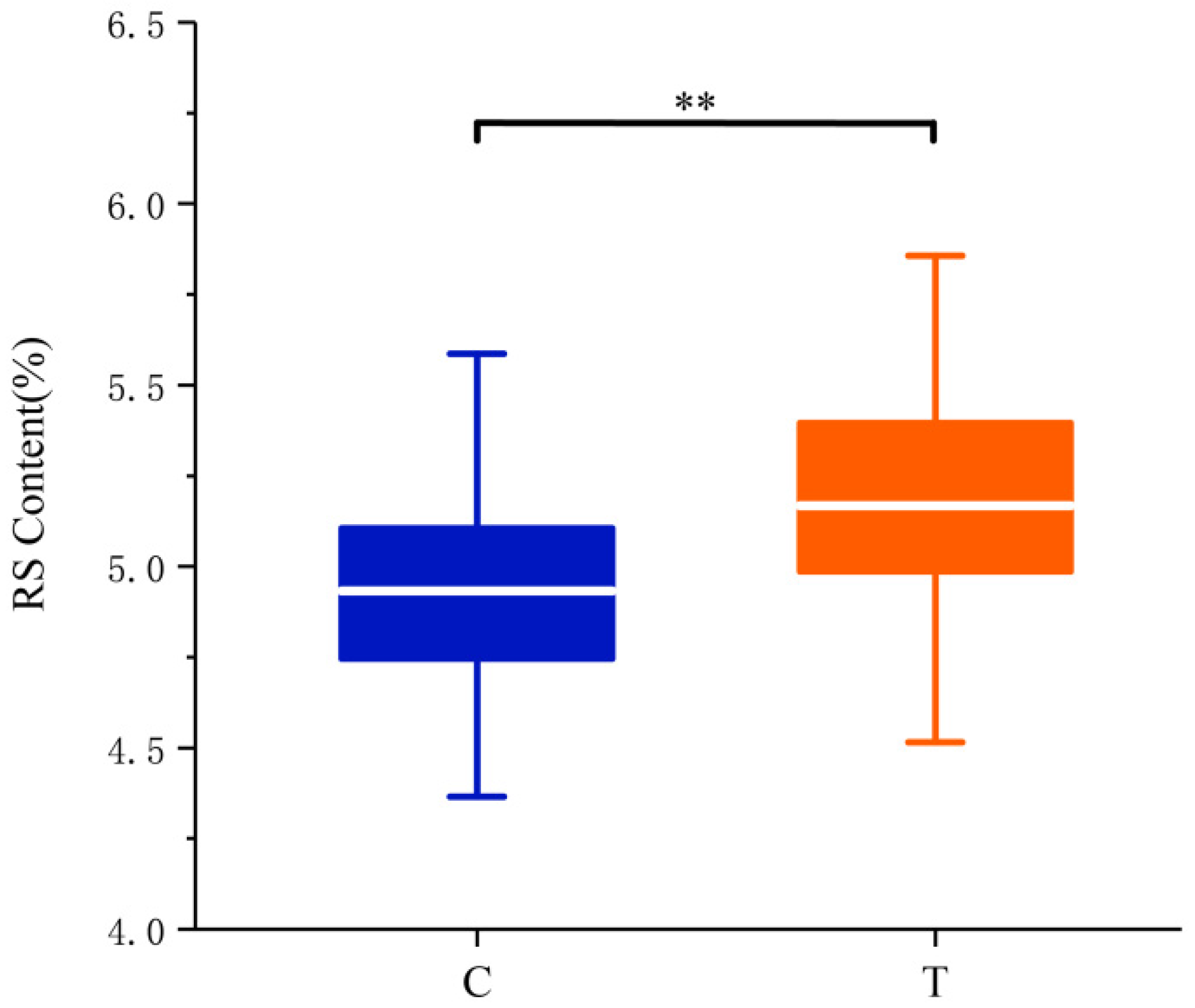
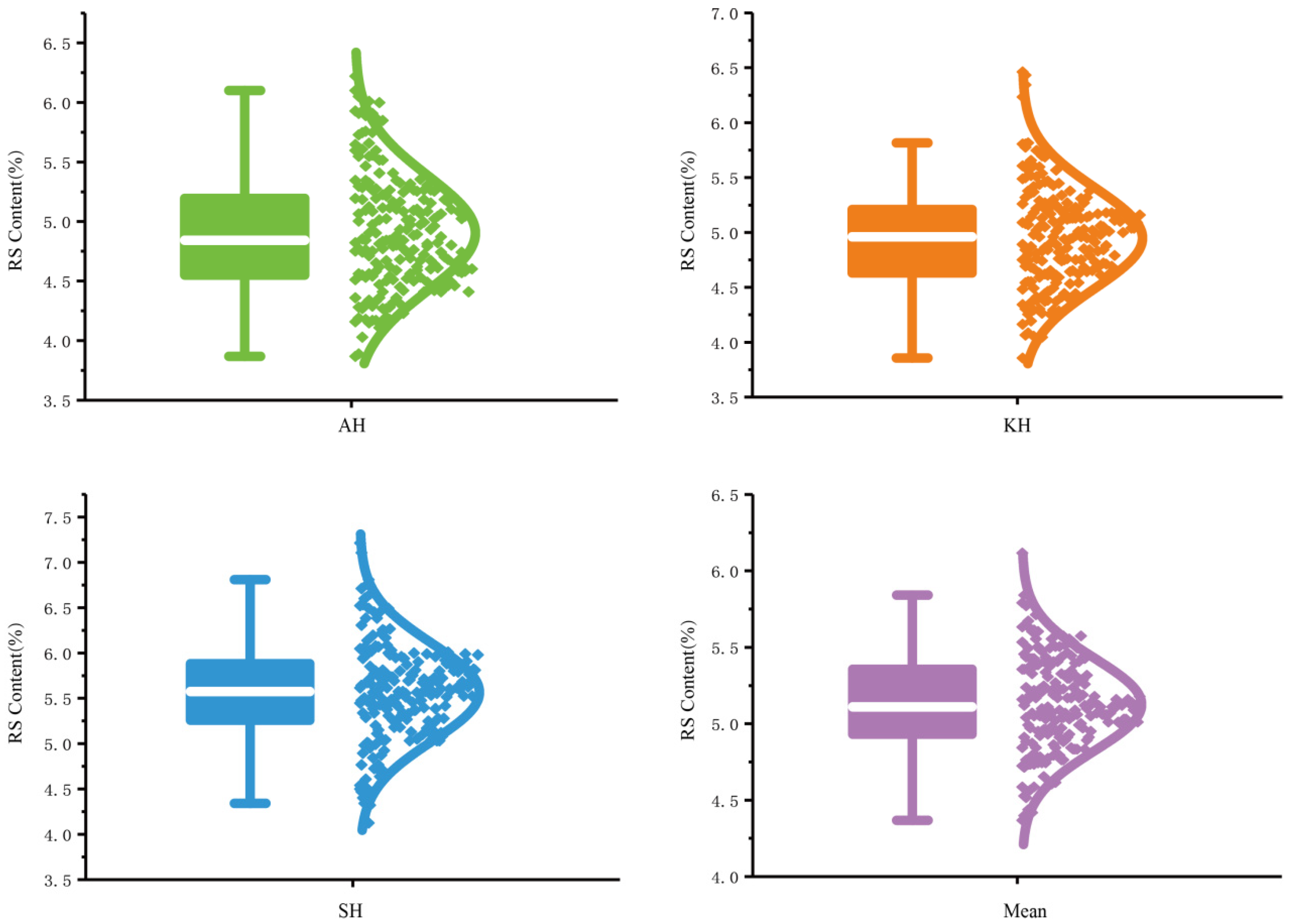
3.1. RS Content Evaluation and Phenotypic Distribution in the GWAS Population
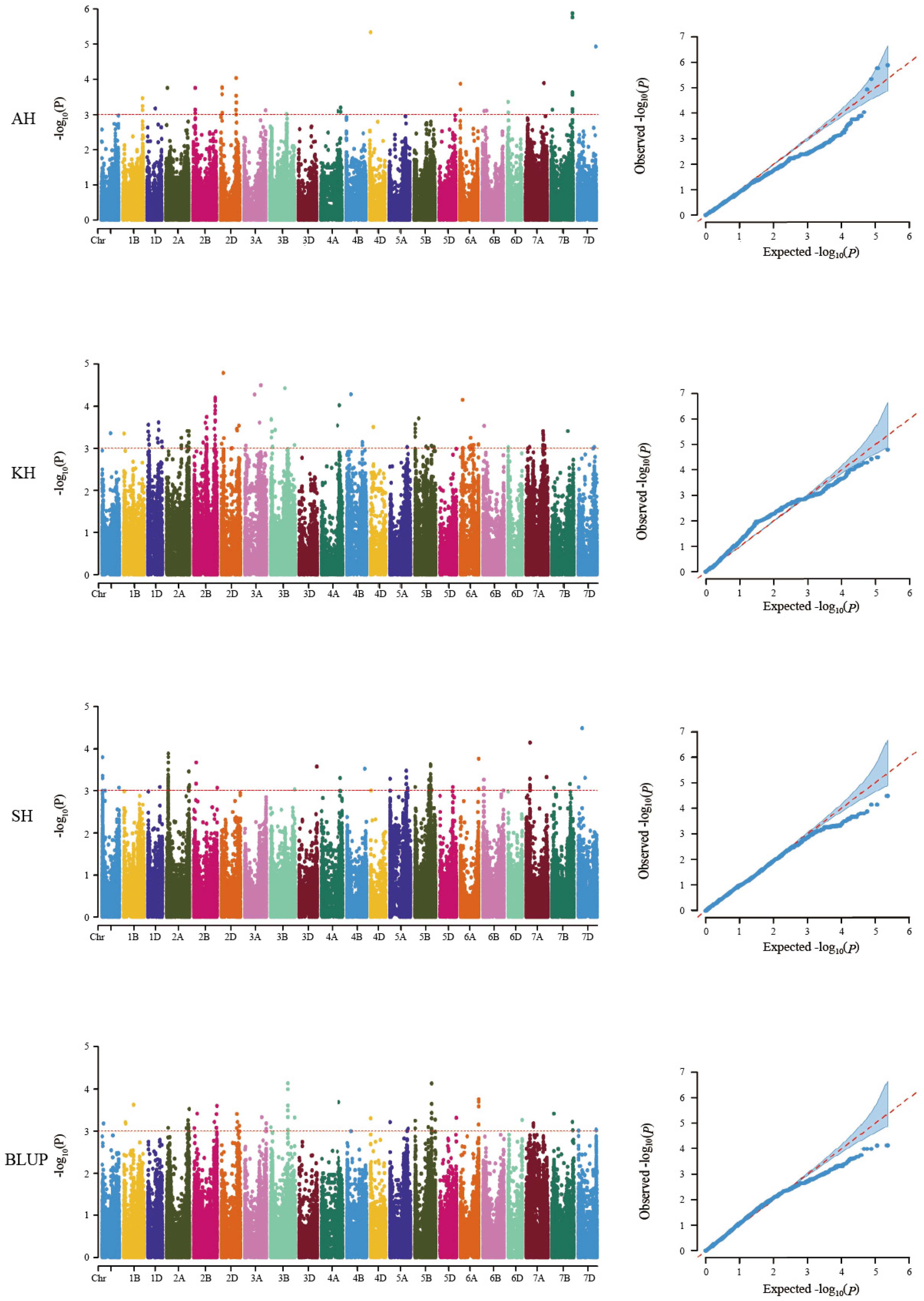
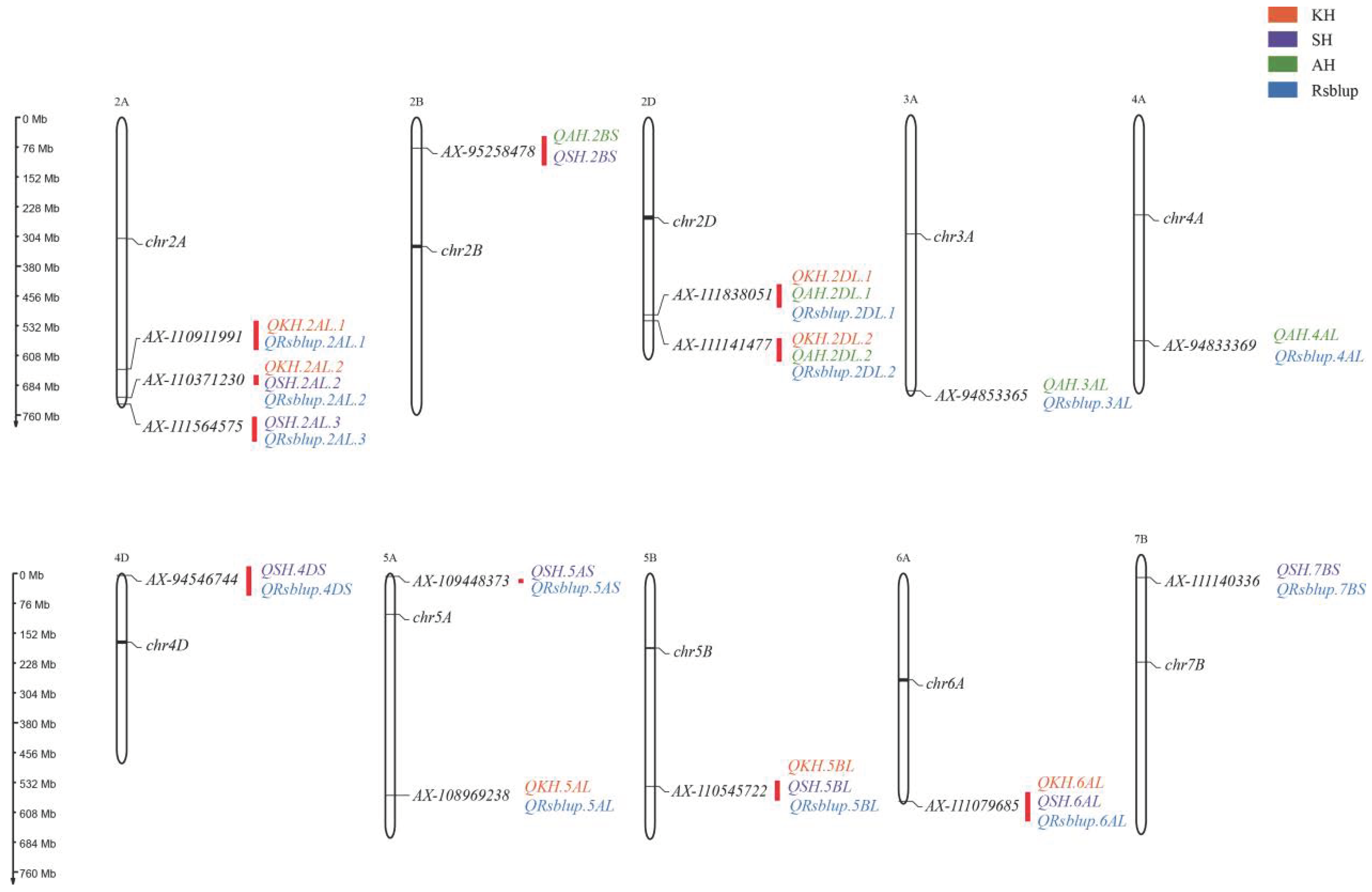
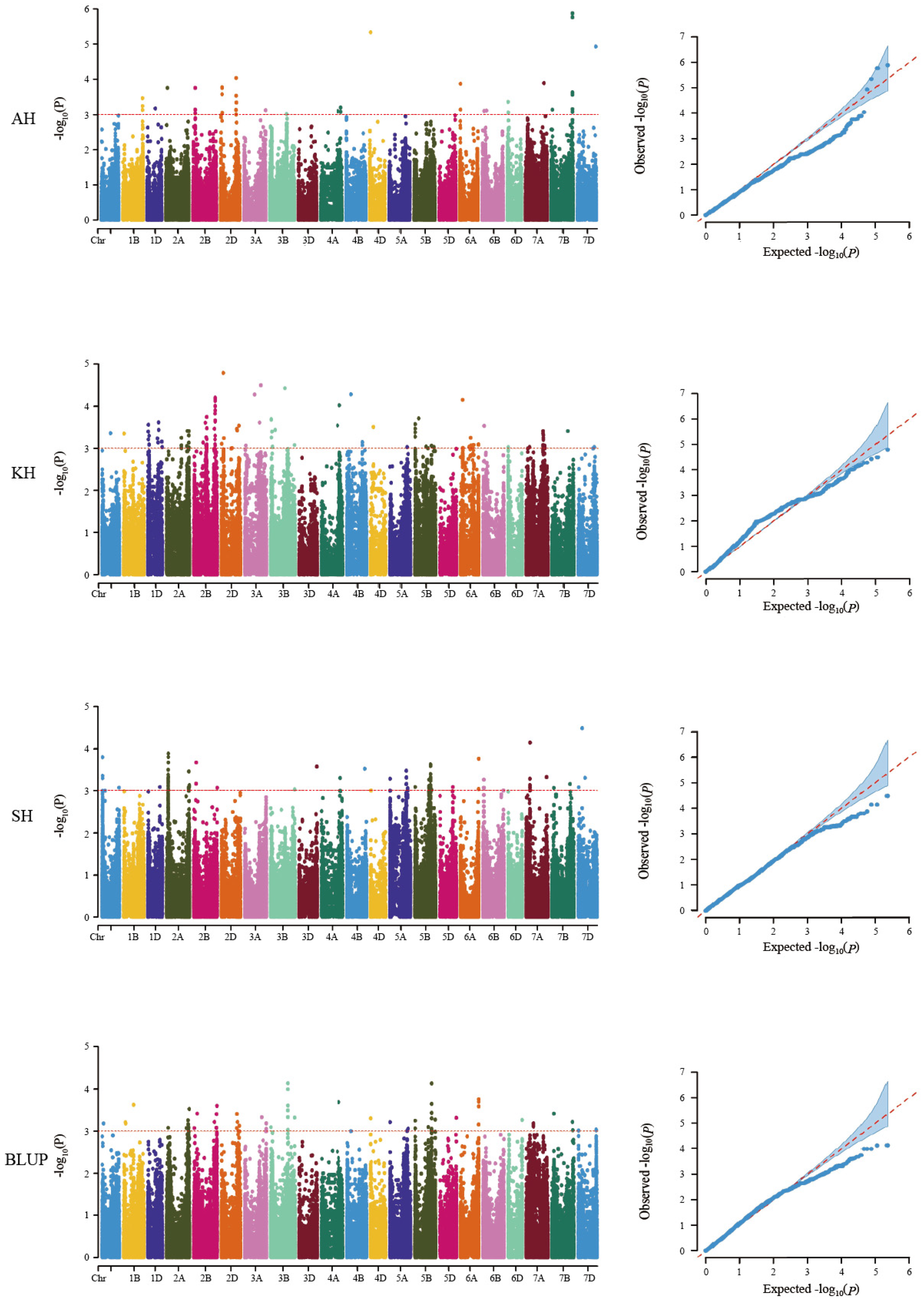
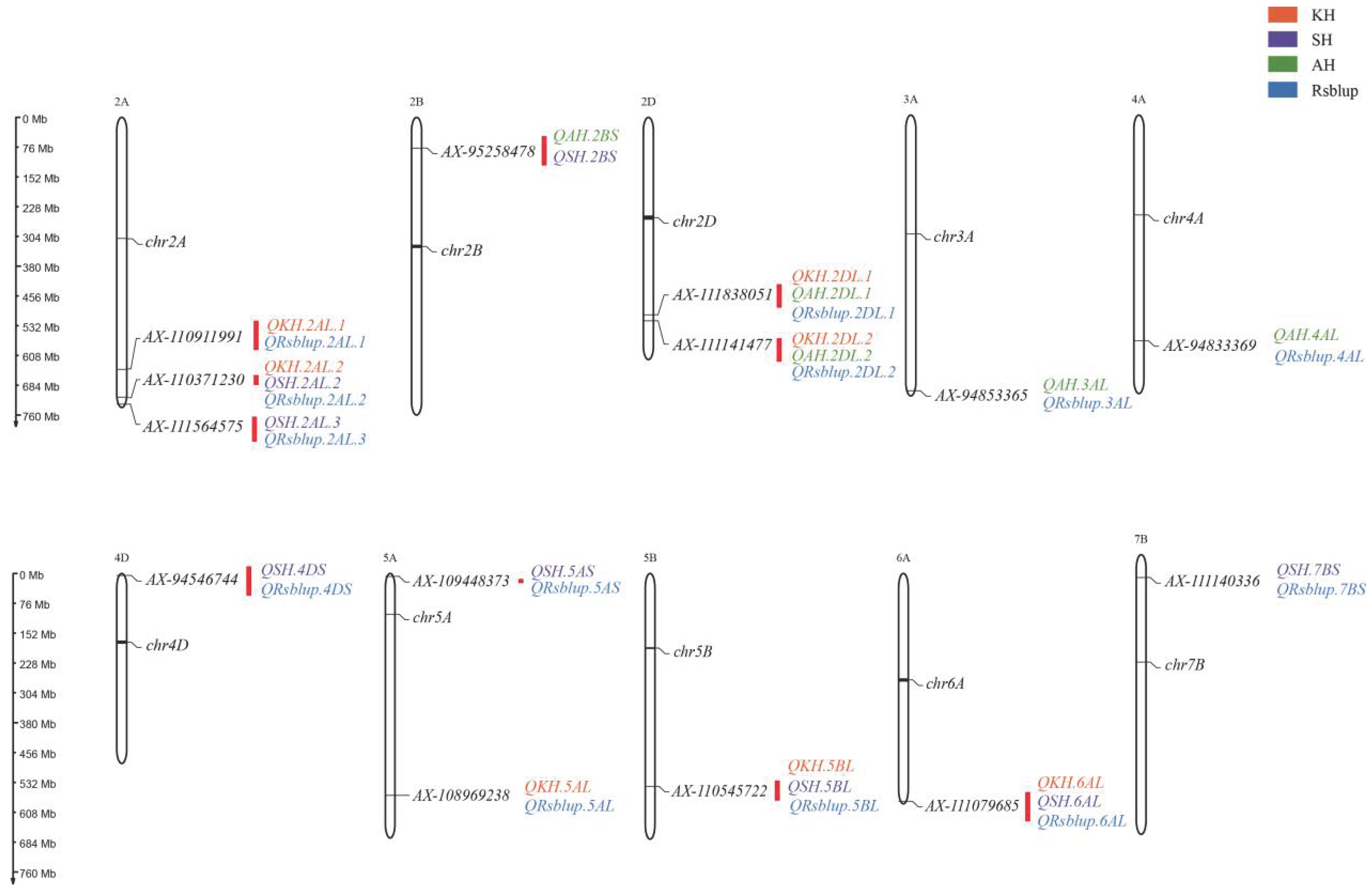
3.2. Detection of the Genetic Loci for RS Content
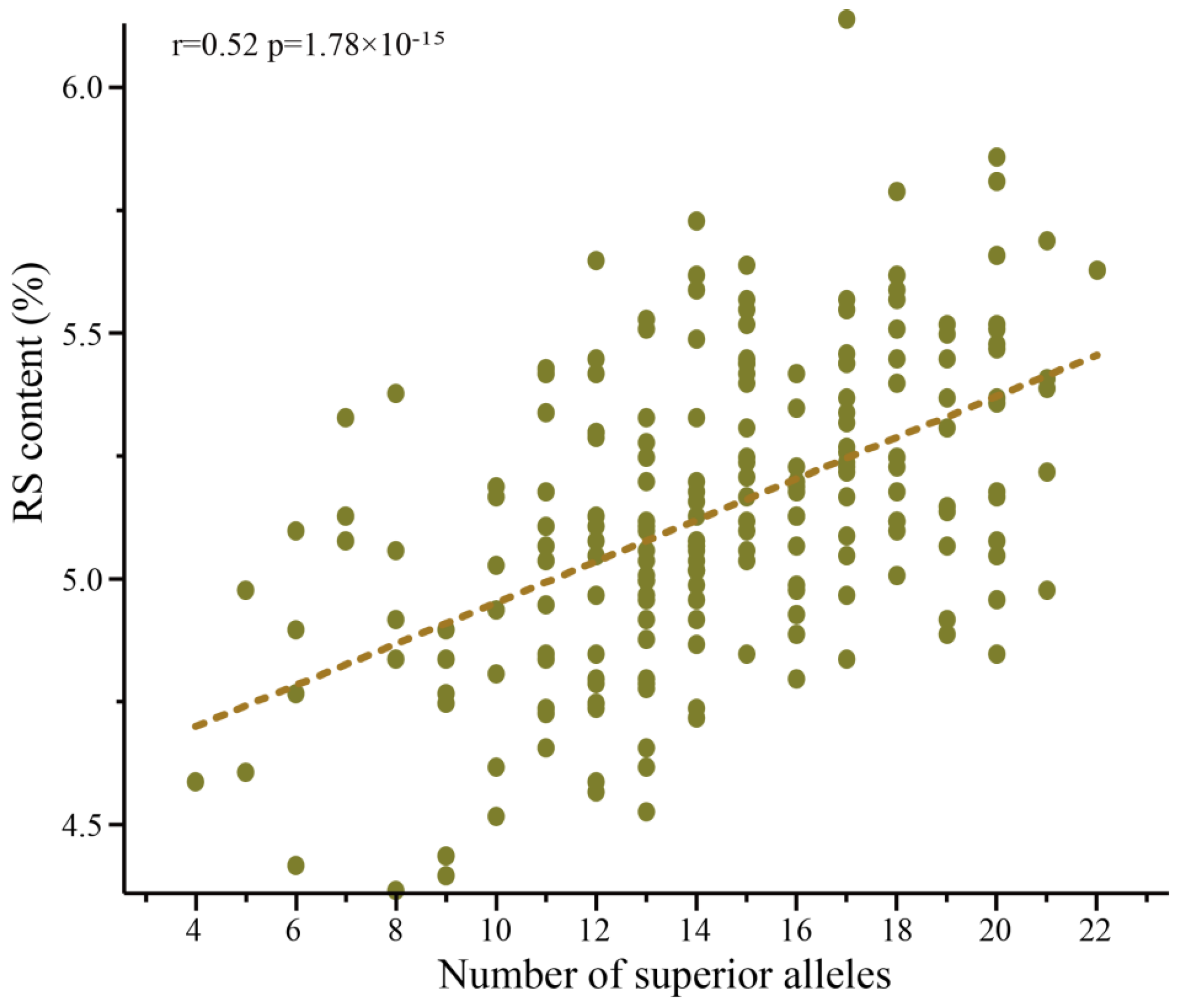
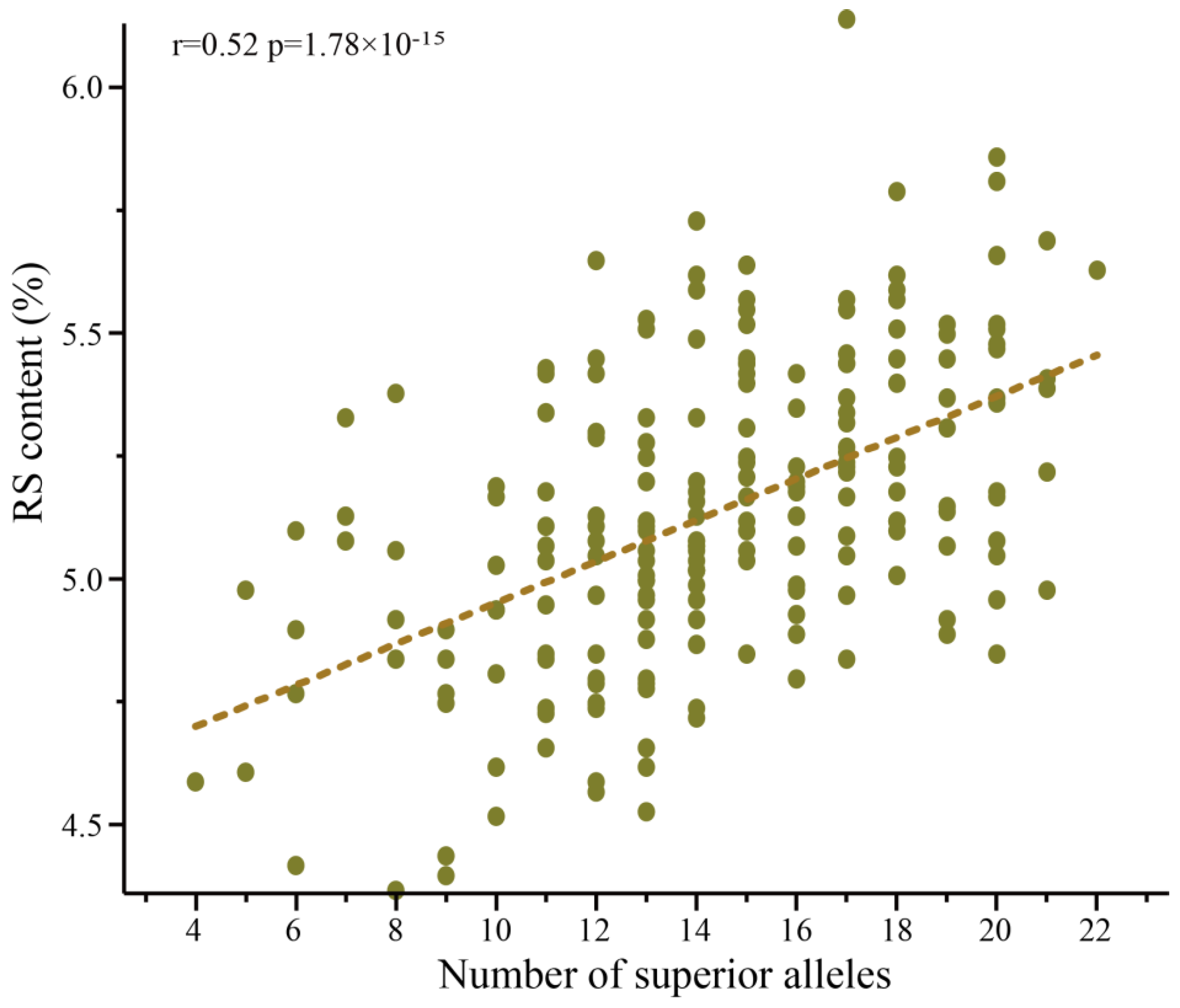
3.3. Additive Effect to the RS Content of the SNP
3.4. Candidate Gene Prediction
3.5. KASP Marker Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Huang, L.; Tan, H.; Zhang, C.; Li, Q.; Liu, Q. Starch biosynthesis in cereal endosperms: An updated review over the last decade. Plant Commun. 2021, 2, 100237. [Google Scholar] [CrossRef]
- Macneill, G.J.; Sahar, M.; Minow, M.A.A.; Patterson, J.A.; Tetlow, I.J.; Emes, M.J. Starch as a source, starch as a sink: The bifunctional role of starch in carbon allocation. J. Exp. Bot. 2017, 68, 4433–4453. [Google Scholar]
- Englyst, H.; Kingman, S.; Cummings, J. Classification and Measurement of Nutritionally Important Starch Fractions. Eur. J. Clin. Nutr. 1992, 46, S33–S50. [Google Scholar]
- Englyst, H.N.; Cummings, J.H. Digestion of the polysaccharides of some cereal foods in the human small intestine. Am. J. Clin. Nutr. 1985, 42, 778–787. [Google Scholar]
- Bin, S.; Yin, Y.; Li, T.; Huang, R.; Zhang, P. Analysis of the resistant starch content in cereals. Feed. Res. 2006, 2006, 2. [Google Scholar]
- Slade, A.J.; McGuire, C.; Loeffler, D.; Mullenberg, J.; Skinner, W.; Fazio, G.; Holm, A.; Brandt, K.M.; Steine, M.N.; Goodstal, J.F.; et al. Development of high amylose wheat through TILLING. BMC Plant Biol. 2012, 12, 69. [Google Scholar] [CrossRef] [Green Version]
- Birt, D.F.; Boylston, T.; Hendrich, S.; Jane, J.L.; Hollis, J.; Li, L.; McClelland, J.; Moore, S.; Phillips, G.J.; Rowling, M.; et al. Resistant starch: Promise for improving human health. Adv. Nutr. 2013, 4, 587–601. [Google Scholar] [CrossRef] [Green Version]
- Woo, K.S.; Seib, P.A. Cross-Linked Resistant Starch: Preparation and Properties. Cereal Chem. 2002, 79, 819–825. [Google Scholar]
- Harris, K.F. An introductory review of resistant starch type 2 from high-amylose cereal grains and its effect on glucose and insulin homeostasis. Nutr. Rev. 2019, 77, 748–764. [Google Scholar] [CrossRef] [Green Version]
- Meseguer Soler, I.; Martinez Para, M.C.; Farre Rovira, R. Dietary fiber (I). Definition, properties and composition. Med. Clin. 1997, 109, 641–644. [Google Scholar]
- Roman, L.; Martinez, M.M. Structural Basis of Resistant Starch (RS) in Bread: Natural and Commercial Alternatives. Foods 2019, 8, 267. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [Green Version]
