Rapeseed (Brassica napus): Processing, Utilization, and Genetic Improvement


Abstract
1. Introduction
2. Rapeseed Seeds Processing
2.1. Hexane Free Oil Extraction
2.2. Aqueous Extraction
2.3. Heat Treatments
2.4. Chemical-Free Extraction for Better Protein Quality
2.5. Preparing Biodiesel from Rapeseed
3. Seed Oil and Protein-Processing Products
3.1. Edible Oil
3.1.1. Biochemical Properties
3.1.2. Health Benefits of Rapeseed Oil
3.1.3. Multiple Usages of Edible Rapeseed Oil
3.2. Inedible Oil
3.2.1. Biodiesel
3.2.2. Other Industrial Applications of Rapeseed Oil
3.3. Rapeseed Meal
3.3.1. Meal Composition
3.3.2. Human Food Application
3.3.3. Animal Fodder
3.3.4. Protein for Bioplastic Based Materials and Cosmetics Fabrication
4. Utilization of Natural Unprocessed Rapeseed
4.1. Vegetable Food, Tea, and Homeopathy
4.2. Honey Production from Rapeseed
4.3. Ornament and Field Attraction
4.4. Agricultural Residues
4.5. Phytoremediation
5. Advancement in Breeding Aiming to Improve Rapeseed
5.1. Hybrid Heterotic Potential
5.2. Pollination Control Systems for Hybrid Seed Production
5.3. Exotic Germplasm to Enhance Heterotic Potential
5.4. Genomic Selection (GS)
5.5. Nested Association Mapping (NAM) and Multi–Parents Advanced Generation Intercrosses (MAGIC)
5.6. Genome Sequencing
5.7. Genome Editing Technologies (GETs)
6. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Al-Shehbaz, I.A.; Beilstein, M.A.; Kellogg, E.A. Systematics and phylogeny of the Brassicaceae (Cruciferae): An overview. Plant Syst. Evol. 2006, 259, 89–120. [Google Scholar] [CrossRef]
- Meyer, R.S.; Purugganan, M.D. Evolution of crop species: Domestication and diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef]
- Canola Council of Canada. About Canola. Available online: https://www.canolacouncil.org/about-canola/ (accessed on 6 July 2021).
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Snowdon, R.; Lühs, W.; Friedt, W. Oilseed Rape: Chapter 2. In Genome Mapping and Molecular Breeding in Plants 2. Oilseeds; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar] [CrossRef]
- Shahzadi, T.; Khan, F.A.; Zafar, F.; Ismail, A.; Amin, E.; Riaz, S. An Overview of Brassica Species for Crop Improvement. Am.-Eurasian J. Agric. Environ. Sci. 2015, 15, 1568–1573. [Google Scholar] [CrossRef]
- Daun, J.K. Origin, Distribution, and Production. In Canola; Elsevier: Amsterdam, The Netherlands, 2011; pp. 1–28. [Google Scholar] [CrossRef]
- FAOSTAT. Crops and Livestock Products (Production). Available online: http://www.fao.org/faostat/en/#search/Rapeseed%20 (accessed on 24 August 2021).
- STATISTA. Worldwide Production of Rapeseed by Country. Available online: https://www.statista.com/statistics/263930/worldwide-production-of-rapeseed-by-country (accessed on 24 August 2021).
- World Rapeseed Production. Available online: https://www.worldagriculturalproduction.com/crops/rapeseed.aspx (accessed on 6 July 2021).
- Gulden, R.H.; Warwick, S.I.; Thomas, A.G. The biology of Canadian weeds. 137. Brassica napus L. and B. rapa L. Can. J. Plant Sci. 2008, 88, 951–996. [Google Scholar] [CrossRef]
- STATISTA. Worldwide Oilseed Production since 2008. Available online: https://www.statista.com/statistics/267271/worldwide-oilseed-production-since-2008 (accessed on 8 July 2020).
- Wang, Y.; Hou, Y.; Chen, C.; Zhou, M.G. Detection of resistance in Sclerotinia sclerotiorum to carbendazim and dimethachlon in Jiangsu Province of China. Australas. Plant Pathol. 2014, 43, 307–312. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Y.; Li, X.L.; Li, H.Z. Research progress in clubroot of crucifers. Plant Prot. 2011, 37, 153–158. [Google Scholar]
- Unger, E.H. Processing. In Canola; Elsevier: Amsterdam, The Netherlands, 2011; pp. 163–189. [Google Scholar] [CrossRef]
- Canola Council of Canada. Processing Canola Seed into Oil and Meal in Canada. Available online: https://www.canolacouncil.org/about-canola/processing/ (accessed on 6 July 2021).
- Confortin, T.C.; Todero, I.; Luft, L.; Ugalde, G.A.; Mazutti, M.A.; Oliveira, Z.B.; Bottega, E.L.; Knies, A.E.; Zabot, G.L.; Tres, M.V. Oil yields, protein contents, and cost of manufacturing of oil obtained from different hybrids and sowing dates of canola. J. Environ. Chem. Eng. 2019, 7, 102972. [Google Scholar] [CrossRef]
- Swanson, R.G. Hexane Extraction in Soyfoods Processing; Regents Professor; Department of Food Science, Washington State University: Pullman, WA, USA, 2009. [Google Scholar]
- Gaber, M.A.F.M.; Tujillo, F.J. Improving oil extraction from canola seeds by conventional and advanced methods. Food Eng. Rev. 2018, 10, 198–210. [Google Scholar] [CrossRef]
- Lv, M.; Wu, W. An advanced aqueous method of extracting rapeseed oil with high quality. J. Food Process Eng. 2018, 42, e12957. [Google Scholar] [CrossRef]
- Fetzer, A.; Herfellner, T.; Stäbler, A.; Menner, M.; Eisner, P. Influence of process conditions during aqueous protein extraction upon yield from pre-pressed and cold-pressed rapeseed press cake. Ind. Crops Prod. 2018, 112, 236–246. [Google Scholar] [CrossRef]
- Gerzhova, A.; Mondor, M.; Benali, M.; Aider, M. A comparative study between the electro-activation technique and conventional extraction method on the extractability, composition and physicochemical properties of canola protein concentrates and isolates. Food Biosci. 2015, 11, 56–71. [Google Scholar] [CrossRef]
- Akbari, A.; Wu, J. An integrated method of isolating napin and cruciferin from defatted canola meal. Food Sci. Technol. 2015, 64, 308–315. [Google Scholar] [CrossRef]
- Hénon, G.; Kemény, Z.; Recseg, K.; Zwobada, F.; Kovari, K. Deodorization of Vegetable Oils. Part I: Modeling the Geometrical Isomerization of Polyunsaturated Fatty Acids. J. Am. Oil Chem. Soc. 1999, 76, 73–81. [Google Scholar] [CrossRef]
- Aladedunye, F.A.; Przybylski, R. Degradation and Nutritional Quality Changes of Oil during Frying. J. Am. Oil Chem. Soc. 2009, 86, 149–156. [Google Scholar] [CrossRef]
- Taghvaei, M.; Jafari, S.M. Application and stability of natural antioxidants in edible oils in order to substitute synthetic additives. J. Food Sci. Technol. 2015, 52, 1272–1282. [Google Scholar] [CrossRef] [PubMed]
- Aniolowska, M.; Zahran, H.; Kita, A. The effect of pan frying on thermooxidative stability of refined rapeseed oil and professional blend. J. Food Sci. Technol. 2016, 53, 712–720. [Google Scholar] [CrossRef]
- Wroniak, M.; Rękas, A. Nutritional value of cold-pressed rapeseed oil during long term storage as influenced by the type of packaging material, exposure to light & oxygen and storage temperature. J. Food Sci. Technol. 2016, 53, 1338–1347. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, R.S.; Wang, Z.G.; Wang, B.; Yang, Y.J.; Ju, X.R.; He, R. The effect of refining process on the physicochemical properties and micronutrients of rapeseed oils. PLoS ONE 2019, 14, e0212879. [Google Scholar] [CrossRef]
- Raikos, V.; Neacsu, M.; Duthie, G.; Nicol, F.; Reid, M.; Cantlay, L.L.; Ranawana, V. Proteomic and glucosinolate profiling of rapeseed isolates from meals produced by different oil extraction processes. J. Food Process. Preserv. 2017, 41, e13060. [Google Scholar] [CrossRef]
