
Light Interception and Radiation Use Efficiency of Three Cassava Genotypes with Different Plant Types and Seasonal Variations

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Plant Materials
2.2. Data Collections
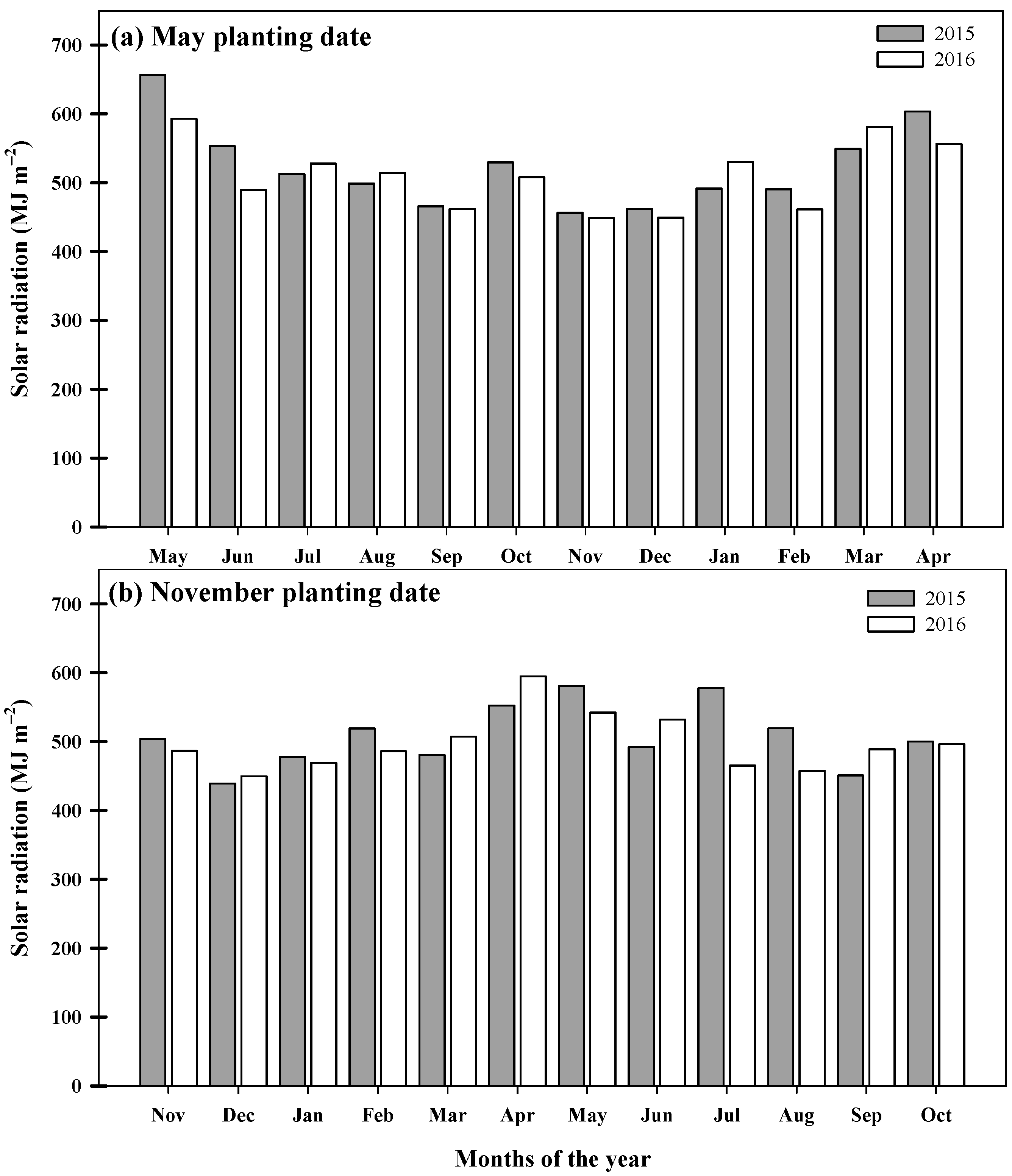
2.2.1. Weather Conditions
2.2.2. Soil Physical and Chemical Properties
2.2.3. Light Interception and Cumulative Solar Radiation
2.2.4. Leaf Area Index (LAI) and Extinction Coefficient (k)
2.2.5. Total Biomass and Yield
2.2.6. Radiation Use Efficiency (RUE)
2.3. Statistical Analysis
3. Results
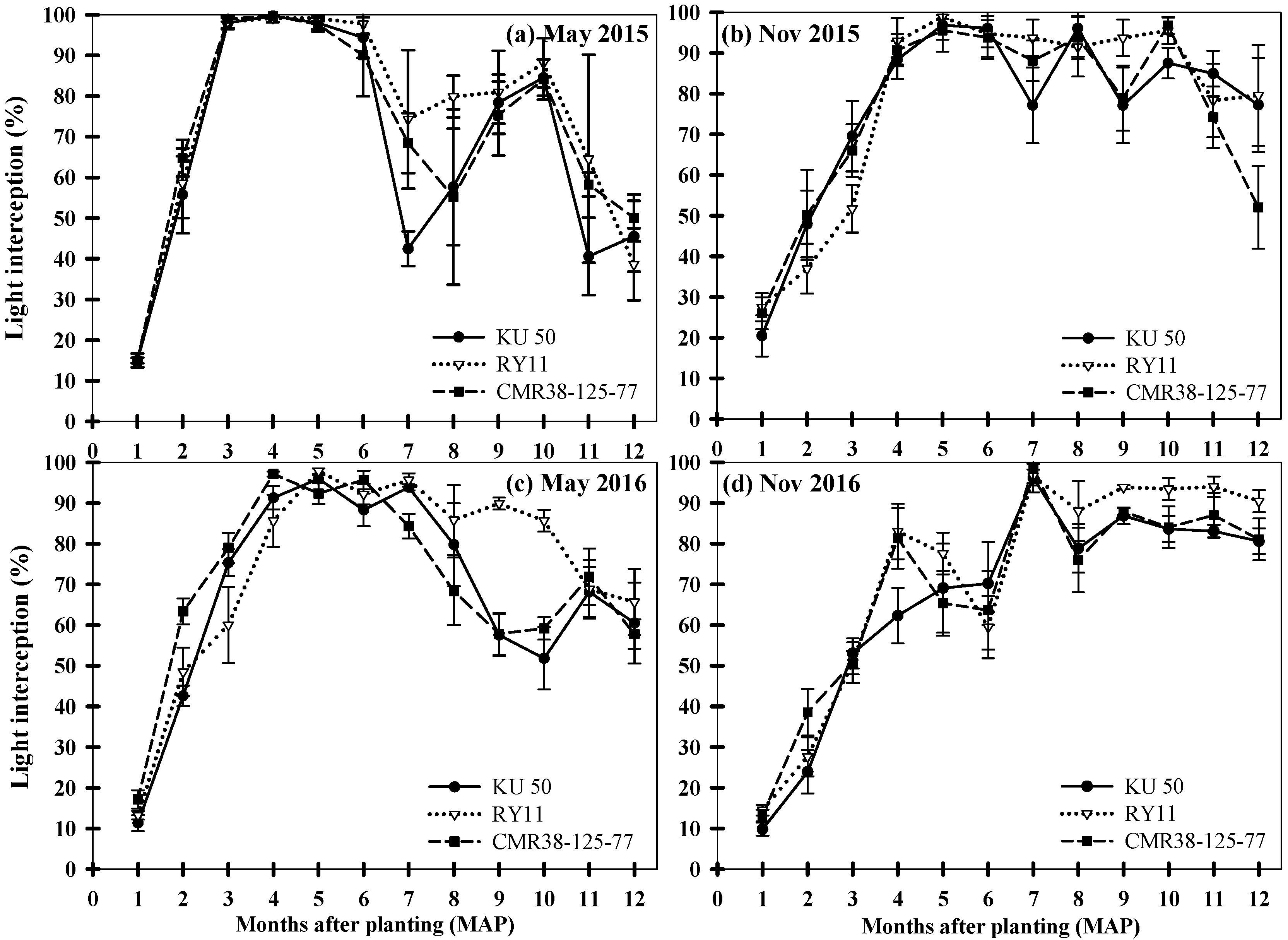
3.1. Light Interception

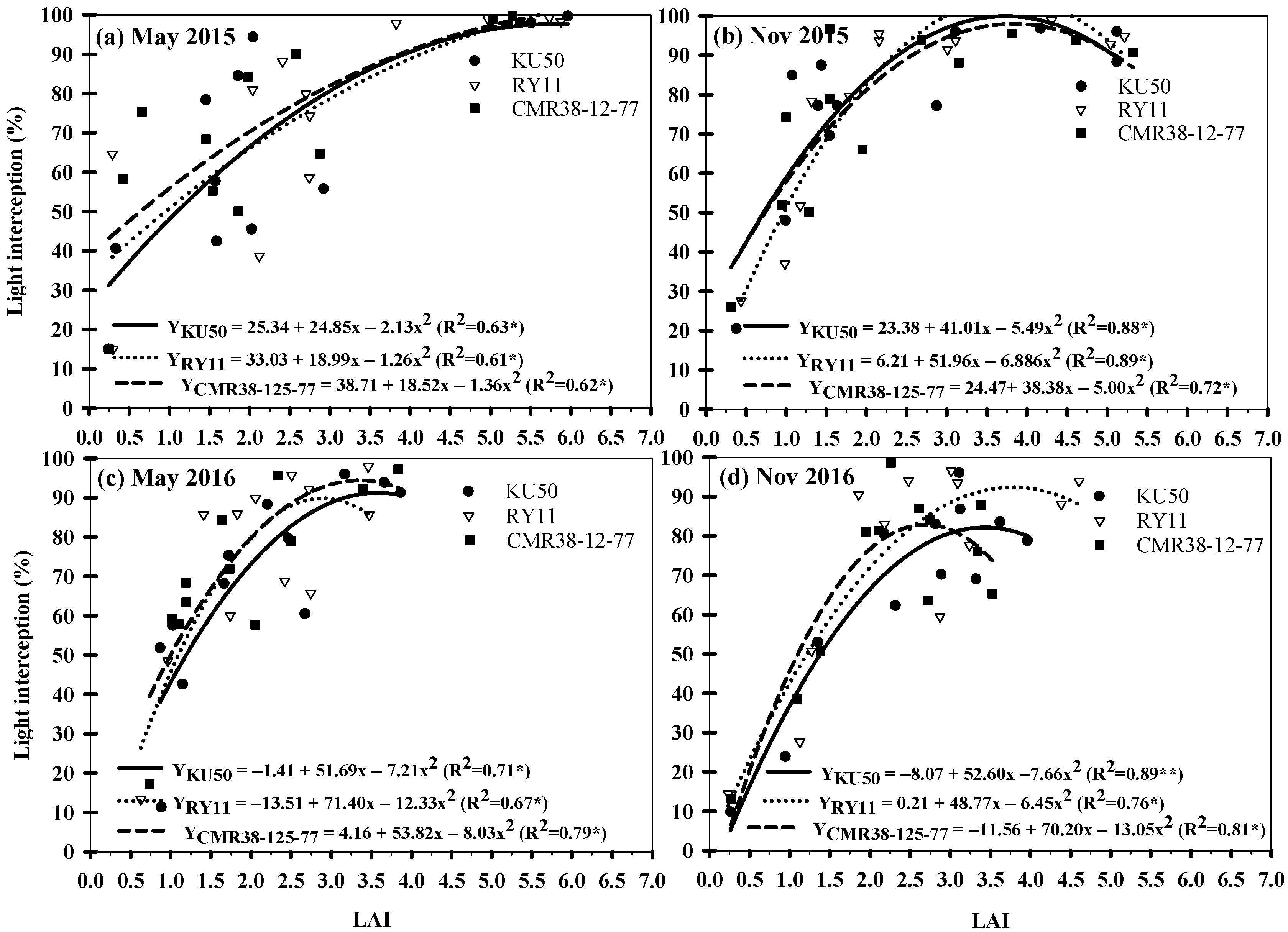
3.2. Light Interception and LAI

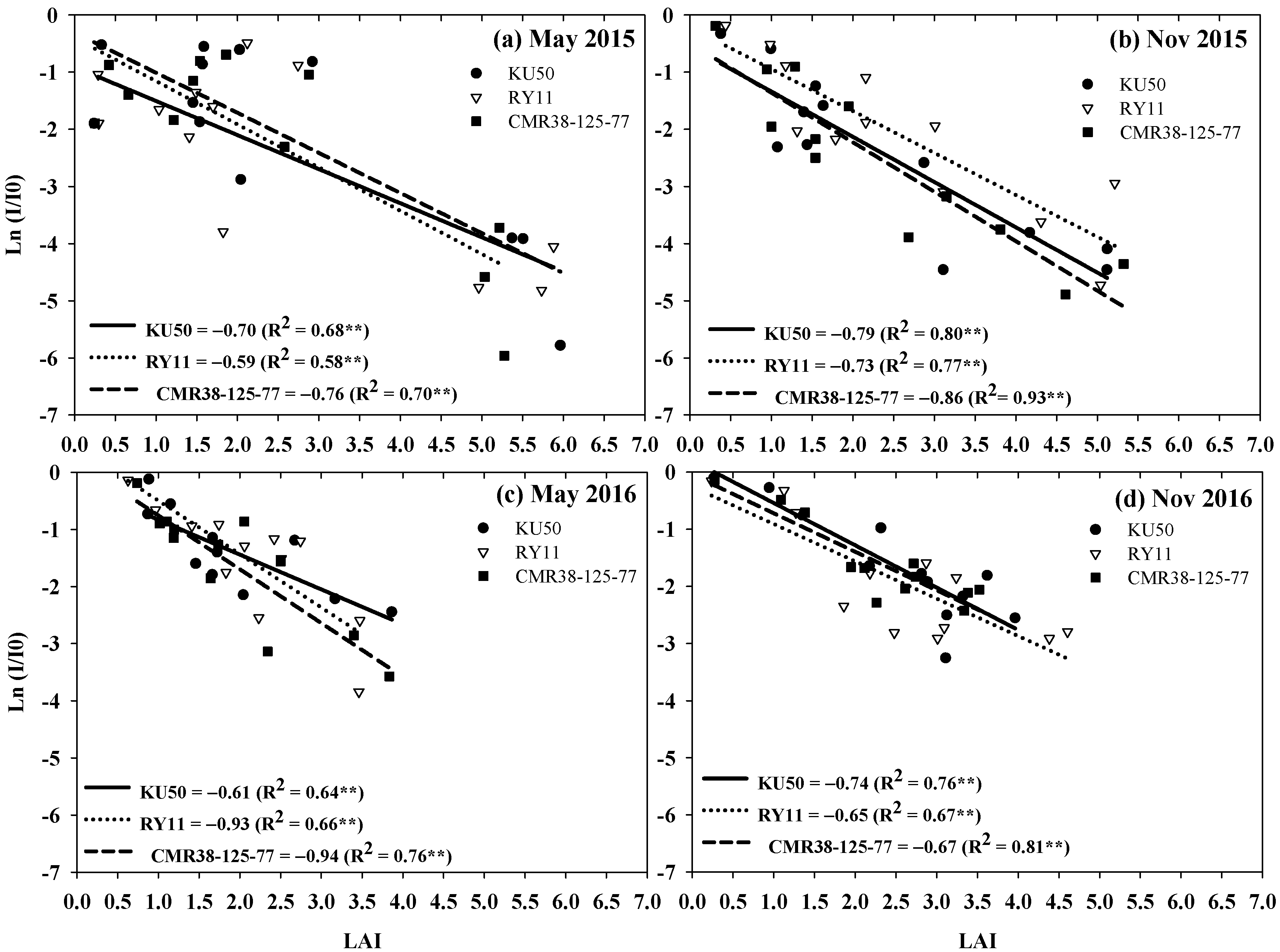
3.3. Extinction Coefficient (k)
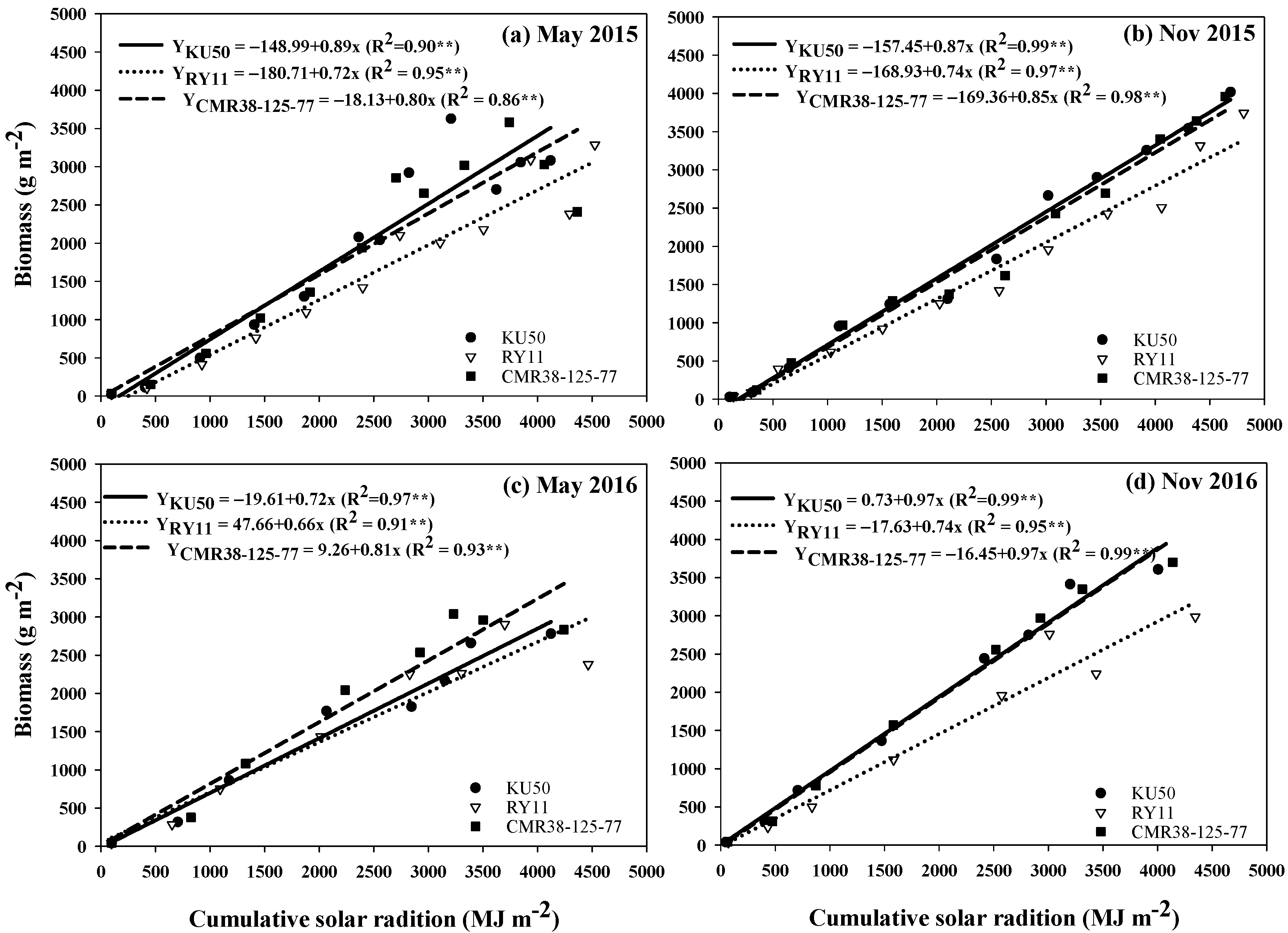
3.4. Cumulative Solar Radiation Interception

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes/Plant Age | 0–3 MAP | 4–6 MAP | 7–9 MAP | 10–12 MAP | F-Test | Total | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| May-15 | ||||||||||
| Kasetsart 50 | 910 | b | 1454 | a | 846 | b | 913 | b | ** | 4121 |
| Rayong 11 | 927 | b | 1474 | a | 1105 | b | 1021 | b | ** | 4526 |
| CMR38-125-77 | 964 | b | 1429 | a | 938 | b | 1035 | b | ** | 4365 |
| F-test | ns | ns | ns | ns | ns | |||||
| Nov-15 | ||||||||||
| Kasetsart 50 | 646 | c | 1455 | a | 1367 | a | 1224 | b | ** | 4691 |
| Rayong 11 | 548 | c | 1480 | a | 1537 | a | 1247 | b | ** | 4811 |
| CMR38-125-77 | 667 | c | 1448 | a | 1430 | a | 1098 | b | ** | 4642 |
| F-test | ns | ns | ns | ns | ns | |||||