- Ze, X.; Ben David, Y.; Laverde-Gomez, J.A.; Dassa, B.; Sheridan, P.O.; Duncan, S.H.; Louis, P.; Henrissat, B.; Juge, N.; Koropatkin, N.M.; et al. Unique Organization of Extracellular Amylases into Amylosomes in the Resistant Starch-Utilizing Human Colonic Firmicutes Bacterium Ruminococcus bromii. MBio 2015, 6, e01058–15. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Willett, W.C.; Manson, J.E.; Hu, F.B.; Rosner, B.; Colditz, G. Relation between changes in intakes of dietary fiber and grain products and changes in weight and development of obesity among middle-aged women. Am. J. Clin. Nutr. 2003, 78, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Peterson, C.M.; Beyl, R.A.; Marlatt, K.L.; Martin, C.K.; Aryana, K.J.; Marco, M.L.; Martin, R.J.; Keenan, M.J.; Eric, R. Effect of 12 wk of resistant starch supplementation on cardiometabolic risk factors in adults with prediabetes: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 492–501. [Google Scholar]
- Cook, T.M.; Mansuy-Aubert, V. Communication between the gut microbiota and peripheral nervous system in health and chronic disease. Gut Microbes 2022, 14, 2068365. [Google Scholar] [CrossRef]
- Corfield, A.P. The Interaction of the Gut Microbiota with the Mucus Barrier in Health and Disease in Human. Microorganisms 2018, 6, 78. [Google Scholar] [CrossRef]
- Kovatcheva-Datchary, P.; Egert, M.; Maathuis, A.; Rajilic-Stojanovic, M.; de Graaf, A.A.; Smidt, H.; de Vos, W.M.; Venema, K. Linking phylogenetic identities of bacteria to starch fermentation in an in vitro model of the large intestine by RNA-based stable isotope probing. Environ. Microbiol. 2009, 11, 914–926. [Google Scholar] [CrossRef]
- Zhang, L.; Ouyang, Y.; Li, H.; Shen, L.; Ni, Y.; Fang, Q.; Wu, G.; Qian, L.; Xiao, Y.; Zhang, J.; et al. Metabolic phenotypes and the gut microbiota in response to dietary resistant starch type 2 in normal-weight subjects: A randomized crossover trial. Sci. Rep. 2019, 9, 4736. [Google Scholar] [CrossRef] [Green Version]
- Scarpellini, E.; Balsiger, L.M.; Maurizi, V.; Rinninella, E.; Gasbarrini, A.; Giostra, N.; Santori, P.; Abenavoli, L.; Rasetti, C. Zinc and gut microbiota in health and gastrointestinal disease under the COVID-19 suggestion. Biofactors 2022, 48, 294–306. [Google Scholar] [CrossRef]
- Sanders, L.M.; Dicklin, M.R.; Palacios, O.M.; Maki, C.E.; Wilcox, M.L.; Maki, K.C. Effects of potato resistant starch intake on insulin sensitivity, related metabolic markers and appetite ratings in men and women at risk for type 2 diabetes: A pilot cross-over randomised controlled trial. J. Hum. Nutr. Diet. 2021, 34, 94–105. [Google Scholar] [CrossRef]
- Song, X.; Dong, H.; Zang, Z.; Wu, W.; Zhu, W.; Zhang, H.; Guan, Y. Kudzu Resistant Starch: An Effective Regulator of Type 2 Diabetes Mellitus. Oxid. Med. Cell. Longev. 2021, 2021, 4448048. [Google Scholar] [CrossRef]
- Murphy, M.M.; Douglass, J.S.; Birkett, A. Resistant starch intakes in the United States. J. Am. Diet. Assoc. 2008, 108, 67–78. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, P.; Xiao, Z. Resistant starch prevents tumorigenesis of dimethylhydrazine-induced colon tumors via regulation of an ER stress-mediated mitochondrial apoptosis pathway. Int. J. Mol. Med. 2018, 41, 1887–1898. [Google Scholar] [CrossRef]
- Lafiandra, D.; Riccardi, G.; Shewry, P.R. Improving cereal grain carbohydrates for diet and health. J. Cereal Sci. 2014, 59, 312–326. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Jiao, G.; Sun, Y.; Chen, J.; Zhong, Y.; Yan, L.; Jiang, D.; Ma, Y.; Xia, L. Modification of starch composition, structure and properties through editing of TaSBEIIa in both winter and spring wheat varieties by CRISPR/Cas9. Plant Biotechnol. J. 2021, 19, 937–951. [Google Scholar] [CrossRef]
- Mishra, A.; Singh, A.; Sharma, M.; Kumar, P.; Roy, J. Development of EMS-induced mutation population for amylose and resistant starch variation in bread wheat (Triticum aestivum) and identification of candidate genes responsible for amylose variation. BMC Plant Biol. 2016, 16, 217. [Google Scholar] [CrossRef]
- Qu, J.; Xu, S.; Zhang, Z.; Chen, G.; Zhong, Y.; Liu, L.; Zhang, R.; Xue, J.; Guo, D. Evolutionary, structural and expression analysis of core genes involved in starch synthesis. Sci. Rep. 2018, 8, 12736. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Wu, Z.L.; Xing, Y.Y.; Zheng, F.G.; Guo, X.L.; Zhang, W.G.; Hong, M.M. Nucleotide sequence of rice waxy gene. Nucleic Acids Res. 1990, 18, 5898. [Google Scholar] [CrossRef] [Green Version]
- Streb, S.; Delatte, T.; Umhang, M.; Eicke, S.; Schorderet, M.; Reinhardt, D.; Zeeman, S.C. Starch granule biosynthesis in Arabidopsis is abolished by removal of all debranching enzymes but restored by the subsequent removal of an endoamylase. Plant Cell 2008, 20, 3448–3466. [Google Scholar] [CrossRef] [Green Version]
- Yun, M.S.; Umemoto, T.; Kawagoe, Y. Rice debranching enzyme isoamylase3 facilitates starch metabolism and affects plastid morphogenesis. Plant Cell Physiol. 2011, 52, 1068–1082. [Google Scholar] [CrossRef] [Green Version]
- Nishi, A.; Nakamura, Y.; Tanaka, N.; Satoh, H. Biochemical and genetic analysis of the effects of amylose-extender mutation in rice endosperm. Plant Physiol. 2001, 127, 459–472. [Google Scholar]
- Regina, A.; Bird, A.; Topping, D.; Bowden, S.; Freeman, J.; Barsby, T.; Kosar-Hashemi, B.; Li, Z.; Rahman, S.; Morell, M. High-amylose wheat generated by RNA interference improves indices of large-bowel health in rats. Proc. Natl. Acad. Sci. USA 2006, 103, 3546–3551. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.H.; Bergman, C.J.; McClung, A.M.; Everette, J.D.; Tabien, R.E. Resistant starch: Variation among high amylose rice varieties and its relationship with apparent amylose content, pasting properties and cooking methods. Food Chem. 2017, 234, 180–189. [Google Scholar]
- Yadav, B.S.; Sharma, A.; Yadav, R.B. Resistant starch content of conventionally boiled and pressure-cooked cereals, legumes and tubers. J. Food Sci. Technol. 2010, 47, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Schoen, A.; Joshi, A.; Tiwari, V.; Gill, B.S.; Rawat, N. Triple null mutations in starch synthase SSIIa gene homoeologs lead to high amylose and resistant starch in hexaploid wheat. BMC Plant Biol. 2021, 21, 74. [Google Scholar] [CrossRef]
- Fahy, B.; Gonzalez, O.; Savva, G.M.; Ahn-Jarvis, J.H.; Warren, F.J.; Dunn, J.; Lovegrove, A.; Hazard, B.A. Loss of starch synthase IIIa changes starch molecular structure and granule morphology in grains of hexaploid bread wheat. Sci. Rep. 2022, 12, 10806. [Google Scholar] [CrossRef]
- Sparla, F.; Falini, G.; Botticella, E.; Pirone, C.; Talame, V.; Bovina, R.; Salvi, S.; Tuberosa, R.; Sestili, F.; Trost, P. New starch phenotypes produced by TILLING in barley. PLoS ONE 2014, 9, e107779. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Kim, J.Y. Understanding Wheat Starch Metabolism in Properties, Environmental Stress Condition, and Molecular Approaches for Value-Added Utilization. Plants 2021, 10, 2282. [Google Scholar] [CrossRef]
- Foschia, M.; Beraldo, P.; Peressini, D. Evaluation of the physicochemical properties of gluten-free pasta enriched with resistant starch. J. Sci. Food Agric. 2017, 97, 572–577. [Google Scholar] [CrossRef]
- Fiszman, S.M.; Salvador, A.; Baixauli, R.; Sanz, T. Muffins with resistant starch: Baking performance in relation to the rheological properties of the batter. J. Cereal Sci. 2008, 47, 502–509. [Google Scholar]
- Rohlfing, K.A.; Paez, A.; Kim, H.J.; White, P.J. Effects of Resistant Starch and Fiber from High-Amylose Non-Floury Corn on Tortilla Texture. Cereal Chem. 2010, 87, 581–585. [Google Scholar]
- Korus, J.; Witczak, M.; Ziobro, R.; Juszczak, L. The impact of resistant starch on characteristics of gluten-free dough and bread. Food Hydrocoll. 2009, 23, 988–995. [Google Scholar]
- Keeling, P.; Banisadr, R.; Barone, L.; Wasserman, B.; Singletary, G. Effect of Temperature on Enzymes in the Pathway of Starch Biosynthesis in Developing Wheat and Maize Grain. Aust. J. Plant Physiol. 1994, 21, 807. [Google Scholar]
- Liu, C.; Zhou, Z.; Wu, Z.; Qin, M.; Shi, X.; Wang, Y.; Li, W.; Yao, W.; Lei, Z.; Zhao, R. In-depth genetic analysis reveals conditioning of polyphenol oxidase activity in wheat grains by cis regulation of TaPPO2A-1 expression level. Genomics 2020, 112, 4690–4700. [Google Scholar] [CrossRef]
- Zhou, Z.; Guan, H.; Liu, C.; Zhang, Z.; Geng, S.; Qin, M.; Li, W.; Shi, X.; Dai, Z.; Lei, Z.; et al. Identification of genomic regions affecting grain peroxidase activity in bread wheat using genome-wide association study. BMC Plant Biol. 2021, 21, 523. [Google Scholar] [CrossRef]
- Shi, X.; Zhou, Z.; Li, W.; Qin, M.; Yang, P.; Hou, J.; Huang, F.; Lei, Z.; Wu, Z.; Wang, J. Genome-wide association study reveals the genetic architecture for calcium accumulation in grains of hexaploid wheat (Triticum aestivum L.). BMC Plant Biol. 2022, 22, 229. [Google Scholar] [CrossRef]
- Zhou, Z.; Shi, X.; Zhao, G.; Qin, M.; Ibba, M.I.; Wang, Y.; Li, W.; Yang, P.; Wu, Z.; Lei, Z.; et al. Identification of Novel Genomic Regions and Superior Alleles Associated with Zn Accumulation in Wheat Using a Genome-Wide Association Analysis Method. Int. J. Mol. Sci. 2020, 21, 1928. [Google Scholar] [CrossRef] [Green Version]
- Mccleary, B.V.; Mcloughlin, C.; Charmier, L.; Mcgeough, P. Measurement of available carbohydrates, digestible and resistant starch in food ingredients and products. Cereal Chem. 2019, 97, 114–137. [Google Scholar]
- Rasheed; Awais; Liu; Jindong; Guo; Qi; Dreisigacker; Susanne; Zhai; Shengnan. Development and validation of KASP assays for genes underpinning key economic traits in bread wheat. Theor. Appl. Genet. Int. J. Breed. Res. Cell Genet. 2016, 129, 1843–1860. [Google Scholar]
- Seung, D.; Smith, A.M. Starch granule initiation and morphogenesis-progress in Arabidopsis and cereals. J. Exp. Bot. 2019, 70, 771–784. [Google Scholar] [CrossRef]
- Hwang, S.K.; Koper, K.; Satoh, H.; Okita, T.W. Rice Endosperm Starch Phosphorylase (Pho1) Assembles with Disproportionating Enzyme (Dpe1) to Form a Protein Complex That Enhances Synthesis of Malto-oligosaccharides. J. Biol. Chem. 2016, 291, 19994–20007. [Google Scholar] [CrossRef] [Green Version]
- Malinova, I.; Qasim, H.M.; Brust, H.; Fettke, J. Parameters of Starch Granule Genesis in Chloroplasts of Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 761. [Google Scholar] [CrossRef]
- Cuesta-Seijo, J.A.; Nielsen, M.M.; Ruzanski, C.; Krucewicz, K.; Beeren, S.R.; Rydhal, M.G.; Yoshimura, Y.; Striebeck, A.; Motawia, M.S.; Willats, W.G.; et al. In vitro Biochemical Characterization of All Barley Endosperm Starch Synthases. Front. Plant Sci. 2015, 6, 1265. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Ono, M.; Utsumi, C.; Steup, M. Functional interaction between plastidial starch phosphorylase and starch branching enzymes from rice during the synthesis of branched maltodextrins. Plant Cell Physiol. 2012, 53, 869–878. [Google Scholar] [CrossRef]
- Tetlow, I.J. Starch biosynthesis in developing seeds. Seed Sci. Res. 2011, 21, 5–32. [Google Scholar]
- Zhou, H.; Wang, L.; Liu, G.; Meng, X.; Jing, Y.; Shu, X.; Kong, X.; Sun, J.; Yu, H.; Smith, S.M.; et al. Critical roles of soluble starch synthase SSIIIa and granule-bound starch synthase Waxy in synthesizing resistant starch in rice. Proc. Natl. Acad. Sci. USA 2016, 113, 12844–12849. [Google Scholar] [CrossRef] [Green Version]
- Baysal, C.; He, W.; Drapal, M.; Villorbina, G.; Medina, V.; Capell, T.; Khush, G.S.; Zhu, C.; Fraser, P.D.; Christou, P. Inactivation of rice starch branching enzyme IIb triggers broad and unexpected changes in metabolism by transcriptional reprogramming. Proc. Natl. Acad. Sci. USA 2020, 117, 26503–26512. [Google Scholar] [CrossRef]
- Hayashi, M.; Crofts, N.; Oitome, N.F.; Fujita, N. Analyses of starch biosynthetic protein complexes and starch properties from developing mutant rice seeds with minimal starch synthase activities. BMC Plant Biol. 2018, 18, 59. [Google Scholar] [CrossRef] [Green Version]
- Abt, M.R.; Zeeman, S.C. Evolutionary innovations in starch metabolism. Curr. Opin. Plant Biol. 2020, 55, 109–117. [Google Scholar] [CrossRef]
- Oa, W.; Chloroplasts, A.; Smith, S.M.; Samuel, C. β-AMYLASE4, a Noncatalytic Protein Required for Starch Breakdown, Acts Upstream of Three Active β-Amylases in Arabidopsis Chloroplasts. Plant Cell 2008, 20, 1040–1058. [Google Scholar]
- Du, L.; Xu, F.; Fang, J.; Gao, S.; Tang, J.; Fang, S.; Wang, H.; Tong, H.; Zhang, F.; Chu, J.; et al. Endosperm sugar accumulation caused by mutation of PHS8/ISA1 leads to pre-harvest sprouting in rice. Plant J. 2018, 95, 545–556. [Google Scholar] [CrossRef] [Green Version]
- Weschke, W.; Panitz, R.; Sauer, N.; Wang, Q.; Neubohn, B.; Weber, H.; Wobus, U. Sucrose transport into barley seeds: Molecular characterization of two transporters and implications for seed development and starch accumulation. Plant J. 2000, 21, 455–467. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Hou, J.; Wang, X.; Li, T.; Majeed, U.; Hao, C.; Zhang, X. The NAC transcription factor NAC019-A1 is a negative regulator of starch synthesis in wheat developing endosperm. J. Exp. Bot. 2020, 71, 5794–5807. [Google Scholar] [CrossRef]
- Gao, Y.; An, K.; Guo, W.; Chen, Y.; Zhang, R.; Zhang, X.; Chang, S.; Vincenzo, R.; Jin, F.; Cao, X. The endosperm-specific transcription factor TaNAC019 regulates glutenin and starch accumulation and its elite allele improves wheat grain quality. Plant Cell 2021, 33, 603–622. [Google Scholar]
- Hennen-Bierwagen, T.A.; Lin, Q.; Grimaud, F.; Planchot, V.; Keeling, P.L.; James, M.G.; Myers, A.M. Proteins from multiple metabolic pathways associate with starch biosynthetic enzymes in high molecular weight complexes: A model for regulation of carbon allocation in maize amyloplasts. Plant Physiol. 2009, 149, 1541–1559. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Li, J.; Li, Y. Resistant starch formation in rice: Genetic regulation and beyond. Plant Commun. 2022, 3, 100329. [Google Scholar] [CrossRef]
- Hirano, H.Y.; Sano, Y. Enhancement of Wx gene expression and the accumulation of amylose in response to cool temperatures during seed development in rice. Plant Cell Physiol. 1998, 39, 807–812. [Google Scholar]
- Fan, X.; Li, Y.; Lu, Y.; Zhang, C.; Li, E.; Li, Q.; Tao, K.; Yu, W.; Wang, J.; Chen, Z.; et al. The interaction between amylose and amylopectin synthesis in rice endosperm grown at high temperature. Food Chem. 2019, 301, 125258. [Google Scholar] [CrossRef]
- Hakata, M.; Kuroda, M.; Miyashita, T.; Yamaguchi, T.; Kojima, M.; Sakakibara, H.; Mitsui, T.; Yamakawa, H. Suppression of alpha-amylase genes improves quality of rice grain ripened under high temperature. Plant Biotechnol. J. 2012, 10, 1110–1117. [Google Scholar] [CrossRef] [Green Version]
- Radchuk, V.V.; Borisjuk, L.; Sreenivasulu, N.; Merx, K.; Mock, H.P.; Rolletschek, H.; Wobus, U.; Weschke, W. Spatiotemporal profiling of starch biosynthesis and degradation in the developing barley grain. Plant Physiol. 2009, 150, 190–204. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Li, X.; Zhang, H.; Wang, L.; Zhu, Z.; Gao, J.; Li, C.; Zhu, Y. High temperature inhibits the accumulation of storage materials by inducing alternative splicing of OsbZIP58 during filling stage in rice. Plant Cell Environ. 2020, 43, 1879–1896. [Google Scholar] [CrossRef]
- You, Q.; Huang, J.; Feng, W.; Su, X.; Wang, W.; Yang, M. Study on the method of resistant starch measurement in wheat flour. J. Henan Univ. Technol. 2014, 35, 4. [Google Scholar]
- Štěrbová, L.; Bradová, J.; Sedlá?ek, T.; Holasová, M.; Fiedlerová, V.; Dvořáček, V.; Smrčková, P. Influence of technological processing of wheat grain on starch digestibility and resistant starch content. Starch Stärke 2016, 68, 593–602. [Google Scholar]
- Bao, J.; Xin, Z.; Xu, F.; Qiang, H.; Park, Y.J. Genome-wide association study of the resistant starch content in rice grains: Genome-wide association study of resistant starch in rice. Starch Stärke 2017, 69, 1600343. [Google Scholar]
- Zhang, C.; Zhu, J.; Chen, S.; Fan, X.; Li, Q.; Lu, Y.; Wang, M.; Yu, H.; Yi, C.; Tang, S.; et al. Wx(lv), the Ancestral Allele of Rice Waxy Gene. Mol. Plant 2019, 12, 1157–1166. [Google Scholar] [CrossRef] [Green Version]
- Zeng, D.; Liu, T.; Ma, X.; Wang, B.; Zheng, Z.; Zhang, Y.; Xie, X.; Yang, B.; Zhao, Z.; Zhu, Q.; et al. Quantitative regulation of Waxy expression by CRISPR/Cas9-based promoter and 5’UTR-intron editing improves grain quality in rice. Plant Biotechnol. J. 2020, 18, 2385–2387. [Google Scholar] [CrossRef]
- Zhou, H.; Xia, D.; Zhao, D.; Li, Y.; Li, P.; Wu, B.; Gao, G.; Zhang, Q.; Wang, G.; Xiao, J.; et al. The origin of Wx(la) provides new insights into the improvement of grain quality in rice. J. Integr. Plant Biol. 2021, 63, 878–888. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, Y.; Chen, Z.; Chen, F.; Pan, L.; Lu, Y.; Li, Q.; Fan, X.; Sun, Z.; Liu, Q. Characteristics of Grain Physicochemical Properties and the Starch Structure in Rice Carrying a Mutated ALK/SSIIa Gene. J. Agric. Food Chem. 2020, 68, 13950–13959. [Google Scholar] [CrossRef]
- Chen, Z.; Lu, Y.; Feng, L.; Hao, W.; Li, C.; Yang, Y.; Fan, X.; Li, Q.; Zhang, C.; Liu, Q. Genetic Dissection and Functional Differentiation of ALK(a) and ALK(b), Two Natural Alleles of the ALK/SSIIa Gene, Responding to Low Gelatinization Temperature in Rice. Rice 2020, 13, 39. [Google Scholar] [CrossRef]
- Guo, D.; Ling, X.; Zhou, X.; Li, X.; Wang, J.; Qiu, S.; Yang, Y.; Zhang, B. Evaluation of the Quality of a High-Resistant Starch and Low-Glutelin Rice (Oryza sativa L.) Generated through CRISPR/Cas9-Mediated Targeted Mutagenesis. J. Agric. Food Chem. 2020, 68, 9733–9742. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamamori, M.; Hirano, H.; Hidaka, S.; Nagamine, T. Production of waxy (amylose-free) wheats. Mol. Gen. Genet. 1995, 248, 253–259. [Google Scholar] [CrossRef]
- Shimbata, T.; Nakamura, T.; Vrinten, P.; Saito, M.; Yonemaru, J.; Seto, Y.; Yasuda, H. Mutations in wheat starch synthase II genes and PCR-based selection of a SGP-1 null line. Theor. Appl. Genet. 2005, 111, 1072–1079. [Google Scholar] [CrossRef]
- Saito, M.; Vrinten, P.; Ishikawa, G.; Graybosch, R.; Nakamura, T. A novel codominant marker for selection of the null Wx-B1 allele in wheat breeding programs. Mol. Breed. 2009, 23, 209–217. [Google Scholar]
- Yamamori, M.; Kato, M.; Yui, M.; Kawasaki, M. Resistant starch and starch pasting properties of a starch synthase IIa-deficient wheat with apparent high amylose. Crop Pasture Sci. 2006, 57, 531–535. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Integrated QTL a | Peak SNP | −Log10 P | R2(%) b | Locations c |
|---|---|---|---|---|---|
| 1 | qRS.2AL-1 | AX-110911991 | 3.04–3.06 | 5.56–7.08 | KH, BLUP |
| 2 | qRS.2AL-2 | AX-110371230 | 3.02–3.45 | 5.41–7.75 | KH, SH, BLUP |
| 3 | qRS.2AL-3 | AX-111564575 | 3.09–3.52 | 7.06–8.24 | SH, BLUP |
| 4 | qRS.2BS | AX-95258478 | 3.03–3.75 | 5.71–7.12 | AH, SH |
| 5 | qRS.2DL-1 | AX-111838051 | 3.06–4.03 | 5.56–8.07 | KH, AH, BLUP |
| 6 | qRS.2DL-2 | AX-111141477 | 3.21–3.43 | 6.01–8.00 | KH, AH, BLUP |
| 7 | qRS.3AL | AX-94853365 | 3.05–3.19 | 5.62–5.85 | AH, BLUP |
| 8 | qRS.4AL | AX-94833369 | 3.09–3.69 | 5.63–6.94 | AH, BLUP |
| 9 | qRS.4DS | AX-94546744 | 3.00–3.30 | 5.52–7.82 | SH, BLUP |
| 10 | qRS.5AS | AX-109448373 | 3.21–3.28 | 5.90–7.53 | SH, BLUP |
| 11 | qRS.5AL | AX-108969238 | 3.00–3.03 | 6.95–7.04 | KH, BLUP |
| 12 | qRS.5BL | AX-110545722 | 3.01–4.12 | 5.38–7.93 | KH, SH, BLUP |
| 13 | qRS.6AL | AX-111079685 | 3.03–3.76 | 5.41–8.81 | KH, SH, BLUP |
| 14 | qRS.7BS | AX-111140336 | 3.06–3.41 | 5.48–6.42 | SH, BLUP |
| Code | Peak SNP | SNP | No. of Variety a | RS Content (%) b |
|---|---|---|---|---|
| 1 | AX-110911991 | A/G | 157/49 | 5.09/5.21 * |
| 2 | AX-110371230 | A/G | 41/159 | 5.22/5.11 * |
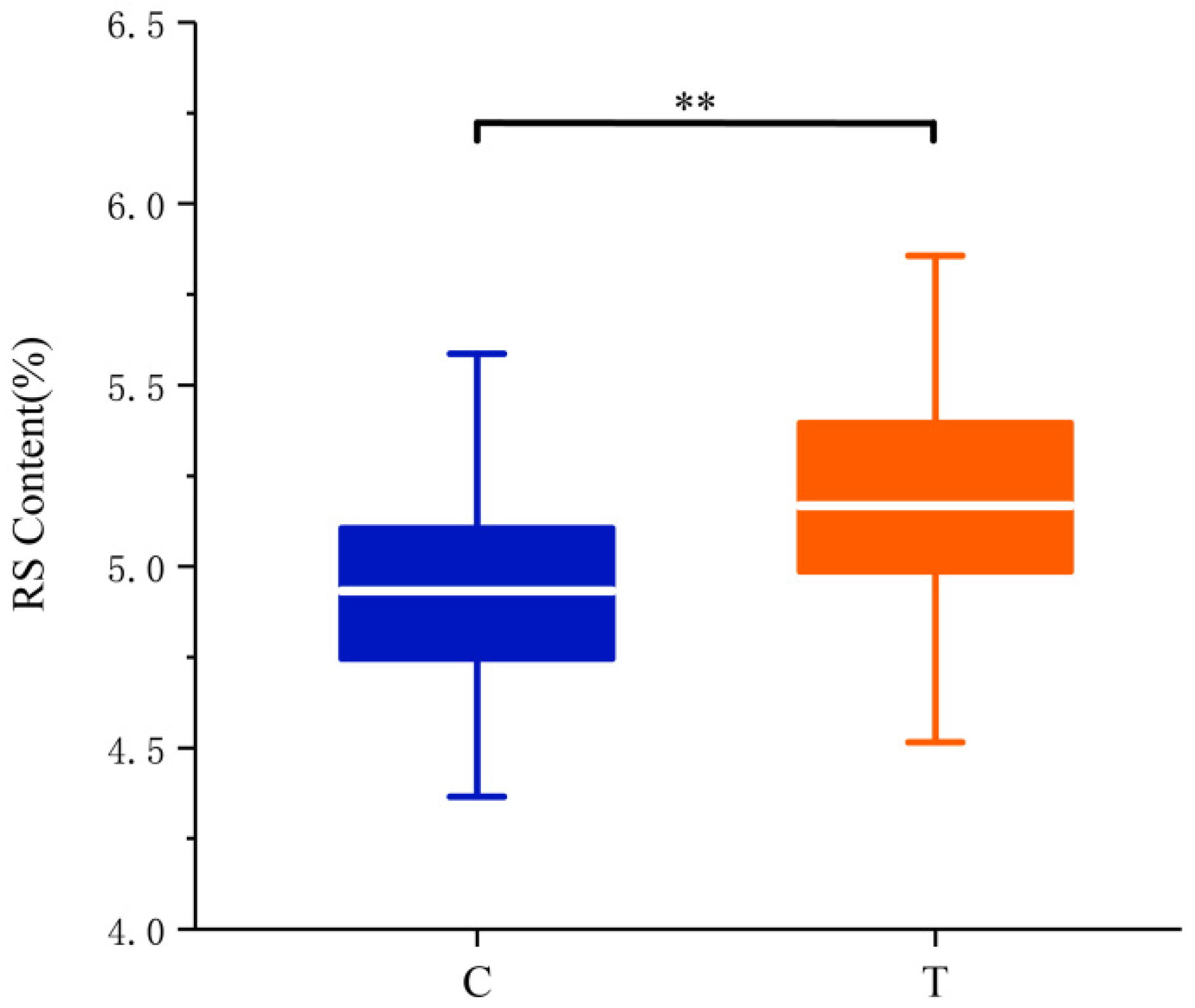
| 3 | AX-111564575 | C/T | 36/162 | 4.99/5.17 ** |
| 4 | AX-95258478 | A/R | 182/25 | 5.11/5.27 * |
| 5 | AX-111838051 | C/G | 80/125 | 5.18/5.08 * |
| 6 | AX-111141477 | A/G | 80/125 | 5.18/5.08 * |
| 7 | AX-94853365 | C/S | 167/39 | 5.16/4.99 ** |
| 8 | AX-94833369 | C/Y | 179/28 | 5.16/4.89 *** |
| 9 | AX-94546744 | C/T/Y | 26/25/152 | 4.88/5.18/5.16 (a/b/b) |
| 10 | AX-109448373 | A/G/R | 43/59/102 | 5.02/5.15/5.16 (a/b/b) |
| 11 | AX-108969238 | C/T | 149/55 | 5.16/5.05 * |
| 12 | AX-110545722 | A/G | 109/98 | 5.17/5.07 * |
| 13 | AX-111079685 | A/G | 138/65 | 5.17/5.05 ** |
| 14 | AX-111140336 | C/T | 146/59 | 5.07/5.27 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, J.; Deng, H.; Wang, Y.; Liu, C.; Geng, S.; Li, W.; Qin, M.; Dai, Z.; Shi, X.; Yang, P.; et al. Starch Bio-Synthetic Pathway Genes Contribute to Resistant Starch Content Differentiation in Bread Wheat. Agronomy 2022, 12, 2967. https://doi.org/10.3390/agronomy12122967
Hou J, Deng H, Wang Y, Liu C, Geng S, Li W, Qin M, Dai Z, Shi X, Yang P, et al. Starch Bio-Synthetic Pathway Genes Contribute to Resistant Starch Content Differentiation in Bread Wheat. Agronomy. 2022; 12(12):2967. https://doi.org/10.3390/agronomy12122967
Chicago/Turabian StyleHou, Jinna, Hui Deng, Yingdong Wang, Congcong Liu, Shenghui Geng, Wenxu Li, Maomao Qin, Ziju Dai, Xia Shi, Pan Yang, and et al. 2022. "Starch Bio-Synthetic Pathway Genes Contribute to Resistant Starch Content Differentiation in Bread Wheat" Agronomy 12, no. 12: 2967. https://doi.org/10.3390/agronomy12122967
APA StyleHou, J., Deng, H., Wang, Y., Liu, C., Geng, S., Li, W., Qin, M., Dai, Z., Shi, X., Yang, P., Tian, B., Yao, W., Wu, Z., Lei, Z., & Zhou, Z. (2022). Starch Bio-Synthetic Pathway Genes Contribute to Resistant Starch Content Differentiation in Bread Wheat. Agronomy, 12(12), 2967. https://doi.org/10.3390/agronomy12122967