- Schutyser, M.A.I.; Pelgrom, P.J.M.; van der Goot, A.J.; Boom, R.M. Dry fractionation for sustainable production of functional legume protein concentrates. Trends Food Sci. Technol. 2015, 45, 327–335. [Google Scholar] [CrossRef]
- Barakat, A.; Mayer-Laigle, C. Electrostatic Separation as an Entry into Environmentally EcoFriendly Dry Biorefining of Plant Materials. J. Chem. Eng. Process Technol. 2017, 8, 4–354. [Google Scholar] [CrossRef]
- Mayer-Laigle, C.; Barakat, A.; Barron, C.; Delenne, J.Y.; Frank, X.; Mabille, F.; Rouau, X.; Sadoudi, A.; Samson, M.F.; Lullien-Pellerin, V. DRY biorefineries: Multiscale modeling studies and innovative processing. Innov. Food Sci. Emerg. Technol. 2017, 46, 131–139. [Google Scholar] [CrossRef]
- Jafari, M.; Rajabzadeh, A.R.; Tabtabaei, S.; Marsolais, F.; Legge, R.L. Physicochemical characterization of a navy bean (Phaseolus vulgaris) protein fraction produced using a solvent-free method. Food Chem. 2016, 208, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Laguna, O.; Barakat, A.; Alhamada, H.; Durand, E.; Baréa, B.; Fine, F.; Villeneuve, P.; Citeau, M.; Dauguet, S.; Lecomte, J. Production of proteins and phenolic compounds enriched fractions from rapeseed and sunflower meals by dry fractionation processes. Ind. Crops Prod. 2018, 118, 160–172. [Google Scholar] [CrossRef]
- Kdidi, S.; Vaca-Medina, G.; Peydecastaing, J.; Oukarroum, A.; Fayoud, N.; Barakat, A. Electrostatic separation for sustainable production of rapeseed oil cake protein concentrate: Effect of mechanical disruption on protein and lignocellulosic fiber separation. Powder Technol. 2019, 344, 10–16. [Google Scholar] [CrossRef]
- Mejicanos, G.A.; Regassa, A.; Nyachoti, C.M. Effect of high canola meal content on growth performance, nutrient digestibility and fecal bacteria in nursery pigs fed either corn or wheat based diets. Anim. Feed Sci. Technol. 2017, 231, 59–66. [Google Scholar] [CrossRef]
- Xu, L.; Diosady, L.L. Removal of phenolic compounds in the production of high-quality canola protein isolates. Food Res. 2002, 35, 23–30. [Google Scholar] [CrossRef]
- Hald, C.; Dawid, C.; Tressel, R.; Hofmann, T. Kaempferol 3-O-(2‴-O-Sinapoyl-β-sophoroside) Causes the Undesired Bitter Taste of Canola/Rapeseed Protein Isolates. J. Agric. Food Chem. 2019, 67, 372–378. [Google Scholar] [CrossRef]
- Zago, E.; Lecomte, J.; Barouh, N.; Aouf, C.; Carré, P.; Fine, F.; Villeneuve, P. Influence of rapeseed meal treatments on its total phenolic content and composition in sinapine, sinapic acid and canolol. Ind. Crops Prod. 2015, 76, 1061–1070. [Google Scholar] [CrossRef]
- Thuneke, K.; Remmele, E.; Widmann, B.; Wilharm, T. Standardisation of rapeseed oil as a fuel. In Proceedings of the 1st World Conference on Biomass for Energy and Industry, Sevilla, Spain, 5–9 June 2000. [Google Scholar]
- Vijayaraghavan, K.; Hemanathan, K. Biodiesel production from freshwater algae. Energy Fuels 2009, 23, 5448. [Google Scholar] [CrossRef]
- Kai, T.; Mak, G.L.; Wada, S.; Nakazato, T.; Takanashi, H.; Uemura, Y. Production of biodiesel fuel from canola oil with dimethyl carbonate using an active sodium methoxide catalyst prepared by crystallization. Bioresour. Technol. 2014, 163, 360–363. [Google Scholar] [CrossRef]
- Dworakowska, S.; Bednarz, S.; Bogdal, D. Production of biodiesel from rapeseed oil. In Proceedings of the 1st World Sustainability Forum, Basel, Switzerland, 1–30 November 2011. [Google Scholar]
- Ma, F.; Clements, L.D.; Hanna, M.A. The effects of catalyst, free fatty acids and water on transesterification of beef tallow. Trans. ASAE 1998, 41, 1261–1264. [Google Scholar] [CrossRef]
- Tyson, K.S.; Bozell, J.; Wallace, R.; Petersen, E.; Moens, L. Biomass Oil Analysis: Research Needs and Recommendations; NREL/TP-510-34796; National Renewable Energy Laboratory, US Department of Energy: Washington, DC, USA, 2004. [Google Scholar] [CrossRef]
- Goembira, F.; Matsuura, K.; Saka, S. Biodiesel production from rapeseed oil by various supercritical carboxylate esters. Fuel 2012, 97, 373–378. [Google Scholar] [CrossRef]
- Canadian Food Inspection Agency. The Biology of Brassica napus L. (Canola/Rapeseed). Available online: https://inspection.canada.ca/plant-varieties/plants-with-novel-traits/applicants/directive-94-08/biology-documents/brassica-napus-l-/eng/1330729090093/1330729278970 (accessed on 25 August 2021).
- CanolaInfo. Comparison of Dietary Fats. Available online: https://www.canolainfo.org/quadrant/media/files/downloads/pdfs/DietaryFatChartprint2016.pdf (accessed on 6 July 2021).
- Liu, H.; Biliaderis, C.G.; Przybylski, R.; Eskin, N.A.M. Phase transitions of canola sediments. J. Am. Oil Chem. Soc. 1993, 70, 441–448. [Google Scholar] [CrossRef]
- Ackman, R.G. Rapeseed Fatty Acids—An Ideal Mixture for Health, Nutrition, and Food Use. In Canola and Rapeseed. Production, Chemistry, Nutrition and Processing Technology; Shahidi, F., Ed.; Avi Book; Van Nostrand Reinhold: New York, NY, USA, 1990; pp. 81–98. [Google Scholar] [CrossRef]
- Vecchio, A.J. High-laurate rapeseed. INFORM 1996, 7, 230–243. [Google Scholar]
- Tso, P.; Ding, S.; DeMichele, K.; Huang, Y.S. Intestinal Absorption of High γ-Linolenic Acid Canola Oil in Lymph Fistula Rats. In γ-Linolenic Acid: Recent Advances in Biotechnology and Clinical Applications; Huang, Y.-S., Ziboh, V.A., Eds.; AOCS Press: Champaign, IL, USA, 2001; pp. 321–334. [Google Scholar] [CrossRef]
- Pongracz, G. Hitzestabilitaet der tocopherol. Fat Sci. Technol. 1988, 90, 249–258. [Google Scholar]
- Matthäus, B. High oleic low linolenic rapeseed oil as alternative to common used frying oils. Qual. Nutr. Process. Process. Technol. 2006, 108, 200–211. [Google Scholar]
- Cmolik, J.; Pokorny, J.; Reblova, Z.; Svoboda, Z. Tocopherol retention in physically refined rapeseed oil as a function of deodorization temperature. Eur. J. Lipid Sci. Technol. 2008, 110, 754–759. [Google Scholar] [CrossRef]
- Rudzinska, M.; Wasowicz, E.; Uchman, W. Plant sterols in food technology. Acta Sci. Pol. Technol. Aliment. 2005, 4, 147–156. [Google Scholar]
- Ryan, E.; Chopra, J.; McCarthy, F.; Maguire, A.R.; O’Brien, N.M. Qualitative and quantitative comparison of the cytotoxic and apoptotic potential of phytosterol oxidation products with their corresponding cholesterol oxidation products. Br. J. Nutr. 2005, 94, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Hovenkamp, E.; Demonty, I.; Plat, J.; Lutjohann, D.; Mensink, R.P.; Trautwein, E.A. Biological effects of oxidized phytosterols: A review of the current knowledge. Prog. Lipid Res. 2008, 47, 37–49. [Google Scholar] [CrossRef]
- Javitt, N.B. Oxysterols: Novel biologic roles for the 21st century. Steroids 2008, 73, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Koski, A.; Pekkarinen, S.; Hopia, A.; Wahala, K.; Heinonen, M. Processing of rapeseed oil: Effects on sinapic acid derivative content and oxidative stability. Eur. Food Res. Technol. 2003, 217, 110–114. [Google Scholar] [CrossRef]
- USDA. Food Data Central. Available online: https://fdc.nal.usda.gov/ (accessed on 27 August 2021).
- Institute of Medicine. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; National Academies Press: Washington, DC, USA, 2005. [Google Scholar] [CrossRef]
- Mensink, R.P.; Zock, P.L.; Kester, A.D.; Katan, M.B. Effects of dietary fatty acids and carbohydrates on the ratio of serum total to HDL cholesterol and on serum lipids and apolipoproteins: A meta-analysis of 60 controlled trials. Am. J. Clin. Nutr. 2003, 77, 1146–1155. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; AHA Science Advisory. Monounsaturated fatty acids and risk of cardiovascular disease. Circulation 1999, 100, 1253–1258. [Google Scholar] [CrossRef]
- Galassetti, P.; Pontello, A. Dietary effects on oxidation of low-density lipoprotein and atherogenesis. Curr. Atheroscler. Rep. 2006, 8, 523–529. [Google Scholar] [CrossRef]
- Louheranta, A.M.; Sarkkinen, E.S.; Vidgren, H.M.; Schwab, U.S.; Uusitupa, M.I. Association of the fatty acid profile of serum lipids with glucose and insulin metabolism during 2 fat-modified diets in subjects with impaired glucose tolerance. Am. J. Clin. Nutr. 2002, 76, 331–337. [Google Scholar] [CrossRef][Green Version]
- Ros, E. Dietary cis-monounsaturated fatty acids and metabolic control in type 2 diabetes. Am. J. Clin. Nutr. 2003, 78, 617S–625S. [Google Scholar] [CrossRef]
- Due, A.; Larsen, T.M.; Hermansen, K.; Stender, S.; Holst, J.J.; Toubro, S.; Martinussen, T.; Astrup, A. Comparison of the effects on insulin resistance and glucose tolerance of 6-mo high-monounsaturated-fat, low-fat, and control diets. Am. J. Clin. Nutr. 2008, 87, 855–862. [Google Scholar] [CrossRef]
- Riserus, U.; Willett, W.C.; Hu, F.B. Dietary fats and prevention of type 2 diabetes. Prog. Lipid Res. 2009, 48, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Salar, A.; Faghih, S.; Pishdad, G.R. Rice bran oil and canola oil improve blood lipids compared to sunflower oil in women with type 2 diabetes: A randomized, single-blind, controlled trial. J. Clin. Lipidol. 2016, 10, 299–305. [Google Scholar] [CrossRef]
- Vessby, B.; Uusitupa, M.; Hermansen, K.; Riccardi, G.; Rivellese, A.A.; Tapsell, L.C.; Nälsén, C.; Berglund, L.; Louheranta, A.; Rasmussen, B.M.; et al. Substituting dietary saturated for monounsaturated fat impairs insulin sensitivity in healthy men and women: The KANWU Study. Diabetologia 2001, 44, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Adams-Huet, B.; Brinkley, L.; Grundy, S.M.; Garg, A. Lipid, glycemic, and insulin responses to meals rich in saturated, cis-monounsaturated, and polyunsaturated (n-3 and n-6) fatty acids in subjects with type 2 diabetes. Diabetes Care 2007, 30, 2993–2998. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Bonanome, A.; Grundy, S.M.; Zhang, Z.J.; Unger, R.H. Comparison of a high-carbohydrate diet with a high-monounsaturated-fat diet in patients with non-insulin-dependent diabetes-mellitus. N. Engl. J. Med. 1988, 319, 829–834. [Google Scholar] [CrossRef]
- Campbell, L.V.; Marmot, P.E.; Dyer, J.A.; Borkman, M.; Storlien, L.H. The high-monounsaturated fat diet as a practical alternative for NIDDM. Diabetes Care 1994, 17, 177–182. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J.; Nutrition Committee. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Arterioscler. Thromb. Vasc. Biol. 2003, 23, e20–e30. [Google Scholar] [CrossRef] [PubMed]
- Hardman, W.E. Dietary canola oil suppressed growth of implanted MDA-MB 231 human breast tumors in nude mice. Nutr. Cancer 2007, 57, 177–183. [Google Scholar] [CrossRef]
- Duda, M.K.; O’Shea, K.M.; Stanley, W.C. Omega-3 polyunsaturated fatty acid supplementation for the treatment of heart failure: Mechanisms and clinical potential. Cardiovasc. Res. 2009, 84, 33–41. [Google Scholar] [CrossRef]
- Fetterman, J.J.; Zdanowicz, M.M. Therapeutic potential of n-3 polyunsaturated fatty acids in disease. Am. J. Health-Syst. Pharm. 2009, 66, 1169–1179. [Google Scholar] [CrossRef]
- Hu, F.B.; Stampfer, M.J.; Manson, J.E.; Rimm, E.B.; Wolk, A.; Colditz, G.A.; Hennekens, C.H.; Willett, W.C. Dietary intake of alpha-linolenic acid and risk of fatal ischemic heart disease among women. Am. J. Clin. Nutr. 1999, 69, 890–897. [Google Scholar] [CrossRef]
- Albert, C.M.; Oh, K.; Whang, W.; Manson, J.E.; Chae, C.U.; Stampfer, M.J.; Willett, W.C.; Hu, F.B. Dietary alpha-linolenic acid intake and risk of sudden cardiac death and coronary heart disease. Circulation 2005, 112, 3232–3238. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Ascherio, A.; Hu, F.B.; Stampfer, M.J.; Willett, W.C.; Siscovick, D.S.; Rimm, E.B. Interplay between different polyunsaturated fatty acids and risk of coronary heart disease in men. Circulation 2005, 111, 157–164. [Google Scholar] [CrossRef]
- Sioen, I.; Hacquebard, M.; Hick, G.; Maindiaux, V.; Larondelle, Y.; Carpentier, Y.A.; De Henauw, S. Effect of ALA-enriched food supply on cardiovascular risk factors in males. Lipids 2009, 44, 603–611. [Google Scholar] [CrossRef]
- Hall, A.V.; Parbtani, A.; Clark, W.F.; Spanner, E.; Keeney, M.; Chin-Yee, I.; Philbrick, D.J.; Holub, B.J. Abrogation of MRL/lpr lupus nephritis by dietary flaxseed. Am. J. Kidney Dis. 1993, 22, 326–332. [Google Scholar] [CrossRef]
- Clark, W.F.; Parbtani, A.; Huff, M.W.; Spanner, E.; de Salis, H.; Chin-Yee, I.; Philbrick, D.J.; Holub, B.J. Flaxseed: A potential treatment for lupus nephritis. Kidney Int. 1995, 48, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, D.; Bankovic-Calic, N.; Peng, C.Y.; Ogborn, M.R.; Aukema, H.M. Dietary flax oil during pregnancy and lactation retards disease progression in rat off spring with inherited kidney disease. Pediatr. Res. 2006, 60, 729–733. [Google Scholar] [CrossRef][Green Version]
- Nguemeni, C.; Delplanque, B.; Rovère, C.; Simon-Rousseau, N.; Gandin, C.; Agnani, G.; Nahon, J.L.; Heurteaux, C.; Blondeau, N. Dietary supplementation of alpha-linolenic acid in an enriched rapeseed oil diet protects from stroke. Pharmacol. Res. 2010, 61, 226–233. [Google Scholar] [CrossRef]
- Whelan, J.; Rust, C. Innovative dietary sources of n-3 fatty acids. Ann. Rev. Nutr. 2006, 26, 75–103. [Google Scholar] [CrossRef] [PubMed]
- Rainer, L.; Heiss, C.J. Conjugated linoleic acid: Health implications and effects on body composition. J. Am. Diet. Assoc. 2004, 104, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Micallef, M.A.; Garg, M.L. Beyond blood lipids: Phytosterols, statins and omega-3 polyunsaturated fatty acid therapy for hyperlipidemia. J. Nutr. Biochem. 2009, 20, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, F.; Poli, A. Phytosterols and cardiovascular health. Pharmacol. Res. 2010, 61, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Gylling, H.; Simonen, P. Phytosterols, Phytostanols, and Lipoprotein Metabolism. Nutrients 2015, 7, 7965–7977. [Google Scholar] [CrossRef] [PubMed]
- Trautwein, E.A.; Koppenol, W.P.; de Jong, A.; Hiemstra, H.; Vermeer, M.A.; Noakes, M.; Luscombe-Marsh, N.D. Plant sterols lower LDL-cholesterol and triglycerides in dyslipidemic individuals with or at risk of developing type 2 diabetes; a randomized, double-blind, placebo-controlled study. Nutr. Diabetes 2018, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Woyengo, T.A.; Ramprasath, V.R.; Jones, P.J. Anticancer effects of phytosterols. Eur. J. Clin. Nutr. 2009, 63, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Pae, M.; Dao, M.C.; Smith, D.; Meydani, S.N.; Wu, D. Dietary Supplementation with Tocotrienols Enhances Immune Function in C57BL/6 Mice. J. Nutr. 2010, 140, 1335–1341. [Google Scholar] [CrossRef]
- Bhatia, E.; Doddivenaka, C.; Zhang, X.; Bommareddy, A.; Krishnan, P.; Matthees, D.P. Chemopreventive effects of dietary canola oil on colon cancer development. Nutr. Cancer 2011, 63, 242–247. [Google Scholar] [CrossRef]
- Stricker, H.; Duchini, F.; Facchini, M.; Mombelli, G. Canola oil decreases cholesterol and improves endothelial function in patients with peripheral arterial occlusive disease—A pilot study. Artery Res. 2008, 2, 67–73. [Google Scholar] [CrossRef]
- Palomaki, A.; Pohjantasahti-Maaroos, H.; Wallenius, M.; Kankkunen, P.; Aro, H.; Husgafvel, S.; Pihlava, J.M.; Oksanen, K. Effects of dietary cold-pressed turnip rapeseed oil and butter on serum lipids, oxidized LDL and arterial elasticity in men with metabolic syndrome. Lipids Health Dis. 2010, 9, 137. [Google Scholar] [CrossRef]
- Iggman, D.; Gustafsson, I.B.; Berglund, L.; Vessby, B.; Marckmann, P.; Riserus, U. Replacing dairy fat with rapeseed oil causes rapid improvement of hyperlipidaemia: A randomized controlled study. J. Intern. Med. 2011, 270, 356–364. [Google Scholar] [CrossRef]
- McLennan, P.L.; Dallimore, J.A. Dietary canola oil modifies myocardial fatty acids and inhibits cardiac arrhythmias in rats. J. Nutr. 1995, 125, 1003–1009. [Google Scholar] [CrossRef]
- FDA. Qualified Health Claims: Letter of Enforcement Discretion—Unsaturated Fatty Acids from Canola Oil and Reduced Risk of Coronary Heart Disease; Docket No. 2006Q-0091; U.S. Food and Drug Administration: Washington, DC, USA, 2009. Available online: http://www.fda.gov/Food/LabelingNutrition/LabelClaims/QualifiedHealthClaims/ucm072958.htm (accessed on 22 June 2019).
- Baudet, M.F.; Jacotot, B. Dietary fats and lecithin-cholesterol acyltransferase activity in healthy humans. Ann. Nutr. Metab. 1988, 32, 352–359. [Google Scholar] [CrossRef]
- Lichtenstein, A.H.; Ausman, L.M.; Carrasco, W.; Jenner, J.L.; Gualtieri, L.J.; Goldin, B.R.; Ordovas, J.M.; Schaefer, E.J. Effects of canola, corn, and olive oils on fasting and postprandial plasma lipoproteins in humans as part of a National Cholesterol Education Program Step 2 diet. Arterioscler. Thromb. 1993, 13, 1533–1542. [Google Scholar] [CrossRef]
- Matheson, B.; Walker, K.Z.; Taylor, D.M.; Peterkin, R.; Lugg, D.; O’Dea, K. Effect on serum lipids of monounsaturated oil and margarine in the diet of an Antarctic Expedition. Am. J. Clin. Nutr. 1996, 63, 933–938. [Google Scholar] [CrossRef]
- Sarkkinen, E.S.; Uusitupa, M.I.; Gylling, H.; Miettinen, T.A. Fat-modified diets influence serum concentrations of cholesterol precursors and plant sterols in hypercholesterolemic subjects. Metabolism 1998, 47, 744–750. [Google Scholar] [CrossRef]
- Becker, C.C.; Lund, P.; Holmer, G.; Jensen, H.; Sandstrom, B. Effects of butter oil blends withincreased concentrations of stearic, oleic and linolenic acid on blood lipids in young adults. Eur. J. Clin. Nutr. 1999, 53, 535–541. [Google Scholar] [CrossRef][Green Version]
- Kratz, M.; Cullen, P.; Kannenberg, F.; Kassner, A.; Fobker, M.; Abuja, P.M.; Assmann, G.; Wahrburg, U. Effects of dietary fatty acids on the composition and oxidizability of low-density lipoprotein. Eur. J. Clin. Nutr. 2002, 56, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Uusitupa, M.; Schwab, U.; Makimattila, S.; Karhapaa, P.; Sarkkinen, E.; Maliranta, H.; Agren, J.; Penttilä, I. Effects of two high-fat diets with different fatty acid compositions on glucose and lipid metabolism in healthy young women. Am. J. Clin. Nutr. 1994, 59, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.A.; Kendall, C.W.C.; Vuksan, V.; Faulkner, D.; Augustin, L.S.A.; Mitchell, S.; Ireland, C.; Srichaikul, K.; Mirrahimi, A.; Chiavaroli, L.; et al. Effect of Lowering the Glycemic Load With Canola Oil on Glycemic Control and Cardiovascular Risk Factors: A Randomized Controlled Trial. Diabetes Care 2014, 37, 1806–1814. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Kris-Etherton, P.M.; West, S.G.; Lamarche, B.; Jenkins, D.J.A.; Fleming, J.A. Effects of Canola and High-Oleic Acid Canola Oils on Abdominal Fat Mass in Individuals with Central Obesity. Obesity 2016, 24, 2261–2268. [Google Scholar] [CrossRef]
- Papazzo, A.; Conlan, X.; Lexis, L.; Lewandowski, P. The effect of short-term canola oil ingestion on oxidative stress in the vasculature of stroke-prone spontaneously hypertensive rats. Lipids Health Dis. 2011, 10, 180. [Google Scholar] [CrossRef]
- Sun, Y.; Magnussen, C.G.; Dwyer, T.; Oddy, W.H.; Venn, A.J.; Smith, K.J. Cross-Sectional Associations between Dietary Fat-Related Behaviors and Continuous Metabolic Syndrome Score among Young Australian Adults. Nutrients 2018, 10, 972. [Google Scholar] [CrossRef]
- Ramsden, C.E.; Zamora, D.; Leelarthaepin, B.; Majchrzak-Hong, S.F.; Faurot, K.R.; Suchindran, C.M. Use of dietary linoleic acid for secondary prevention of coronary heart disease and death: Evaluation of recovered data from the Sydney Diet Heart Study and updated meta-analysis. BMJ 2013, 346, e8707. [Google Scholar] [CrossRef] [PubMed]
- Hamley, S. The effect of replacing saturated fat with mostly n-6 polyunsaturated fat on coronary heart disease: A meta-analysis of randomised controlled trials. Nutr. J. 2017, 16, 30. [Google Scholar] [CrossRef] [PubMed]
- Mboma, J.; Leblanc, N.; Angers, P.; Rocher, A.; Vigor, C.; Oger, C. Effects of Cyclic Fatty Acid Monomers from Heated Vegetable Oil on Markers of Inflammation and Oxidative Stress in Male Wistar Rats. J. Agric. Food Chem. 2018, 66, 7172–7180. [Google Scholar] [CrossRef]
- Przybylski, R.; Eskin, N.A.M. Oil Composition and Properties. In Canola; Elsevier: Amsterdam, The Netherlands, 2011; pp. 189–227. [Google Scholar] [CrossRef]
- Przybylski, R.; Gruczynska, E.; Aladedunye, F. Performance of regular and modified canola and soybean oils in rotational frying. J. Am. Oil Chem. Soc. 2013, 90, 1271–1280. [Google Scholar] [CrossRef]
- Roiaini, M.; Ardiannie, T.; Norhayati, H. Physicochemical properties of canola oil, olive oil and palm olein blends. Int. Food Res. J. 2015, 22, 1228–1234. [Google Scholar]
- Peterson, C.L.; Reece, D.L.; Hammond, B.L.; Thompson, J.; Beck, S.M. Processing, characterization, and performance of eight fuels from lipids. Appl. Eng. Agric. 1997, 13, 71–79. [Google Scholar] [CrossRef]
- Boland, M. Rapeseed. Agricultural Marketing Resource Center. 2012. Available online: http://www.agmrc.org/commodities__products/grains__oilseeds/rapeseed (accessed on 17 October 2014).
- Iveta, Z.; Eva, C. The utilization of rapeseed for biofuels production in the EU. Visegr. J. Bioecon. Sustain. Dev. 2013, 1, 11–14. [Google Scholar] [CrossRef]
- Hamelinck, C.; Koper, M.; Berndes, G.; Englund, O.; Diaz-Chavez, R.; Kunen, E.; Walden, D. Biofuels Baseline 2008; The Netherlands. 2011. Available online: http://www.ecofys.com/files/files/ecofys_2011_biofuels_baseline(2008).pdf (accessed on 19 August 2019).
- Pahl, G. Biodiesel: Growing a New Energy Economy; Chelsea Green Publishing Company: Chelsea, VT, USA, 2008. [Google Scholar]
- Aukema, H.; Campbell, L. Oil nutrition and utilization. In Canola; Elsevier: Amsterdam, The Netherlands, 2011; pp. 245–280. [Google Scholar] [CrossRef]
- O’Connor, D. Lifecycle Analysis Canola Biodiesel; (S&T) 2 Consultants Inc.: Winnipeg, MB, Canada, 2011; p. 22. Available online: http://www.canolacouncil.org/media/504977/don_o_connor.pdf (accessed on 22 August 2019).
- Finco, A.; Bentivoglio, D.; Nijkamp, P. Integrated evaluation of biofuel production options in agriculture: An exploration of sustainable policy scenarios. Int. J. Foresight Innov. Policy 2012, 8, 173–188. [Google Scholar] [CrossRef]
- Wolinetz, M.; Hein, M. Biofuels in Canada 2017, Tracking Biofuel Consumption, Feedstocks and Avoided Greenhouse Gas Emissions; Navius Research Inc.: Vancouver, BC, Canada, 2017; Volume 15, p. 22. [Google Scholar]
- CRFA. Environmental Benefits of Biodiesel; Canadian Renewable Fuels Association: Ottawa, ON, Canada, 2010; Available online: http://www.greenfuels.org (accessed on 20 July 2019).
- Health Canada. Human Health Risk Assessment for Biodiesel Production, Distribution and Use in Canada. Available online: https://www.canada.ca/en/health-canada/services/publications/healthy-living/human-health-risk-assessment-biodiesel-production-distribution-use-canada.html (accessed on 22 September 2019).
- Bio Cube Corporation. Available online: https://biocubeco.com/faqs/will-biodiesel-damage-seals-and-other-components/ (accessed on 28 June 2019).
- Zelmer, C.D.; McVetty, P.B.E. Industrial Products. In Biology and Breeding of Crucifers; Gupta, S.K., Ed.; CRC Press: Boca Raton, FL, USA, 2009; pp. 343–360. [Google Scholar]
- Sonntag, N.O.V. Separation of Fatty Acids. In Fatty Acids; Pryde, E.H., Ed.; AOCS Press: Champaign, IL, USA, 1979; pp. 125–156. [Google Scholar]
- Leonard, C. Sources and commercial applications of high erucic vegetable oils. Lipid Technol. 1994, 6, 79–83. [Google Scholar]
- Senthilselvan, A.; Zhang, Y.; Dosman, J.A.; Barber, E.M.; Holfeld, L.E.; Kirychuk, S.P.; Cormier, Y.; Hurst, T.S.; Rhodes, C.S. Positive human health effects of dust suppression with canola oil in swine barns. Am. J. Respir. Crit. Care Med. 1997, 156, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Frolicious. Benefits of Omega 3, 6, 9 Fatty Acids for Hair. Available online: https://www.frolicious.de/2016/01/31/benefits-of-omega-3-6-9-fatty-acids-for-hair/ (accessed on 15 November 2019).
- Lush Retail Ltd. Rapeseed Oil; Coconut Oil. Available online: https://www.lush.com/uk/en/i/rapeseed-oil-coconut-oil (accessed on 10 June 2020).
- Biopesticides and Pollution Prevention Division. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/registration/fs_PC-011332_25-Nov-09.pdf (accessed on 30 October 2019).
- Thurston County Health Department. Rapeseed Oil. Available online: https://www.co.thurston.wa.us/health/ehipm/pdf_insect/insecticide%20actives/rapeseedOil.pdf (accessed on 7 May 2019).
- Arntfield, S.D.; Hickling, D. Meal Nutrition and Utilization. In Canola; Elsevier: Amsterdam, The Netherlands, 2011; pp. 281–312. [Google Scholar] [CrossRef]
- Newkirk, R. Meal Nutrient Composition. In Canola; Elsevier: Amsterdam, The Netherlands, 2011; pp. 229–244. [Google Scholar] [CrossRef]
- Barthet, V.J.; Daun, J.K. Seed Morphology, Composition, and Quality. In Canola; Elsevier: Amsterdam, The Netherlands, 2011; pp. 119–163. [Google Scholar] [CrossRef]
- Downey, R.K.; Craig, B.M.; Young, C.G. Breeding rapeseed for oil and meal quality. JAOCS 1969, 46, 121. [Google Scholar] [CrossRef]
- Stefansson, B.R. Rapeseed breeding in Canada. Chapter 4. In The Story of Rapeseed in Western Canada; Saskatchewan Wheat Pool: Regina, SK, Canada, 1974. [Google Scholar]
- Clandinin, D.R. Rapeseed oil meal studies: 4. Effect of sinapine, the bitter substance in rapeseed oil meal, on the growth of chickens. Poult. Sci. 1961, 40, 484–487. [Google Scholar] [CrossRef]
- Zhu, L.P.; Wang, J.P.; Ding, X.M.; Bai, S.P.; Zeng, Q.F.; Su, Z.W.; Xuan, Y.; Applegate, T.J.; Zhang, K.Y. The effects of varieties and levels of rapeseed expeller cake on egg production performance, egg quality, nutrient digestibility, and duodenum morphology in laying hens. Poult. Sci. 2019, 98, 4942–4953. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Pan, J.; Xie, R.; Qin, J.; He, R.; Ma, H. Screening of strains used for producing rapeseed peptide and degradation of glucosinolates by solid-state fermentation. Sci. Technol. Food Ind. 2015, 36, 164–168. [Google Scholar] [CrossRef]
- Sun, L.; Liu, P.; Zhuo, W.; Fang, X.; Fan, H.; Wu, H. Production of high protein and polypeptide rapeseed meal by microbial fermentation. China Oils Fats 2017, 42, 94–98. [Google Scholar]
- Drazbo, A.; Kozlowski, K.; Ognik, K.; Zaworska, A.; Jankowski, J. The effect of raw and fermented rapeseed cake on growth performance, carcass traits, and breast meat quality in turkey. Poult. Sci. 2019, 98, 6161–6169. [Google Scholar] [CrossRef] [PubMed]
- Tie, Y.; Li, L.; Liu, J.; Liu, C.L.; Fu, J.J.; Xiao, X.J.; Wang, G.; Wang, J. Two-step biological approach for treatment of rapeseed meal. J. Food Sci. 2020, 85, 340–348. [Google Scholar] [CrossRef]
- Hansen, J.O.; Overland, M.; Skrede, A.; Anderson, D.M.; Collins, S.A. A meta-analysis of the effects of dietary canola/double low rapeseed meal on growth performance of weanling and growing-finishing pigs. Anim. Feed Sci. Technol. 2020, 259, 114302. [Google Scholar] [CrossRef]
- Pastuszewska, P.; Ochtabinska, A.; Morawski, A. A note on the nutritional adequacy of stock diets for laboratory rats and mice. J. Anim. Feed Sci. 2000, 9, 533–542. [Google Scholar] [CrossRef]
- Aider, M.; Barbana, C. Canola proteins: Composition, extraction, functional properties, bioactivity, applications as a food ingredient and allergenicity—A practical and critical review. Trends Food Sci. Technol. 2011, 22, 21–39. [Google Scholar] [CrossRef]
- Tan, S.H.; Mailer, R.J.; Blanchard, C.L.; Agboola, S.O. Canola proteins for human consumption: Extraction, profile, and functional properties. J. Food Sci. 2011, 76, R16–R28. [Google Scholar] [CrossRef] [PubMed]
- Fu, T. Rapeseed quality improvement. Crop Res. 2007, 21, 159–162. [Google Scholar]
- Khattab, R.Y.; Arntfield, S.D. Functional properties of raw and processed canola meal. LWT Food Sci. Technol. 2009, 42, 1119–1124. [Google Scholar] [CrossRef]
- Uruakpa, F.O.; Arntfield, S.D. Rheological characteristics of commercial canola protein isolate-κ carrageenan systems. Food Hydrocoll. 2004, 18, 419–427. [Google Scholar] [CrossRef]
- Vioque, J.; Sanchez-Vioque, R.; Clements, A.; Pedroche, J.; Millan, F. Partially hydrolyzed rapeseed protein isolates with improved functional properties. J. Am. Oil Chem. Soc. 2000, 77, 447–450. [Google Scholar] [CrossRef]
- Pinterits, A.; Arntfield, S.D. The effect of limited proteolysis on canola protein gelation. Food Chem. 2007, 102, 1337–1343. [Google Scholar] [CrossRef]
- Wu, J.; Muir, A.D. Comparative structural, emulsifying, and biological properties of 2 major canola proteins, cruciferin and napin. J. Food Sci. 2008, 73, C210–C216. [Google Scholar] [CrossRef] [PubMed]
- Yoshie-Stark, Y.; Wada, Y.; Wasche, A. Chemical composition, functional properties, and bioactivities of rapeseed protein isolates. Food Chem. 2008, 107, 32–39. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Muir, A.D. Production of angiotensin I-converting enzyme inhibitory peptides from defatted canola meal. Bioresour. Technol. 2009, 100, 5283–5287. [Google Scholar] [CrossRef]
- Cumby, N.; Zhong, Y.; Haczk, M.; Shahidi, R. Antioxidant activity and water-holding capacity of canola protein hydrolysates. Food Chem. 2008, 109, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Marczak, E.D.; Usui, H.; Fujita, H.; Yang, Y.; Yokoo, M.; Lipkowski, A.W.; Yoshikawa, M. New antihypertensive peptides isolated from rapeseed. Peptides 2003, 24, 791–798. [Google Scholar] [CrossRef]
- Chabanon, G.; Alves da Costa, L.; Farges, B.; Harscoat, C.; Chenu, S.; Goergen, J.L.; Marc, A.; Marc, I.; Chevalot, I. Influence of the rapeseed hydrolysis process on CHO cell growth. Bioresour. Technol. 2008, 99, 7143–7151. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Tian, S.; Small, D.M. Generation of meat-like flavourings from enzymatic hydrolysates of proteins from Brassica sp. Food Chem. 2010, 119, 167–172. [Google Scholar] [CrossRef]
- Burcon. Canola Protein. Available online: https://burcon.live-website.com/products/canola-proteins/ (accessed on 6 July 2021).
- Schweizer, M.; Segall, K.; Medina, S.; Willardsen, R.; Tergesen, J. Rapeseed/canola protein isolates for use in the food industry. Qual. Nutr. Process. Process. Technol. 2007, 160–162. [Google Scholar]
- Helm, A.G. Safety Assessment of Rapeseed Protein (IsolexxTM); Food Safety Authority of Ireland: Dublin, Ireland, 2012; pp. 1–7. Available online: https://www.fsai.ie/uploadedFiles/Science_and_Health/Novel_Foods/Applications/2012%20Rapeseed%20protein.pdf (accessed on 22 July 2019).
- Tan, S.H.; Mailer, R.J.; Blanchard, C.L.; Agboola, S.O. Emulsifying properties of proteins extracted from Australian canola meal. LWT-Food Sci. Technol. 2014, 57, 376–382. [Google Scholar] [CrossRef]
- European Food Safety Authority Panel. Scientific Opinion on the safety of “rapeseed protein isolate” as a Novel Food ingredient. EFSA Panel Diet. Prod. Nutr. Allerg. NDA 2013, 11, 3420. [Google Scholar] [CrossRef]
- Petit, H.V.; Veira, D.M. Effect of post-weaning protein supplementation of beef steers fed grass silage on performance during the finishing phase, and carcass quality. Can. J. Anim. Sci. 1994, 74, 699–701. [Google Scholar] [CrossRef]
- Patterson, H.H.; Whittier, J.C.; Rittenhouse, L.R.; Schutz, D.N. Performance of beef cows receiving cull beans, sunflower meal, and canola meal as protein supplements while grazing native winter range in Eastern Colorado. J. Anim. Sci. 1999, 77, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.Z.; Xu, L.; Li, C.; Beauchemin, K.A. Effects of supplemental canola meal and various types of distillers’ grains on growth performance of backgrounded steers. Can. J. Anim. Sci. 2013, 93, 281–286. [Google Scholar] [CrossRef]
- Paula, E.M.; Broderick, G.A.; Faciola, A.P. Effects of replacing soybean meal with canola meal for lactating dairy cows fed 3 different ratios of alfalfa to corn silage. J. Dairy Sci. 2020, 103, 1463–1471. [Google Scholar] [CrossRef] [PubMed]
- Damiran, D.; McKinnon, J.J. Evaluation of wheat-based dried distillers grains with solubles or canola meal derived from Brassica napus seed as an energy source for feedlot steers. Transl. Anim. Sci. 2018, 2 (Suppl. S1), S139–S144. [Google Scholar] [CrossRef] [PubMed]
- Huhtanen, P.; Hetta, M.; Swensson, C. Evaluation of canola meal as a protein supplement for dairy cows: A review and a meta-analysis. Can. J. Anim. Sci. 2011, 91, 529–543. [Google Scholar] [CrossRef]
- Martineau, R.; Ouellet, D.R.; Lapierre, H. Feeding canola meal to dairy cows: A meta-analysis on lactational responses. J. Dairy Sci. 2013, 96, 1701–1714. [Google Scholar] [CrossRef]
- Chibisa, G.E.; Christensen, D.A.; Mutsvangwa, T. Replacing canola meal as the major protein source with wheat dried distillers’ grains alters omasal fatty acid flow and milk fatty acid composition in dairy cows. Can. J. Anim. Sci. 2013, 93, 137–147. [Google Scholar] [CrossRef][Green Version]
- Auldist, M.J.; Marett, L.C.; Greenwood, J.S.; Wright, M.M.; Hannah, M.; Jacobs, J.L.; Wales, W.J. Replacing wheat with canola meal in a partial mixed ration increases the milk production of cows grazing at a restricted pasture allowance in spring. Anim. Prod. Sci. 2014, 54, 869–878. [Google Scholar] [CrossRef]
- Brand, T.S.; Brandt, D.A.; Cruywagen, C.W. Utilisation of growing-finishing pig diets containing high levels of solvent or expeller oil extracted canola meal. N. Z. J. Agric. Res. 2001, 44, 31–35. [Google Scholar] [CrossRef][Green Version]
- Schone, F.; Rudolph, B.; Kirchheim, U.; Knapp, G. Counteracting the negative effects of rapeseed and rapeseed press cake in pig diets. Br. J. Nutr. 1997, 78, 947–962. [Google Scholar] [CrossRef]
- Do, S.H.; Kim, B.O.; Fang, L.H.; You, D.H.; Hong, J.S.; Kim, Y.Y. Various levels of rapeseed meal in weaning pig diets from weaning to finishing periods. AJAS 2017, 30, 1292–1302. [Google Scholar] [CrossRef]
- Lee, J.W.; Woyengo, T.A. Growth performance, organ weights, and blood parameters of nursery pigs fed diets containing increasing levels of cold-pressed canola cake. J. Anim. Sci. 2018, 96, 4704–4712. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.Q.; Huang, C.F.; Li, Y.K.; Li, P.L.; Liu, H.; Chen, Y.F.; Li, D.F.; Lai, C.H. Adaptation duration for net energy determination of high fiber diets in growing pigs. Anim. Feed Sci. Technol. 2018, 241, 15–26. [Google Scholar] [CrossRef]
- Mejicanos, G.A.; Kim, J.W.; Nyachoti, C.M. Tail-end dehulling of canola meal improves apparent and total tract digestibility of phosphorus when fed to growing pigs. J. Anim. Sci. 2018, 96, 1430–1440. [Google Scholar] [CrossRef] [PubMed]
- Velayudhan, D.E.; Hossain, M.M.; Regassa, A.; Nyachoti, C.M. Effect of canola meal inclusion as a major protein source in gestation and lactation sow diets with or without enzymes on reproductive performance, milk composition, fecal bacterial profile and nutrient digestibility. Anim. Feed Sci. Technol. 2018, 241, 141–150. [Google Scholar] [CrossRef]
- Vincent, I.C.; Williams, H.L.; Hill, R. Feeding British rapeseed meals to pregnant and lactating ewes. Anim. Prod. 1988, 47, 283–289. [Google Scholar] [CrossRef]
- Vincent, I.C.; Thompson, J.; Hill, R. The voluntary feed intake and weight gain of lambs given concentrate feeds containing rapeseed meal with a range of glucosinolate contents. Anim. Prod. 1990, 50, 587. [Google Scholar] [CrossRef]
- Hill, R. Rapeseed meal in the diets of ruminants. Nutr. Abs. Rev. B 1991, 61, 139–155. [Google Scholar]
- Reis, P.J.; Tunks, D.A.; Munro, S.G. Effects of the infusion of amino acids into the abomasum of sheep, with emphasis on the relative value of methionine, cysteine and homocysteine for wool growth. J. Agric. Sci. 1990, 114, 59–68. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; Sykes, J.M.; Bignell, W.C. Influence of lupins and canola supplements on plasma amino acids, wool fibre diameter and liveweight in genetically divergent first cross Merino lambs. In Proceedings of the World Congress on Fats and Oils, Sydney, Australia, 27–30 September 2009. [Google Scholar]
- Sutton, E.I. Canola Meal for Other Species. Horses. Feed Intake and Performance. In Canola Nutritionist Manual; Canola Council of Canada: Winnipeg, MB, Canada, 1988. [Google Scholar]
- Lebas, F.; Colin, M. Utilization of rapeseed oil meal in growing rabbit feeding. Effect of dehulling. Ann. Zootech. 1977, 26, 93–97. [Google Scholar] [CrossRef]
- Brand, T.S.; De Brabander, L.; Van Schalkwyk, S.J.; Pfister, B.; Hays, J.P. The true metabolisable energy content of canola oilcake meal and full-fat rapeseed seed for ostriches (Struthio camelus). Br. Poult. Sci. 2000, 41, 201–203. [Google Scholar] [CrossRef]
- Badshah, A.; Aurang, Z.; Nizakat, B.; Sajjad, A.; Chaudry, M.A.; Sattar, A. Utilization of rapeseed meal/cake in poultry feed. Part II. Effect of incorporating higher levels of rapeseed cake in poultry diet on laying performance of brown-egg layer. Pak. J. Sci. Ind. Res. 2001, 44, 171–174. [Google Scholar]
- Marcu, N.; Banto, E.; Sut-Gherman, M.; Dinea, M.; Ludu, O.; Ceghezi, J. The effect of soybean meal substitution with rape meal in laying hens nutrition. Bull. Univ. Stiint. Agric. Med. Vet. Cluj-Napoca Ser. Zooteh. Biotehnol. 2004, 60, 138–142. [Google Scholar]
- Perez-Maldonado, R.A.; Barram, K.M. Evaluation of Australian Canola Meal for Production and Egg Quality in Two Layer Strains. In Proceedings of the 16th Australian Poultry Science Symposium, Sydney, Australia, 14–16 February 2004; pp. 171–174. [Google Scholar]
- Newkirk, R.W.; Classen, H.L. The effects of toasting canola meal on body weight, feed conversion efficiency, and mortality in broiler chickens. Poult. Sci. 2002, 81, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, K.R.; Devegowda, G.; Khosravinia, H. Effects of enzyme addition to broiler diets containing varying levels of double zero rapeseed meal. Asian-Aust. J. Anim. Sci. 2006, 19, 1354–1360. [Google Scholar] [CrossRef]
- Manyeula, F.; Mlambo, V.; Marume, U.; Sebola, N.A. Nutrient digestibility, haemo-biochemical parameters and growth performance of an indigenous chicken strain fed canola meal-containing diets. Trop. Anim. Health Prod. 2019, 51, 2343–2350. [Google Scholar] [CrossRef]
- Manyeula, F.; Mlambo, V.; Marume, U.; Sebola, N.A. Partial replacement of soybean products with canola meal in indigenous chicken diets: Size of internal organs, carcass characteristics and breast meat quality. Poult. Sci. 2020, 99, 256–262. [Google Scholar] [CrossRef]
- Mnisi, C.M.; Mlambo, V. Growth performance, haematology, serum biochemistry and meat quality characteristics of Japanese quail (Coturnix coturnix japonica) fed canola meal-based diets. Anim. Nutr. 2018, 4, 37–43. [Google Scholar] [CrossRef]
- Mnisi, C.M.; Mlambo, V. Protease treatment of canola meal-containing Japanese quail diets: Effect on physiological parameters and meat quality traits. J. Appl. Anim. Res. 2018, 46, 1389–1394. [Google Scholar] [CrossRef]
- Drażbo, A.; Ognik, K.; Zaworska, A.; Ferenc, K.; Jankowski, J. The effect of raw and fermented rapeseed cake on the metabolic parameters, immune status, and intestinal morphology of turkeys. Poult. Sci. 2018, 97, 3910–3920. [Google Scholar] [CrossRef]
- Rad-Spicea, M.; Rogiewicza, A.; Jankowskib, J.; Slominski, B.A. Yellow-seeded B. napus and B. juncea canola. Part 1. Nutritive value of the meal for broiler chickens. Anim. Feed Sci. Technol. 2018, 240, 66–77. [Google Scholar] [CrossRef]
- Kozlowskia, K.; Mikulskia, D.; Rogiewiczc, A.; Zdunczykb, Z.; Rad-Spicec, M.; Jeroch, H.; Jankowskia, J.; Slominskic, B.A. Yellow-seeded B. napus and B. juncea canola. Part 2. Nutritive value of the meal for turkeys. Anim. Feed Sci. Technol. 2018, 240, 102–116. [Google Scholar] [CrossRef]
- Bell, J.M. Factors affecting the nutritional value of canola meal: A review. Can. J. Anim. Sci. 1993, 73, 679–697. [Google Scholar] [CrossRef]
- Jiang, Y.; Deyholos, M.K. Transcriptome analysis of secondary-wall-enriched seed coat tissues of canola (Brassica napus L.). Plant Cell Rep. 2010, 29, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Higgs, D.A.; Dosanjh, B.S.; Little, M.; Roy, R.J.J.; McBride, J.R. Potential for including Canola Products (Meal and Oil) in Diets for Oreochromis mossambicus × O. aureus Hybrids. In Proceedings of the Third International Symposium on Feeding and Nutrition in Fish, Toba, Japan, 28 August–1 September 1989; pp. 301–314. [Google Scholar]
- Lim, C.; Beames, R.M.; Eales, J.G.; Prendergast, A.F.; McLeese, J.M.; Shearer, K.D.; Higgs, D.A. Nutritive values of low and high fibre canola meals for shrimp. Aquacult. Nutr. 1997, 3, 269–279. [Google Scholar] [CrossRef]
- Abdul-Aziz, G.M.; El-Nady, M.A.; Shalaby, A.S.; Mahmoud, S.H. Partial substitution of soybean meal protein by different plant protein sources in diets for Nile tilapia fingerlings. Bull. Fac. Agric. Univ. Cairo 1999, 50, 189–202. [Google Scholar] [CrossRef]
- Glencross, B. Pilot Assessment of the Potential for Canola Meal and Oil Use in Aquaculture Feeds; Final Report for the Grains Research and Development Corporation. Fisheries Research Contract Report No. 5; Department of Fisheries, Western Australia: North Beach, Australia, 2003. [Google Scholar]
- Xu, W.J.; Jin, J.Y.; Zou, T.; Han, D.; Liu, H.K.; Zhu, X.M. Growth, feed utilization and metabolic responses of three gibel carp (Carassius gibelio) strains to fishmeal and plant protein-based diets. Aquac. Nutr. 2018, 25, 319–332. [Google Scholar] [CrossRef]
- Von Danwitz, A.; Schulz, C. Effects of dietary rapeseed glucosinolates, sinapic acid and phytic acid on feed intake, growth performance and fish health in turbot (Psetta maxima L.). Aquaculture 2020, 516, 734624. [Google Scholar] [CrossRef]
- Lim, C.; Klesius, P.H.; Higgs, D.A. Substitution of canoila meal for soybean meal in diets for channel catfish, Ictalurus punctatus. J. World Aquac. Soc. 1998, 29, 161–168. [Google Scholar] [CrossRef]
- Bandara, N.; Akbari, A.; Esparza, Y.; Wu, J. Canola Protein: A Promising Protein Source for Delivery, Adhesive, and Material Applications. J. Am. Oil Chem. Soc. 2018, 95, 1075–1090. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Q.; Rempel, C. Processing and characteristics of canola protein-based biodegradable packaging: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 475–485. [Google Scholar] [CrossRef]
- Delgado, M.; Felix, M.; Bengoechea, C. Development of bioplastic materials: From rapeseed oil industry by products to added-value biodegradable biocomposite materials. Ind. Crops Prod. 2018, 125, 401–407. [Google Scholar] [CrossRef]
- Li, S.; Ball, B.; Donner, E.; Thompson, M.R.; Rempel, C.; Liu, Q. Mechanical properties of green canola meal composites and reinforcement with cellulose fibers. Polym. Bull. 2019, 76, 1257–1275. [Google Scholar] [CrossRef]
- Azargohar, R.; Soleimani, M.; Nosran, S.; Bond, T.; Karunakaran, C.; Dalai, A.K.; Tabil, L.G. Thermo-physical characterization of torrefied fuel pellet from co-pelletization of canola hulls and meal. Ind. Crops Prod. 2019, 128, 424–435. [Google Scholar] [CrossRef]
- Rivera, D.; Rommi, K.; Fernandes, M.M.; Lantto, R.; Tzanov, T. Biocompounds from rapeseed oil industry co-stream as active ingredients for skin care applications. Int. J. Cosmet. Sci. 2015, 37, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, P.; Shi, H.; Zhou, Y.; Huang, H.; Tang, S. Preliminary results of yield, income and nutrition of double low rapeseed (Brassica napus L.) cultivated for oilseed vegetable-dual-purpose. Chin. Agric. Sci. Bull. 2009, 25, 224–227. [Google Scholar]
- Martinez, S.; Losada, P.; Franco, I.; Carballo, J. Protein, amino acid, ash and mineral contents in Brassica spp. Grown in northwest Spain. Int. J. Food Sci. Technol. 2010, 46, 146–153. [Google Scholar] [CrossRef]
- Van Duyn, M.A.; Pivonka, E. Overview of health benefits of fruit and vegetable consumption for the dietetics professional. Selective literature. J. Am. Diet. Assoc. 2000, 100, 1511–1521. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Hecker, K.D.; Bonanome, A.; Coval, S.M.; Binkoski, A.E.; Hilpert, K.F.; Griel, A.E.; Etherton, T.D. Bioactive compounds in foods: Their role in the prevention of cardiovascular disease and cancer. Am. J. Med. 2002, 113, 71–88. [Google Scholar] [CrossRef]
- Akubugwo, I.E.; Obasi, N.A.; Chinyere, G.C.; Ugbogu, A.E. Nutritional and chemical value of Amaranthus hybridus L. leaves from Nigeria. Afr. J. Biotechnol. 2007, 6, 2833–2839. [Google Scholar] [CrossRef]
- Boffetta, P.; Couto, E.; Wichmann, J.; Ferrari, P.; Trichopoulos, D.; Bueno-de-Mesquita, H.B.; van Duijnhoven, F.J.; Büchner, F.L.; Key, T.; Boeing, H.; et al. Fruit and vegetable intake and overall cancer risk in the european prospective investigation into cancer and nutrition (EPIC). JNCI-J. Natl. Cancer Inst. 2010, 102, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K. Vegetables and Fruit in the Prevention of Chronic Age-Related Diseases. In Molecular Basis of Nutrition and Aging; Molecular Nutrition; Academic Press: Cambridge, MA, USA, 2016; pp. 707–722. [Google Scholar] [CrossRef]
- Neugart, S.; Baldermann, S.; Ngwene, B.; Wesonga, J.; Schreiner, M. Indigenous leafy vegetables of Eastern Africa—A source of extraordinary secondary plant metabolites. Food Res. Int. 2017, 100, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Perles, R.; Moreno, D.A.; García-Viguera, C. Analysis of the tumoral cytotoxicity of green tea-infusions enriched with broccoli. Food Chem. 2012, 132, 1197–1206. [Google Scholar] [CrossRef]
- Michels, A.J.; Frei, B.; Vitamin, C. Biochemical, Physiological, and Molecular Aspects of Human Nutrition; Stipanuk, M.H., Caudill, M.A., Eds.; Elsevier/Saunders: St. Louis, MO, USA, 2012; pp. 626–654. [Google Scholar] [CrossRef]
- Naderi, N.; House, J.D. Chapter Five—Recent Developments in Folate Nutrition. Adv. Food Nutr. Res. 2018, 83, 195–213. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, N.; Wang, C.; Yu, G. Research on functional properties and modification of dietary fiber. Grain Process. 2010, 35, 57–59. [Google Scholar]
- Khattab, R.; Goldberg, E.; Lin, L.; Thiyam, U. Quantitative analysis and free-radical-scavenging activity of chlorophyll, phytic acid, and condensed tannins in canola. Food Chem. 2010, 122, 1266–1272. [Google Scholar] [CrossRef]
- Ling, W.H.; Jones, P.J.H. Dietary phytosterols: A review of metabolism, benefits, and side effects. Life Sci. 1995, 57, 195–206. [Google Scholar] [CrossRef]
- Venkatramesh, M.; Karunanandaa, B.; Sun, B.; Grunter, C.A.; Boddupalli, S.; Kishore, G.M. Expression of a streptomyces 3-hydroxysteroid oxidase gene in oilseeds for converting phytosterols to phytostanols. Phytochemistry 2003, 62, 39–46. [Google Scholar] [CrossRef]
- Verhoeven, D.T.; Verhagen, H.; Goldbohm, R.A.; van den Brandt, P.A.; van Poppel, G. A review of mechanisms underlying anticarcinogenicity by brassica vegetables. Chem. Biol. Interact. 1997, 103, 79–129. [Google Scholar] [CrossRef]
- Herr, I.; Büchler, M.W. Dietary constituents of broccoli and other cruciferous vegetables: Implications for prevention and therapy of cancer. Cancer Treat. Rev. 2010, 36, 377–382. [Google Scholar] [CrossRef]
- Yu, X.; Xu, X.; Teng, G. Inhibition effect of glucosinolates on tumour cells. Sci. Technol. Food Ind. 2010, 1, 165–167. [Google Scholar]
- Avato, P.; Argentieri, M.P. Brassicaceae: A rich source of health improving phytochemicals. Phytochem. Rev. 2015, 14, 1019–1033. [Google Scholar] [CrossRef]
- Jeffery, E.H. Cancer prevention with Brassica vegetables. Acta Hortic. 2014, 1106, 143–146. [Google Scholar] [CrossRef]
- Traka, M.H. Health Benefits of Glucosinolates. In Advances in Botanical Research; Elsevier Ltd.: Amsterdam, The Netherlands, 2016; Volume 80, pp. 247–279. [Google Scholar] [CrossRef]
- Ho, C.L.; Tan, H.Q.; Chua, K.J.; Kang, A.; Lim, K.H.; Ling, K.L.; Yew, W.S.; Lee, Y.S.; Thiery, J.P.; Chang, M.W. Engineered commensal microbes for diet-mediated colorectal-cancer chemoprevention. Nat. Biomed. Eng. 2018, 2, 27–37. [Google Scholar] [CrossRef]
- Mawson, R.; Heaney, R.K.; Zdunczyk, Z.; Kozlowska, H. Rapeseed meal-glucosinolates and their antinutritional effects 2. Flavor and palatability. Nahrung-Food 1993, 37, 336–344. [Google Scholar] [CrossRef]
- Borgen, B.H.; Tangstad, O.P.; Ahuja, I.; Rossiter, J.T.; Bones, A.M. Removing the mustard oil bomb from seeds: Transgenic ablation of myrosin cells in oilseed rape (Brassica napus) produces mineless seeds. J. Exp. Bot. 2010, 61, 1683–1697. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.; Williamson, G. A critical review of the bioavailability of glucosinolates and related compounds. Nat. Prod. Rep. 2004, 21, 425–447. [Google Scholar] [CrossRef] [PubMed]
- Herman-Antosiewicz, A.; Johnson, D.E.; Singh, S.V. Sulforaphane causes autophagy to inhibit release of cytochrome C and apoptosis in human prostate cancer cells. Cancer Res. 2006, 66, 5828–5835. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Shen, L.; Mao, J.; Huang, G.; Yuan, H. Study on extraction of sulforaphane from broccoli. J. Food Sci. Biotechnol. 2009, 28, 647–652. [Google Scholar]
- Kalpana, D.P.D.; Gayathri, R.; Gunassekaran, G.R.; Murugan, S.; Sakthisekaran, D. Apoptotic role of natural isothiocyanate from broccoli (Brassica oleracea italica) in experimental chemical lung carcinogenesis. Pharm. Biol. 2013, 51, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, A.; Abdull Razis, A.F. Apoptosis as a Mechanism of the Cancer Chemopreventive Activity of Glucosinolates: A Review. Asian Pac. J. Cancer Prev. 2018, 19, 1439–1448. [Google Scholar] [CrossRef]
- Lampe, J.W.; Peterson, S. Brassica, biotransformation and cancer risk: Genetic polymorphisms alter the preventive effects of cruciferous vegetables. J. Nutr. 2002, 132, 2991–2994. [Google Scholar] [CrossRef]
- Hwang, E.S.; Jeffery, E.H. Evaluation of urinary N-acetyl cysteinyl allyl isothiocyanate as a biomarker for intake and bioactivity of Brussels sprouts. Food Chem. Toxicol. 2003, 41, 1817–1825. [Google Scholar] [CrossRef]
- Ramirez, M.C.; Singletary, K. Regulation of estrogen receptor alpha expression in human breast cancer cells by sulforaphane. J. Nutr. Biochem. 2009, 20, 195–200. [Google Scholar] [CrossRef]
- Pawlik, A.; Wiczk, A.; Kaczynska, A.; Antosiewicz, J.; Herman Antosiewicz, A. Sulforaphane inhibits growth of phenotypically different breast cancer cells. Eur. J. Nutr. 2013, 52, 1949–1958. [Google Scholar] [CrossRef]
- Abbaoui, B.; Lucas, C.R.; Riedl, K.M.; