European Grapevine Moth and Vitis vinifera L. Phenology in the Douro Region: (A)synchrony and Climate Scenarios



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
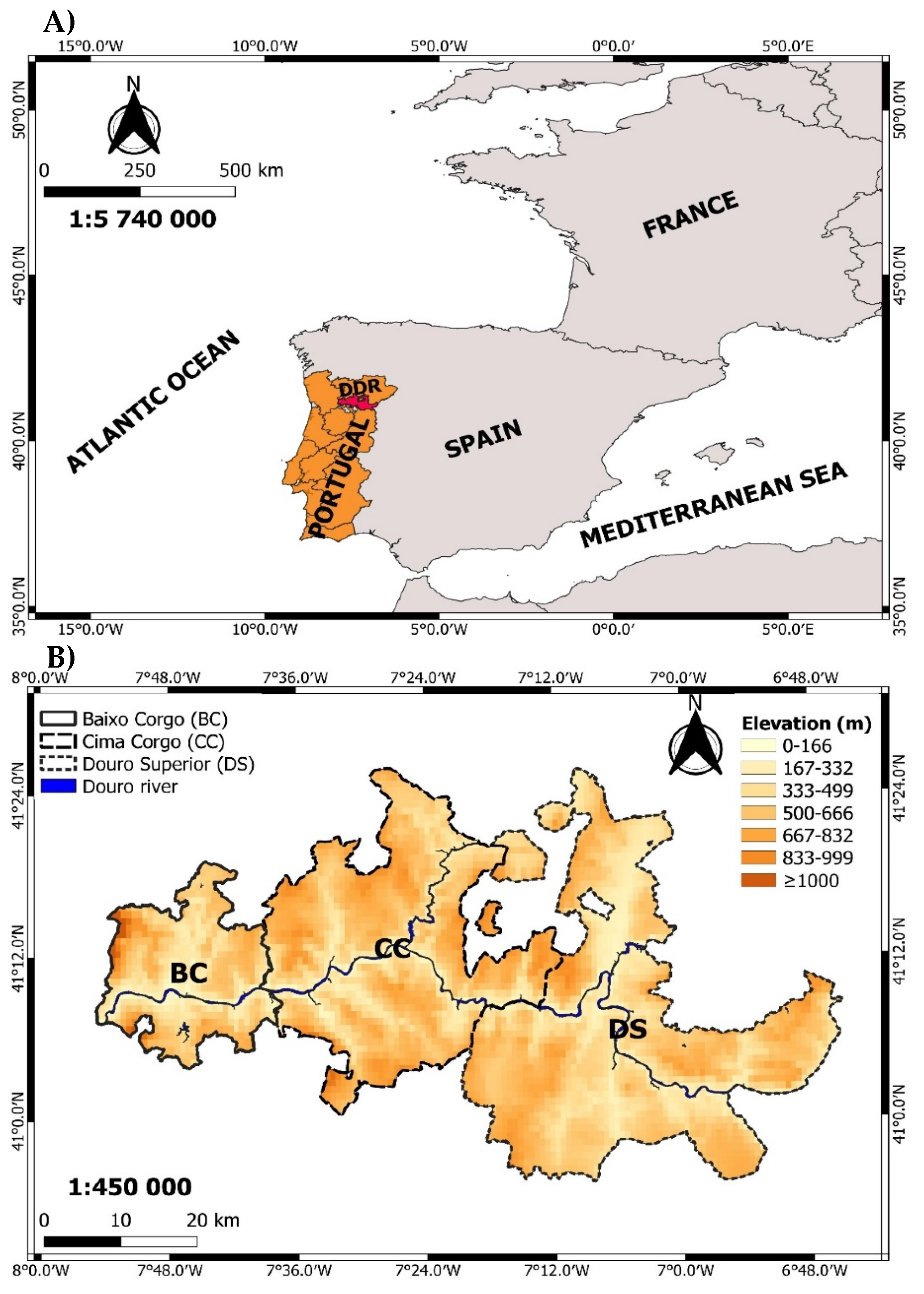
2.1. Study Area and Climate Conditions
2.2. Phenological Models
2.2.1. Grapevine
2.2.2. LB Flight
2.2.3. Robustness and Predictability
2.3. Climate Dataset for the Historic Period and Future Scenarios
2.4. Impact of Future Climate Scenarios
2.5. Interaction of Trophic Levels
3. Results
3.1. Grapevine Phenology
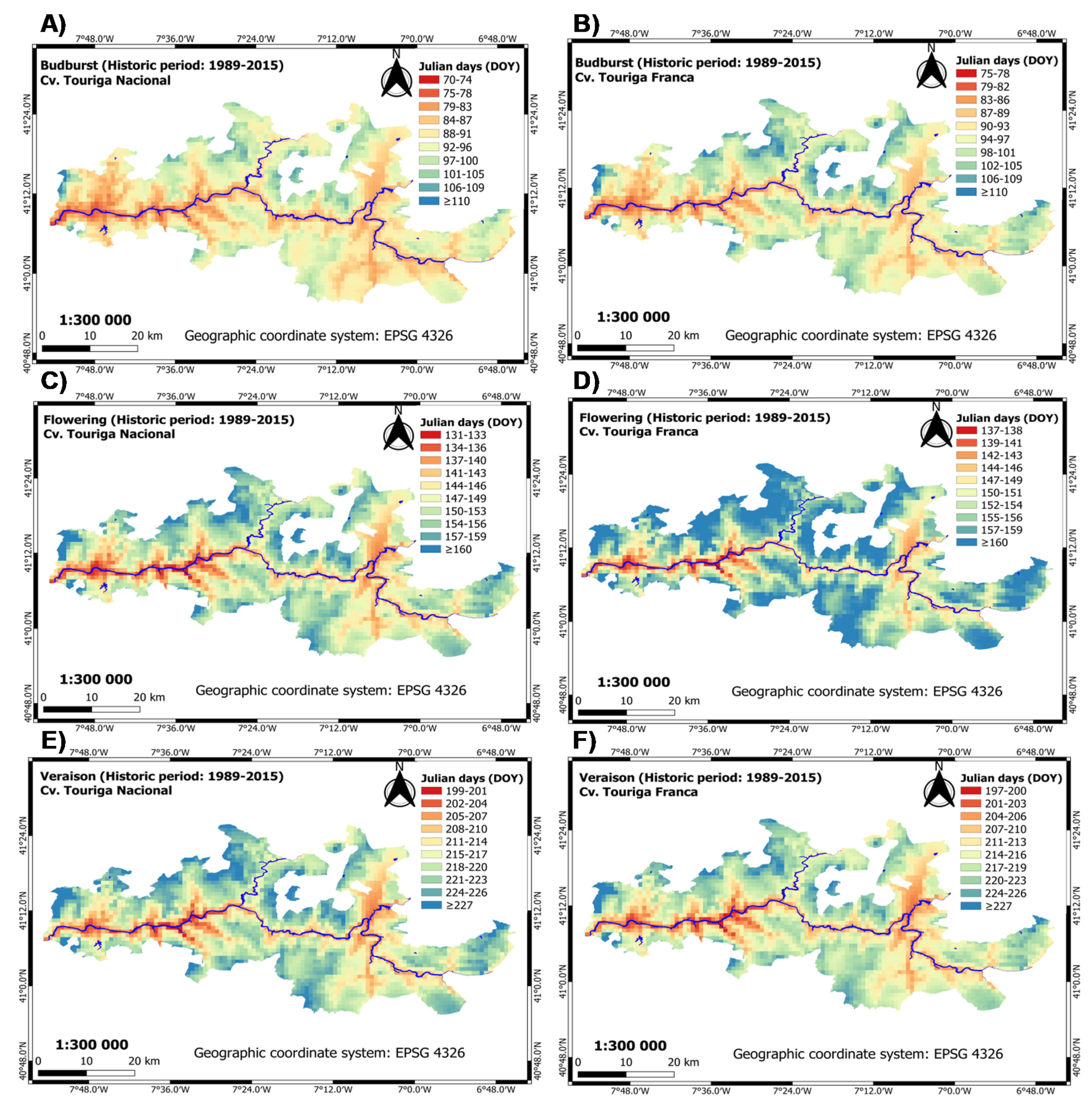
3.1.1. Historic Period
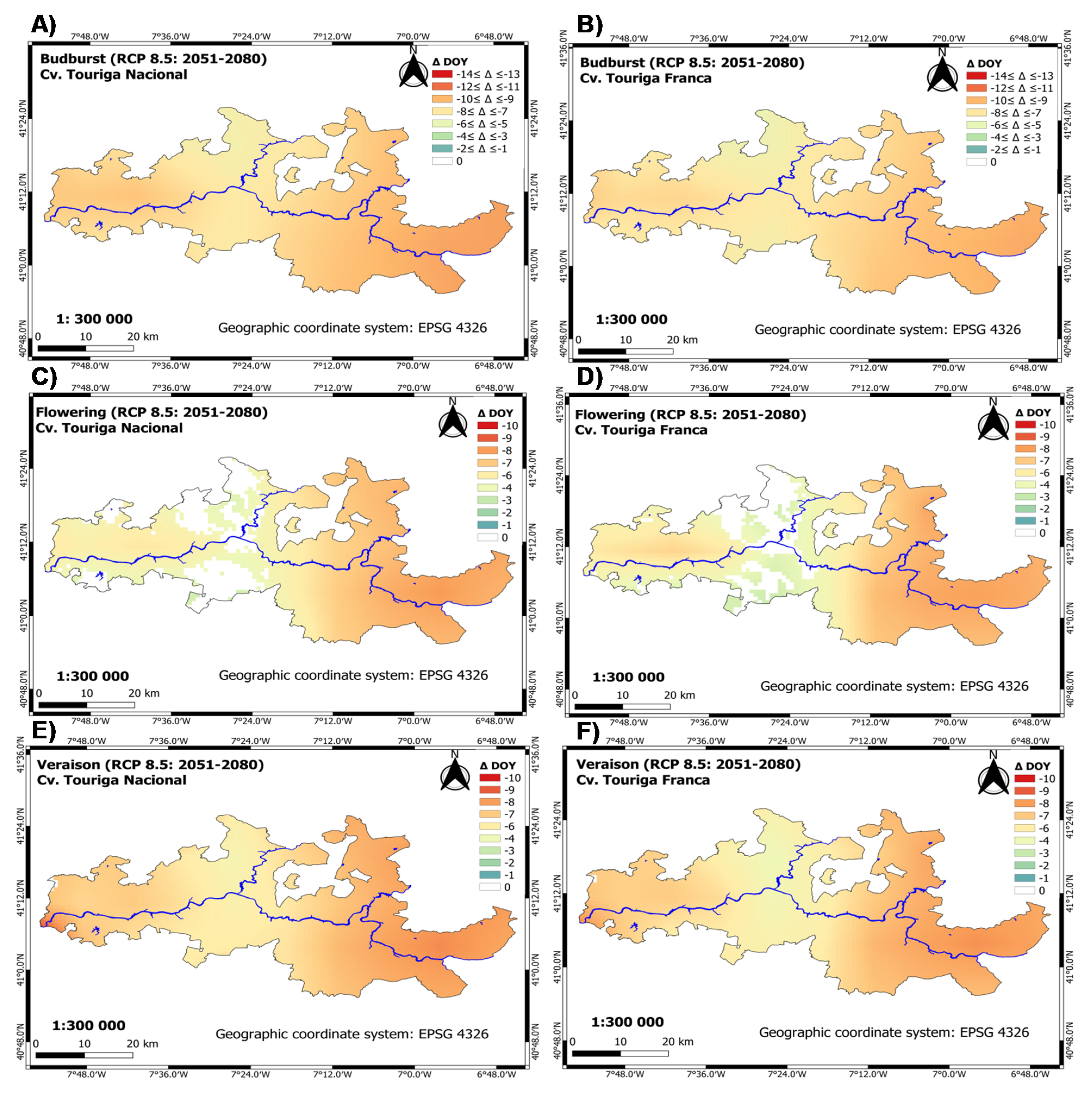
3.1.2. Future Period: Climate Scenarios
3.2. LB Flight Phenology
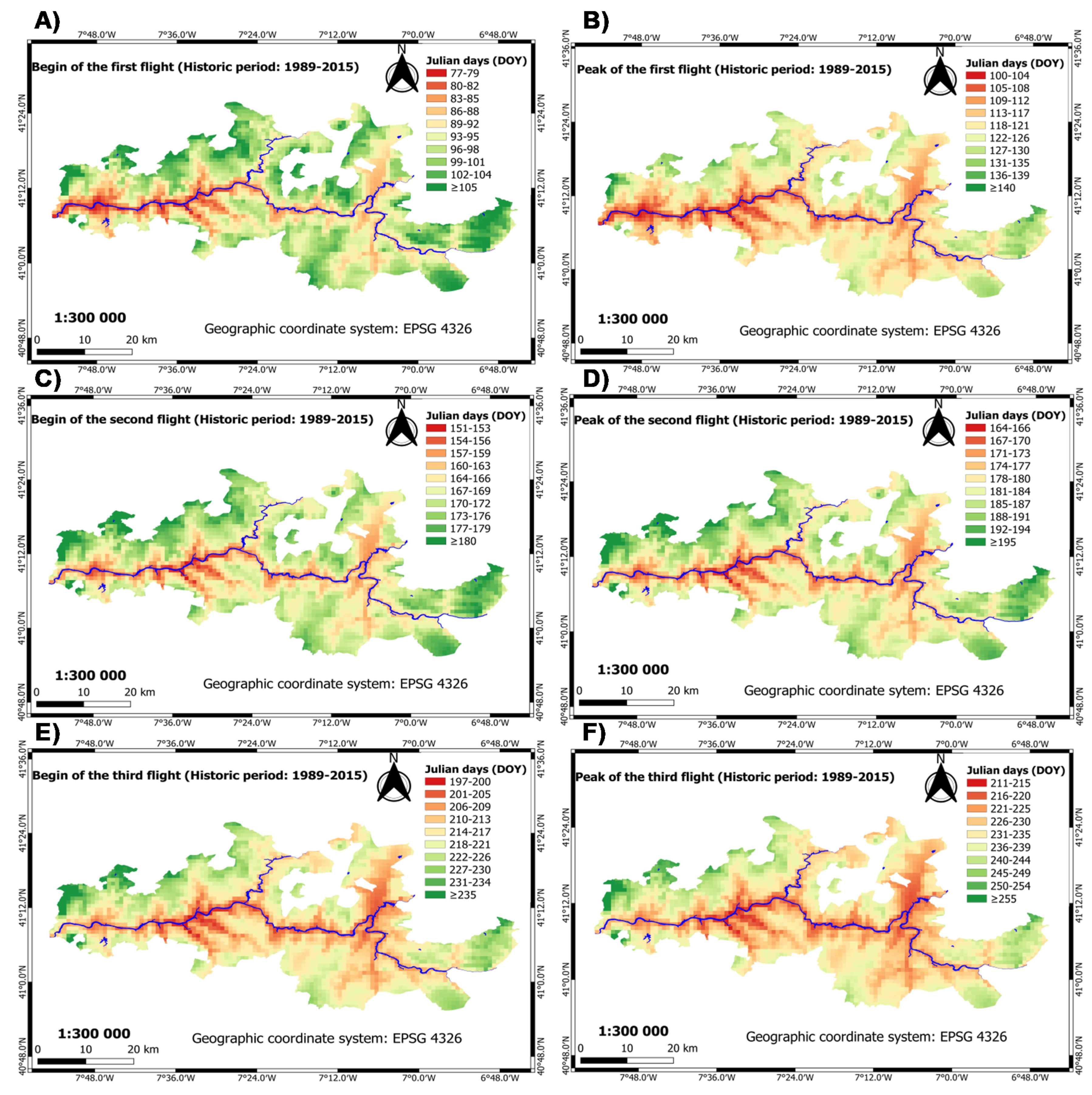
3.2.1. Historic Period
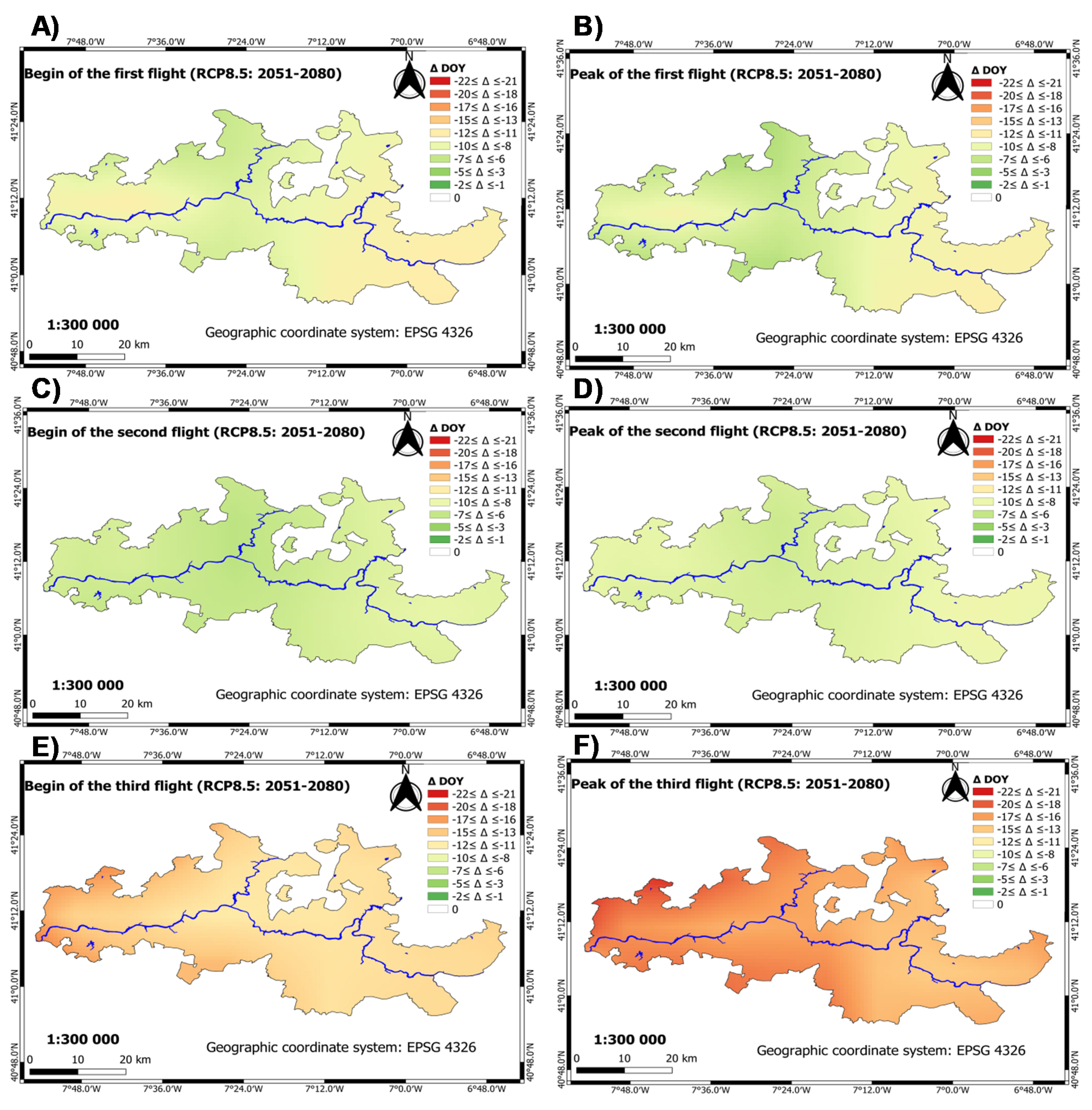
3.2.2. Future Period: Climate Scenarios
3.3. Interaction of Trophic Levels: LB and Grapevine
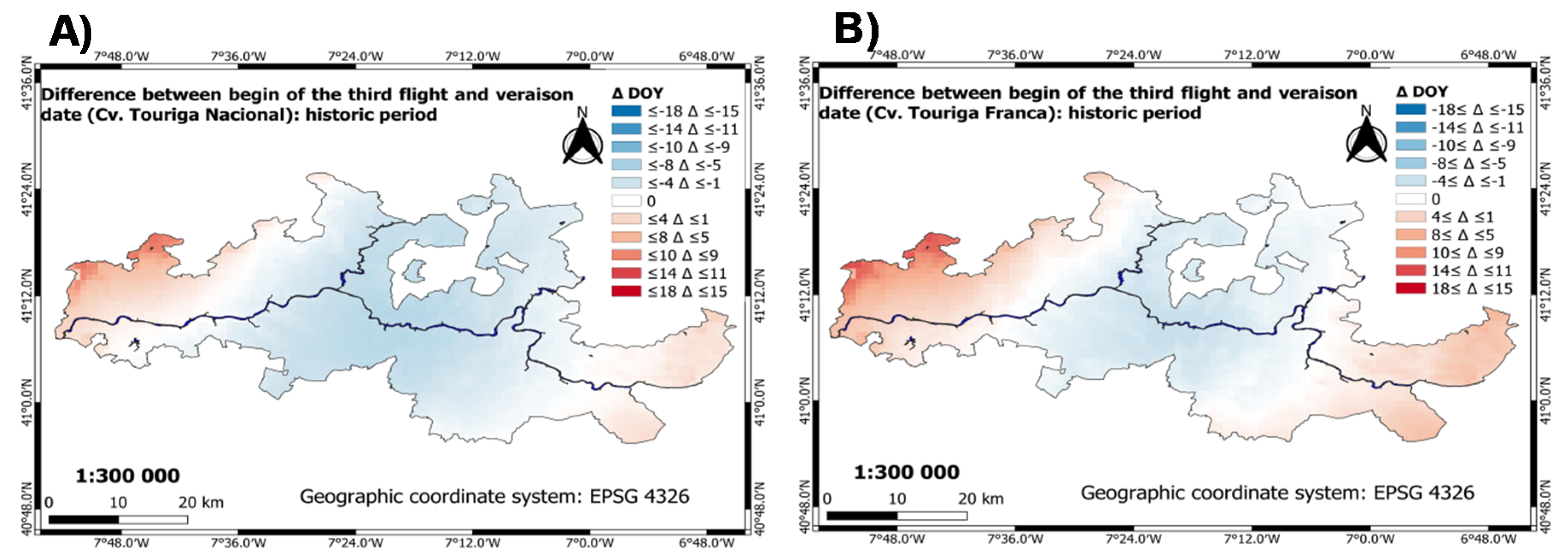
3.3.1. Historic Period
3.3.2. Future Period
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Castex, V.; Beniston, M.; Calanca, P.; Fleury, D.; Moreau, J. Pest management under climate change: The importance of understanding tritrophic relations. Sci. Total Environ. 2018, 616–617, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Reineke, A.; Thiéry, D. Grapevine insect pests and their natural enemies in the age of global warming. J. Pest Sci. 2016, 89, 313–328. [Google Scholar] [CrossRef]
- Caffarra, A.; Rinaldi, M.; Eccel, E.; Rossi, V.; Pertot, I. Modelling the impact of climate change on the interaction between grapevine and its pests and pathogens: European grapevine moth and powdery mildew. Agric. Ecosyst. Environ. 2012, 148, 89–101. [Google Scholar] [CrossRef]
- Castex, V. The Impact of Climate Change on the Development of Agricultural Pests, Their Hosts and Natural Enemies: Assessing Overlaps in Trophic Interactions. Ph.D. Thesis, Université de Genève, Geneva, Switzerland, 2019. [Google Scholar]
- Reis, S.; Fraga, H.; Carlos, C.; Silvestre, J.; Eiras-Dias, J.; Rodrigues, P.; Santos, J.A. Grapevine Phenology in Four Portuguese Wine Regions: Modeling and Predictions. Appl. Sci. 2020, 10, 3708. [Google Scholar] [CrossRef]
- Amerine, M.; Winkler, A. Composition and Quality of Musts and Wines of California Grapes. Hilgardia 1944, 15, 493–675. [Google Scholar] [CrossRef]
- Pouget, R. Etude du Rhythme végétative: Caractères physiologiques liées à la précocité de débourrement chez la vigne. Ann. Amelior. Plantes 1966, 16, 81–100. [Google Scholar]
- Riou, C. The Effect of Climate on Grape Ripening: Application to the Zoning of Sugar Content in the European Community; Office des Publications Officielles des Communautés Européennes: Luxembourg, 1994; p. 322. [Google Scholar]
- Hänninen, H. Modelling Bud Dormancy Release in Trees from Cool and Temperate Regions; Society of Forestry in Finland: Helsinki, Finland, 1990; Volume 213. [Google Scholar]
- Wang, E.; Engel, T. Simulation of phenological development of wheat crops. Agric. Syst. 1998, 58, 1–24. [Google Scholar] [CrossRef]
- Parker, A.K.; Cortazar-Atauri, I.G.; Leeuwen, C.; Chuine, I. General phenological model to characterise the timing of flowering and veraison of Vitis vinifera L. Aust. J. Grape Wine Res. 2011, 17, 206–216. [Google Scholar] [CrossRef]
- Duchêne, E.; Huard, F.; Vincent, D.; Schneider, C.; Merdinoglu, D. The challenge of adapting grapevine varieties to climate change. Clim. Res. 2010, 41, 193–204. [Google Scholar] [CrossRef]
- Molitor, D.; Caffarra, A.; Sinigoj, P.; Pertot, I.; Hoffmann, L.; Junk, J. Late frost damage risk for viticulture under future climate conditions: A case study for the Luxembourgish winegrowing region. Aust. J. Grape Wine Res. 2014, 20, 160–168. [Google Scholar] [CrossRef]
- Cuccia, C.; Bois, B.; Richard, Y.; Parker, A.; Garcia de Cortazar-Atauri, I.; Cornelis, V.; van Leeuwen, C.; Castel, T. Phenological model performance to warmer conditions: Application to Pinot noir in Burgundy. J. Int. Sci. Vigne Vin 2014, 48, 169–178. [Google Scholar] [CrossRef]
- Garcia de Cortazar-Atauri, I.; Duchêne, E.; Destrac Irvine, A.; Barbeau, G.; Rességuier, L.; Lacombe, T.; Parker, A.; Saurin, N.; van Leeuwen, C. Grapevine phenology in France: From past observations to future evolutions in the context of climate change. OENO One 2017, 51, 115. [Google Scholar] [CrossRef]
- Milonas, P.G.; Savopoulou-Soultani, M.; Stavridis, D.G. Day-degree models for predicting the generation time and flight activity of local populations of Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) in Greece. J. Appl. Entomol. 2001, 125, 515–518. [Google Scholar]
- Carlos, C.; Gonçalves, F.; Oliveira, I.; Torres, L. Is a biofix necessary for predicting the flight phenology of Lobesia botrana in Douro Demarcated Region vineyards? Crop Prot. 2018, 110, 57–64. [Google Scholar] [CrossRef]
- Ortega-Lopez, V.; Amo-Salas, M.; Ortiz-Barredo, A.; Diez-Navajas, A.M. Male flight phenology of the European grapevine moth Lobesia botrana (Lepidoptera: Tortricidae) in different wine-growing regions in Spain. Bull. Entomol. Res. 2014, 104, 566–575. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Ponti, L.; Gilioli, G.; Baumgärtner, J. Climate warming effects on grape and grapevine moth (Lobesia botrana) in the Palearctic region. Agric. For. Entomol. 2018, 20, 255–271. [Google Scholar] [CrossRef]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 531–544. [Google Scholar] [CrossRef]
- Honêk, A. Geographical variation in thermal requirements for insect development. EJE 1996, 93, 303–312. [Google Scholar]
- Reis, S.; Martins, J.; Gonçalves, F.; Carlos, C.; Santos, J.A. European grapevine moth in the Douro region: Voltinism and climatic scenarios. OENO One 2021, 55, 335–351. [Google Scholar] [CrossRef]
- Martín-Vertedor, D.; Ferrero-García, J.; Torres-Vila, L.M. Global warming affects phenology and voltinism of Lobesia botrana in Spain. Agric. For. Entomol. 2010, 12, 169–176. [Google Scholar] [CrossRef]
- Kalinkat, G.; Rall, B.C. Effects of Climate Change on the Interactions between Insect Pests and Their Natural Enemies. In Climate Change and Insect Pests; CABI: Wallingford, UK, 2015. [Google Scholar]
- Andresen, T.; de Aguiar, F.B.; Curado, M.J. The Alto Douro Wine Region greenway. Landsc. Urban Plan. 2004, 68, 289–303. [Google Scholar] [CrossRef]
- Santos, M.; Fonseca, A.; Fraga, H.; Jones, G.; Santos, J.A. Bioclimatic conditions of the Portuguese wine denominations of origin under changing climates. Int. J. Climatol. 2020, 40, 927–941. [Google Scholar] [CrossRef]
- Costa, R.; Fraga, H.; Fonseca, A.; García de Cortázar-Atauri, I.; Val, M.C.; Carlos, C.; Reis, S.; Santos, J.A. Grapevine phenology of cv. Touriga franca and Touriga nacional in the Douro wine region: Modelling and climate change projections. Agronomy 2019, 9, 210. [Google Scholar] [CrossRef]
- Jones, G. Uma Avaliação do Clima para a Região Demarcada do Douro: Uma análise das condições climáticas do passado, presente e futuro para a produção de vinho. In ADVID—Associação Para o Desenvolvmento da Viticultura Duriense; ADVID—Associação para o Desenvolvmento da Viticultura Duriense: Vila Real, Portugal, 2013; p. 110. [Google Scholar]
- ADVID. Impact of Climate Change on the Douro Wine Region: Study Proposal. Association for the Development of Viticulture in the Douro Region. December 2007, ADVID. Available online: www.advid.pt (accessed on 1 October 2021).
- Fraga, H.; García de Cortázar Atauri, I.; Malheiro, A.C.; Moutinho-Pereira, J.; Santos, J.A. Viticulture in Portugal: A review of recent trends and climate change projections. OENO One 2017, 51, 61–69. [Google Scholar] [CrossRef]
- Fraga, H.; Costa, R.; Santos, J.A. Modelling the Terroir of the Douro Demarcated Region, Portugal. In E3S Web of Conferences; EDP Sciences: Les Ulis, France, 2018; p. 02009. [Google Scholar]
- Lorenz, D.H.; Eichhorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Growth Stages of the Grapevine: Phenological growth stages of the grapevine (Vitis vinifera L. ssp. vinifera)—Codes and descriptions according to the extended BBCH scale†. Aust. J. Grape Wine Res. 1995, 1, 100–103. [Google Scholar] [CrossRef]
- Baillod, M.; Baggiolini, M. Les stades repères de la vigne. Rev. Suisse Vitic. Arboric. Hortic 1993, 25, 7–9. [Google Scholar]
- Gutierrez, A.P.; Ponti, L.; Cooper, M.L.; Gilioli, G.; Baumgärtner, J.; Duso, C. Prospective analysis of the invasive potential of the European grapevine moth Lobesia botrana (Den. & Schiff.) in California. Agric. For. Entomol. 2012, 14, 225–238. [Google Scholar]
- Gilioli, G.; Pasquali, S.; Marchesini, E. A modelling framework for pest population dynamics and management: An application to the grape berry moth. Ecol. Model. 2016, 320, 348–357. [Google Scholar] [CrossRef]
- Baumgärtner, J.; Gutierrez, A.P.; Pesolillo, S.; Severini, M. A model for the overwintering process of European grapevine moth Lobesia botrana (Denis & Schiffermüller) (Lepidoptera, Tortricidae) populations. J. Entomol. Acarol. Res. 2012, 44, e2. [Google Scholar] [CrossRef][Green Version]
- García de Cortázar-Atauri, I.; Brisson, N.; Gaudillere, J.P. Performance of several models for predicting budburst date of grapevine (Vitis vinifera L.). Int. J. Biometeorol. 2009, 53, 317–326. [Google Scholar] [CrossRef]
- Santos, J.A.; Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Dinis, L.T.; Correia, C.; Moriondo, M.; Leolini, L.; Dibari, C.; Costafreda-Aumedes, S.; et al. A Review of the Potential Climate Change Impacts and Adaptation Options for European Viticulture. Appl. Sci. 2020, 10, 3092. [Google Scholar] [CrossRef]
- Cornes, R.C.; van der Schrier, G.; van den Besselaar, E.J.M.; Jones, P.D. An Ensemble Version of the E-OBS Temperature and Precipitation Data Sets. J. Geophys. Res. Atmos. 2018, 123, 9391–9409. [Google Scholar] [CrossRef]
- Van den Besselaar, E.J.M.; Haylock, M.R.; van der Schrier, G.; Klein Tank, A.M.G. A European daily high-resolution observational gridded data set of sea level pressure. J. Geophys. Res. Atmos. 2011, 116, D11110. [Google Scholar] [CrossRef]
- Hofstra, N.; Haylock, M.R.; New, M.; Jones, P. Testing E-OBS European high-resolution gridded data set of daily precipitation and surface temperature. J. Geophys. Res. Atmos. D Atmos. 2009, 114, D21101. [Google Scholar] [CrossRef]
- Fonseca, A.R.; Santos, J.A. High-resolution temperature datasets in Portugal from a geostatistical approach: Variability and extremes. J. Appl. Meteorol. Climatol. 2018, 57, 627–644. [Google Scholar] [CrossRef]
- Giorgi, F.; Jones, C.; Asrar, G.R. Addressing climate information needs at the regional level: The CORDEX framework. WMO Bull. 2009, 58, 175–183. [Google Scholar]
- Jacob, D.; Petersen, J.; Eggert, B.; Alias, A.; Christensen, O.B.; Bouwer, L.M.; Braun, A.; Colette, A.; Déqué, M.; Georgievski, G.; et al. EURO-CORDEX: New high-resolution climate change projections for European impact research. Reg. Environ. Chang. 2014, 14, 563–578. [Google Scholar] [CrossRef]
- Dyrrdal, A.V.; Stordal, F.; Lussana, C. Evaluation of summer precipitation from EURO-CORDEX fine-scale RCM simulations over Norway. Int. J. Climatol. 2018, 38, 1661–1677. [Google Scholar] [CrossRef]
- Spinoni, J.; Barbosa, P.; Bucchignani, E.; Cassano, J.; Cavazos, T.; Christensen, J.; Christensen, O.B.; Coppola, E.; Evans, J.; Geyer, B. Future global meteorological drought hot spots: A study based on CORDEX data. J. Clim. 2020, 33, 3635–3661. [Google Scholar] [CrossRef]
- Martins, J.; Fraga, H.; Fonseca, A.; Santos, J.A. Climate Projections for Precipitation and Temperature Indicators in the Douro Wine Region: The Importance of Bias Correction. Agronomy 2021, 11, 990. [Google Scholar] [CrossRef]
- Welch, B.L. The generalization of ‘student’s’ problem when several different population varlances are involved. Biometrika 1947, 34, 28–35. [Google Scholar] [CrossRef]
- Fagerland, M.W.; Sandvik, L. Performance of five two-sample location tests for skewed distributions with unequal variances. Contemp. Clin. Trials 2009, 30, 490–496. [Google Scholar] [CrossRef]
- Ruxton, G.D. The unequal variance t-test is an underused alternative to Student’s t-test and the Mann–Whitney U test. Behav. Ecol. 2006, 17, 688–690. [Google Scholar] [CrossRef]
- Nagarkatti, S.; Tobin, P.C.; Saunders, M.C. Diapause Induction in the Grape Berry Moth (Lepidoptera: Tortricidae). Environ. Entomol. 2001, 30, 540–544. [Google Scholar] [CrossRef]
- Fraga, H.; Santos, J.A.; Moutinho-Pereira, J.; Carlos, C.; Silvestre, J.; Eiras-Dias, J.; Mota, T.; Malheiro, A.C. Statistical modelling of grapevine phenology in Portuguese wine regions: Observed trends and climate change projections. J. Agric. Sci. 2015, 154, 795–811. [Google Scholar] [CrossRef]
- Droulia, F.; Charalampopoulos, I. Future Climate Change Impacts on European Viticulture: A Review on Recent Scientific Advances. Atmosphere 2021, 12, 495. [Google Scholar] [CrossRef]
- Morales-Castilla, I.; García de Cortázar-Atauri, I.; Cook, B.I.; Lacombe, T.; Parker, A.; van Leeuwen, C.; Nicholas, K.A.; Wolkovich, E.M. Diversity buffers winegrowing regions from climate change losses. Proc. Natl. Acad. Sci. USA 2020, 117, 2864–2869. [Google Scholar] [CrossRef] [PubMed]
- Tomasi, D.; Jones, G.V.; Giust, M.; Lovat, L.; Gaiotti, F. Grapevine Phenology and Climate Change: Relationships and Trends in the Veneto Region of Italy for 1964–2009. Am. J. Enol. Vitic. 2011, 62, 329–339. [Google Scholar] [CrossRef]
- Caffarra, A.; Eccel, E. Projecting the impacts of climate change on the phenology of grapevine in a mountain area. Aust. J. Grape Wine Res. 2011, 17, 52–61. [Google Scholar] [CrossRef]
- Taylor, R.A.J.; Herms, D.A.; Cardina, J.; Moore, R.H. Climate change and pest management: Unanticipated consequences of trophic dislocation. Agronomy 2018, 8, 7. [Google Scholar] [CrossRef]
- Damien, M.; Tougeron, K. Prey–predator phenological mismatch under climate change. Curr. Opin. Insect Sci. 2019, 35, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Stange, E.E.; Ayres, M.P. Climate change impacts: Insects. In Encyclopedia of Life Sciences (ELS); John Wiley & Sons, Ltd.: Chichester, UK, 2010. [Google Scholar]
- Schweiger, O.; Settele, J.; Kudrna, O.; Klotz, S.; Kühn, I. Climate change can cause spatial mismatch of trophically interacting species. Ecology 2008, 89, 3472–3479. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reis, S.; Martins, J.; Gonçalves, F.; Carlos, C.; Santos, J.A. European Grapevine Moth and Vitis vinifera L. Phenology in the Douro Region: (A)synchrony and Climate Scenarios. Agronomy 2022, 12, 98. https://doi.org/10.3390/agronomy12010098
Reis S, Martins J, Gonçalves F, Carlos C, Santos JA. European Grapevine Moth and Vitis vinifera L. Phenology in the Douro Region: (A)synchrony and Climate Scenarios. Agronomy. 2022; 12(1):98. https://doi.org/10.3390/agronomy12010098
Chicago/Turabian StyleReis, Samuel, Joana Martins, Fátima Gonçalves, Cristina Carlos, and João A. Santos. 2022. "European Grapevine Moth and Vitis vinifera L. Phenology in the Douro Region: (A)synchrony and Climate Scenarios" Agronomy 12, no. 1: 98. https://doi.org/10.3390/agronomy12010098
APA StyleReis, S., Martins, J., Gonçalves, F., Carlos, C., & Santos, J. A. (2022). European Grapevine Moth and Vitis vinifera L. Phenology in the Douro Region: (A)synchrony and Climate Scenarios. Agronomy, 12(1), 98. https://doi.org/10.3390/agronomy12010098