| May-16 | ||||||||||
| Kasetsart 50 | 674 | B c | 1361 | a | 1084 | B b | 972 | b | ** | 4091 |
| Rayong 11 | 633 | B c | 1361 | a | 1292 | A ab | 1160 | b | ** | 4445 |
| CMR38-125-77 | 829 | A c | 1412 | a | 992 | B cb | 1012 | b | ** | 4245 |
| F-test | * | ns | ** | ns | ns | |||||
| Nov-16 | ||||||||||
| Kasetsart 50 | 404 | c | 1071 | b | 1345 | a | 1189 | B b | ** | 4008 |
| Rayong 11 | 433 | c | 1151 | b | 1429 | a | 1336 | A a | ** | 4347 |
| CMR38-125-77 | 476 | c | 1105 | b | 1348 | a | 1212 | B ab | ** | 4140 |
| F-test | ns | ns | ns | * | ns | |||||
3.5. Biomass and Cumulative Solar Radiation

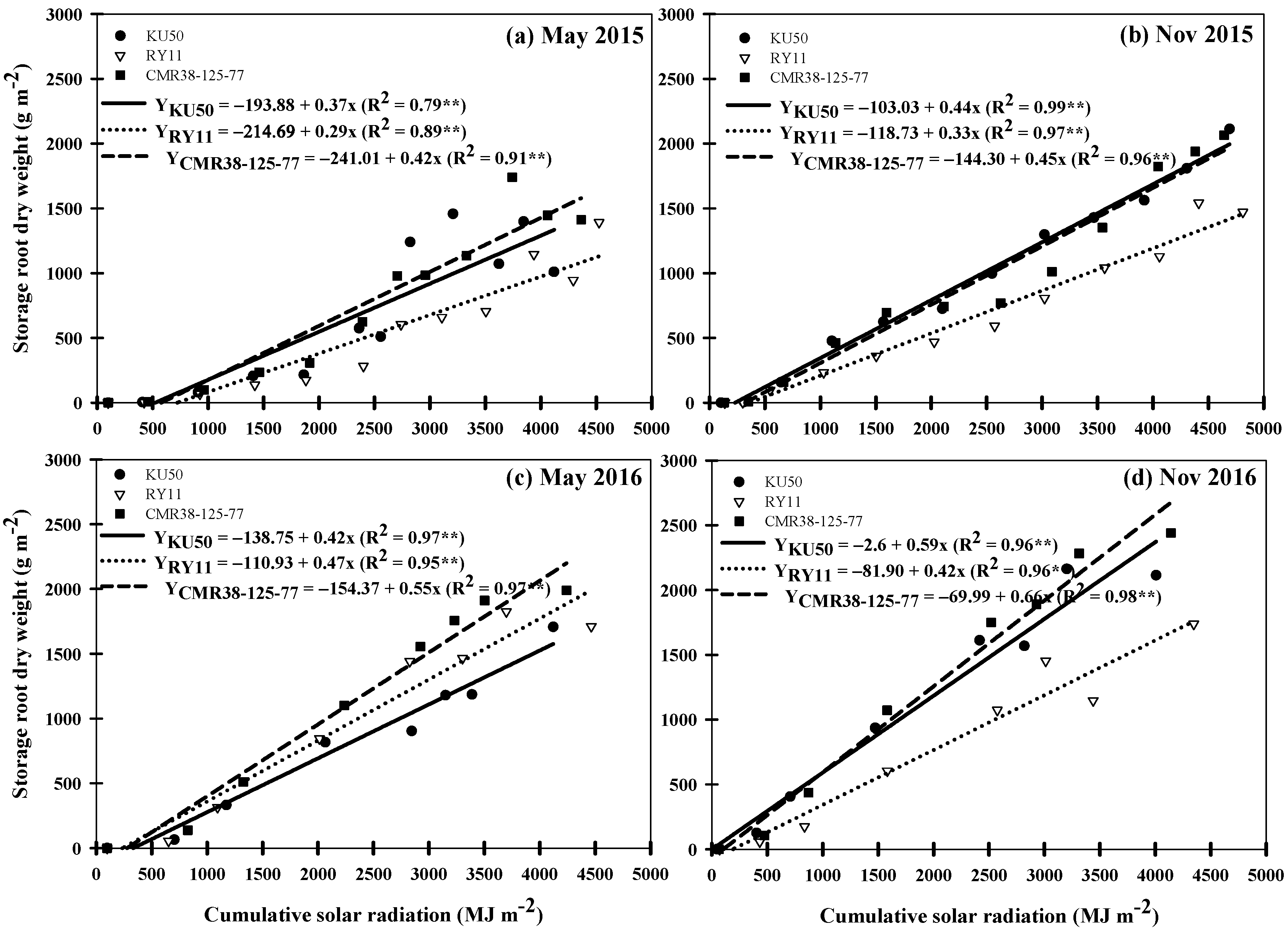
3.6. Storage Root Dry Weight and Cumulative Solar Radiation
3.7. Radiation Use Efficiency for Biomass (RUEbi)
| Genotypes/Plant Age | 0–3 MAP | 4–6 MAP | 7–9 MAP | 10–12 MAP | F-Test | ||||
|---|---|---|---|---|---|---|---|---|---|
| May-15 | |||||||||
| Kasetsart 50 | 0.55 | bc | 1.09 | A ab | 1.31 | a | 0.44 | c | * |
| Rayong 11 | 0.45 | 0.68 | B | 0.68 | 1.18 | ns | |||
| CMR38-125-77 | 0.58 | 0.97 | A | 0.49 | 0.59 | ns | |||
| F-test | ns | * | ns | ns | |||||
| Nov-15 | |||||||||
| Kasetsart 50 | 0.63 | c | 0.62 | c | 1.17 | A a | 0.92 | b | ** |
| Rayong 11 | 0.72 | b | 0.58 | b | 0.77 | B b | 1.06 | a | ** |
| CMR38-125-77 | 0.72 | c | 0.62 | c | 0.93 | AB b | 1.16 | a | ** |
| F-test | ns | ns | * | ns | |||||
| May-16 | |||||||||
| Kasetsart 50 | 0.49 | 0.95 | 0.48 | 0.85 | ns | ||||
| Rayong 11 | 0.47 | 0.74 | 0.70 | 0.37 | ns | ||||
| CMR38-125-77 | 0.46 | b | 1.16 | a | 0.77 | ab | 0.32 | b | * |
| F-test | ns | ns | ns | ns | |||||
| Nov-16 | |||||||||
| Kasetsart 50 | 0.75 | 1.01 | 1.02 | 0.73 | ns | ||||
| Rayong 11 | 0.55 | bc | 0.77 | b | 1.16 | a | 0.28 | c | ** |
| CMR38-125-77 | 0.61 | b | 1.19 | a | 1.05 | a | 0.59 | b | ** |
| F-test | ns | ns | ns | ns | |||||
3.8. Radiation Use Efficiency for Storage Root Dry Weight (RUEsr)
| Genotypes/Plant Age | 0–3 MAP | 4–6 MAP | 7–9 MAP | 10–12 MAP | F-Test | ||||
|---|---|---|---|---|---|---|---|---|---|
| May-15 | |||||||||
| Kasetsart 50 | 0.09 | c | 0.34 | AB b | 0.94 | A a | 0.48 | b | ** |
| Rayong 11 | 0.08 | b | 0.15 | B b | 0.38 | B b | 0.71 | a | * |
| CMR38-125-77 | 0.11 | c | 0.37 | A ab | 0.56 | B a | 0.27 | bc | ** |
| F-test | ns | ** | ** | ns | |||||
| Nov-15 | |||||||||
| Kasetsart 50 | 0.25 | b | 0.39 | A ab | 0.52 | a | 0.56 | a | * |
| Rayong 11 | 0.18 | c | 0.25 | B bc | 0.38 | a | 0.35 | ab | * |
| CMR38-125-77 | 0.24 | c | 0.41 | A bc | 0.43 | b | 0.66 | a | ** |
| F-test | ns | * | ns | ns | |||||
| May-16 | |||||||||
| Kasetsart 50 | 0.10 | c | 0.56 | a | 0.33 | b | 0.55 | A a | * |
| Rayong 11 | 0.09 | 0.55 | 0.52 | 0.21 | B | ns | |||
| CMR38-125-77 | 0.17 | 0.69 | 0.67 | 0.23 | B | ns | |||
| F-test | ns | ns | ns | * | |||||
| Nov-16 | |||||||||
| Kasetsart 50 | 0.33 | 0.76 | A | 0.46 | 0.47 | ns | |||
| Rayong 11 | 0.14 | b | 0.47 | B ab | 0.61 | a | 0.21 | b | * |
| CMR38-125-77 | 0.22 | c | 0.90 | A a | 0.62 | b | 0.44 | bc | ** |
| F-test | ns | * | ns | ns | |||||
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Falade, K.O.; Akingbala, J.O. Utilization of cassava for food. Food Rev. Int. 2010, 27, 51–83. [Google Scholar] [CrossRef]
- El-Sharkawy, M.A. Cassava biology and physiology. Plant Mol. Biol. 2004, 56, 481–501. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.I.; Kongsil, P.; Nguyễn, V.A.; Ou, W.; Sholihin-Srean, P.; Sheela, M.N.; López-Lavalle, L.A.B.; Utsumi, Y.; Lu, C.; Kittipadakul, P.; et al. Cassava breeding and agronomy in Asia: 50 years of history and future directions. Breed. Sci. 2020, 70, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Office of Agricultural Economics. Agricultural Production Index. Available online: http://www.oae.go.th/view/1/Home/EN-US (accessed on 23 January 2019).
- De Vries, C.A.; Ferwerda, J.D.; Flach, M. Choice of food crop in relation to actual and potential production in the tropics. Neth. J. Agric. Sci. 1967, 19, 241–248. [Google Scholar] [CrossRef]
- El-Sharkawy, M.A. Drought-tolerant cassava for Africa, Asia, and Latin-America. BioScience 1993, 43, 441–451. [Google Scholar] [CrossRef]
- Phoncharoen, P.; Banterng, P.; Vorasoot, N.; Jogloy, S.; Theerakulpisut, P.; Hoogenboom, G. Growth rates and yields of cassava at different planting dates in a tropical savanna climate. Sci. Agric. 2019, 76, 376–388. [Google Scholar] [CrossRef]
- Monteith, J.L. Validity of the correlation between intercepted radiation and biomass. Agric. For. Meteorol. 1994, 68, 213–220. [Google Scholar] [CrossRef]
- Pellet, D.; El-Sharkawy, M.A. Cassava varietal response to fertilization: Growth dynamic and implications for cropping sustainability. Exp. Agric. 1997, 33, 353–365. [Google Scholar] [CrossRef]
- Gonias, E.D.; Oosterhuis, D.M.; Bibi, A.C.; Roberts, B.A. Radiation use efficiency of cotton in contrasting environments. Am. J. Plant Sci. 2012, 3, 649–654. [Google Scholar] [CrossRef]
- Raghuveera Rao, P.; Sailaja, B.; Desiraju, S.; Nageswara Rao, D.; Voleti, S.R.; Satyanarayana Reddy, P.; Sudarsana Rao, G.V. Variable radiation use efficiency in rice cultures grown at different locations. Indian J. Plant Physiol. 2012, 17, 57–60. [Google Scholar]
- Liu, X.; Rahman, T.; Yang, F.; Song, C.; Yong, T.; Liu, J.; Zhang, C.; Yang, W. PAR interception and utilization in different maize and soybean intercropping patterns. PLoS ONE 2017, 12, e0169218. [Google Scholar] [CrossRef]
- Monteith, J.L.; Moss, C.J. Climate and the efficiency of crop production in Britain. Philos. Trans. R. Soc. 1977, 281, 277–294. [Google Scholar] [CrossRef]
- Long, S.P.; Zhu, X.G.; Naidu, S.L.; Ort, D.R. Can improvement in photosynthesis increase crop yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef]
- Niinemets, U. Photosynthesis and resource distribution through plant canopies. Plant Cell Environ. 2007, 30, 1052–1071. [Google Scholar] [CrossRef]
- Reynolds, M.; Bonnett, D.; Chapman, S.C.; Furbank, R.T.; Manes, Y.; Mather, D.E.; Parry, M.A. Raising yield potential of wheat. I. Overview of a consortium approach and breeding strategies. J. Exp. Bot. 2011, 62, 439–452. [Google Scholar] [CrossRef]
- Wolf, D.D.; Carson, E.; Brown, R.H. Light interception efficiency measurements. J. Agron. Educ. 1972, 1, 40–42. [Google Scholar] [CrossRef]
- Vos, J.; Evers, J.B.; Buck-Sorlin, G.H.; Andrieu, B.; Chelle, M.; de Visser, P.H.B. Functional–structural plant modelling: A new versatile tool in crop science. J. Exp. Bot. 2010, 61, 2101–2115. [Google Scholar] [CrossRef]
- Tumuhimbise, R.; Melis, R.; Shanahan, P.; Kawuki, R. Genotype × environment interaction effects on early fresh storage root yield and related traits in cassava. Crop J. 2014, 2, 329–337. [Google Scholar] [CrossRef]
- Lahai, T.; Ekanayake, I.J.; Koroma, J.P.C. Influence of canopy structure on yield of cassava cultivars at various toposequences of an inland valley agro ecosystem. J. Agric. Biotechnol. Sustain. Dev. 2013, 5, 36–47. [Google Scholar] [CrossRef]
- Werner, C.; Ryel, R.J.; Correia, O.; Beyschlag, W. Structural and functional variability within the canopy and its relevance for carbon gain and stress avoidance. Acta Oecologica 2001, 22, 129–138. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, X.B.; Chen, Y.H. Effects of irrigation and precision planting patterns on photosynthetic product of wheat. Agron. J. 2016, 108, 2322–2328. [Google Scholar] [CrossRef]
- De Souza, A.P.; Massenburg, L.N.; Jaiswal, D.; Cheng, S.; Shekar, R.; Long, S.P. Rooting for cassava: Insights into photosynthesis and associated physiology as a route to improve yield potential. New Phytol. 2016, 213, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Hughes, G.; Keatinge, J.D.H.; Cooper, P.J.M.; Dee, N.F. Solar radiation interception and utilization by chickpea (Cicer arietinum L.) crops in northern Syria. J. Agric. Sci. 1987, 108, 419–424. [Google Scholar] [CrossRef]
- Hunter, R.B. Increased leaf area (source) and yield of maize in short-season areas. Crop Sci. 1980, 20, 571–574. [Google Scholar] [CrossRef]
- Monsi, M.; Saeki, T. On the factor light in plant communities and its importance for matter production. Ann. Bot. 2005, 95, 549–567. [Google Scholar] [CrossRef]
- Howeler, R.H. Cassava mineral nutrition and fertilization. In Cassava: Biology, Production and Utilization; Hillocks, R.J., Thresh, J.M., Bellotti, A.C., Eds.; CABI Publishing: New York, NY, USA, 2002; pp. 115–147. [Google Scholar]
- Monteith, J.L. Solar radiation and productivity in tropical ecosystems. J. Appl. Ecol. 1972, 9, 747–766. [Google Scholar] [CrossRef]
- LI-COR Inc. LAI-2000 Plant Canopy Analyzer Instruction/Operating Manual; LI-COR Inc.: Lincoln, NE, USA, 1992. [Google Scholar]
- LI-COR Inc. LI-191SA Line Quantum Sensor Brochure; LI-COR Inc.: Lincoln, NE, USA, 2015. [Google Scholar]
- Cooper, J.P. Photosynthetic efficiency of the whole plant. In Food Production and Consumption: The Efficiency of Human Food Chains and Nutrient Cycles; Duckham, A.N., Jones, J.G.W., Roberts, E.H., Eds.; North-Holland Publishing Company: Amsterdam, The Netherlands, 1976; pp. 107–126. [Google Scholar]
- Hay, R.; Walker, A. An Introduction to the Physiology of Crop Yield; Longmans: London, UK, 1989; p. 292. [Google Scholar]
- Alves, A.A.C. Cassava botany and physiology. In Cassava: Biology, Production and Utilization; Hillocks, R.J., Thresh, J.M., Bellotti, A.C., Eds.; CABI Publishing: New York, NY, USA, 2002; pp. 67–89. [Google Scholar]
- Statistix, Version 10; Analytical Software: Tallahassee, FL, USA, 2013.
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons: New York, NY, USA, 1984; pp. 1–628. [Google Scholar]
- Mahakosee, S.; Jogloy, S.; Vorasoot, N.; Theerakulpisut, P.; Banterng, P.; Kesmala, T.; Holbrook, C.C.; Kvien, C. Seasonal variations in canopy size and yield of Rayong 9 cassava genotype under rainfed and irrigated conditions. Agronomy 2019, 9, 362. [Google Scholar] [CrossRef]
- Irikura, V.; Cock, J.H.; Kawano, K. The physiological basis of genotype-temperature interactions in cassava. Field Crops Res. 1979, 2, 227–239. [Google Scholar] [CrossRef]
- Keating, B.A.; Evenson, J.P.; Fukai, S. Environmental effects on growth and developments of cassava (Manihot esculenta Crantz.). I. Crop development. Field Crops Res. 1982, 5, 271–281. [Google Scholar] [CrossRef]
- Fukai, S.; Alcoy, A.B.; Llamelo, A.B.; Patterson, R.D. Effects of solar radiation on growth of cassava (Manihot esculenta Crantz.). I. Canopy development and dry matter growth. Field Crops Res. 1984, 9, 347–360. [Google Scholar] [CrossRef]
- Veltkamp, H.J. Physiological Causes of Yield Variation in Cassava (Manihot Esculenta Crantz). Ph.D. Thesis, University of Wageningen, Wageningen, The Netherlands, 1985; p. 132. [Google Scholar]
- El-Sharkawy, M.A.; De Tafur, S.M.; Cadavid, L.F. Potential photosynthesis of cassava as affected by growth conditions. Crop Sci. 1992, 32, 1336–1342. [Google Scholar] [CrossRef]
- Cock, J.H.; Franklin, D.; Sandoval, G.; Juri, P. The ideal cassava plant for maximum yield. Crop Sci. 1979, 19, 271–279. [Google Scholar] [CrossRef]
- Boote, K.J.; Loomis, R.S. (Eds.) The prediction of canopy assimilation. In Modeling Crop Photosynthesis—From Biochemistry to Canopy; No. 19; CSSA: Fitchburg, WI, USA, 1991; pp. 109–137. [Google Scholar]
- Pradhan, S.; Sehgal, V.K.; Bandyopadhyay, K.K.; Panigrahi, P.; Parihar, C.M.; Jat, S.L. Radiation interception, extinction coefficient and use efficiency of wheat crop at various irrigation and nitrogen levels in a semi-arid location. Indian J. Plant Physiol. 2018, 23, 416–425. [Google Scholar] [CrossRef]
- O’Connell, M.G.; O’Leary, G.J.; Whitfield, D.M.; Connor, D.J. Interception of photosynthetically active radiation and radiation-use efficiency of wheat, field pea and mustard in a semi-arid environment. Field Crops Res. 2004, 85, 111–124. [Google Scholar] [CrossRef]
- El-Sharkawy, M.A.; Cock, J.H. Water use efficiency of cassava. I. Effects of air humidity and water stress on stomatal conductance and gas exchange. Crop Sci. 1984, 24, 497–502. [Google Scholar] [CrossRef]
- Mahakosee, S.; Jogloy, S.; Vorasoot, N.; Theerakulpisut, P.; Banterng, P.; Holbrook, C.C.; Kvien, C. Seasonal variation in canopy size, light penetration and photosynthesis of three cassava genotypes with different canopy architectures. Agronomy 2020, 10, 1554. [Google Scholar] [CrossRef]
- Phoncharoen, P.; Banterng, P.; Vorasoot, N.; Jogloy, S.; Theerakulpisut, P.; Hoogenboom, G. The impact of seasonal environments in a tropical savanna climate on forking, leaf area index, and biomass of cassava genotypes. Agronomy 2019, 9, 19. [Google Scholar] [CrossRef]
- El-Sharkawy, M.A.; Cadavid, L.F. Response of cassava to prolonged water stress imposed at different stages of growth. Exp. Agric. 2002, 38, 333–350. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahakosee, S.; Jogloy, S.; Vorasoot, N.; Theerakulpisut, P.; Toomsan, B.; Holbrook, C.C.; Kvien, C.K.; Banterng, P. Light Interception and Radiation Use Efficiency of Three Cassava Genotypes with Different Plant Types and Seasonal Variations. Agronomy 2022, 12, 2888. https://doi.org/10.3390/agronomy12112888
Mahakosee S, Jogloy S, Vorasoot N, Theerakulpisut P, Toomsan B, Holbrook CC, Kvien CK, Banterng P. Light Interception and Radiation Use Efficiency of Three Cassava Genotypes with Different Plant Types and Seasonal Variations. Agronomy. 2022; 12(11):2888. https://doi.org/10.3390/agronomy12112888
Chicago/Turabian StyleMahakosee, Supattra, Sanun Jogloy, Nimitr Vorasoot, Piyada Theerakulpisut, Banyong Toomsan, Carl Corley Holbrook, Craig K. Kvien, and Poramate Banterng. 2022. "Light Interception and Radiation Use Efficiency of Three Cassava Genotypes with Different Plant Types and Seasonal Variations" Agronomy 12, no. 11: 2888. https://doi.org/10.3390/agronomy12112888
APA StyleMahakosee, S., Jogloy, S., Vorasoot, N., Theerakulpisut, P., Toomsan, B., Holbrook, C. C., Kvien, C. K., & Banterng, P. (2022). Light Interception and Radiation Use Efficiency of Three Cassava Genotypes with Different Plant Types and Seasonal Variations. Agronomy, 12(11), 2888. https://doi.org/10.3390/agronomy12112888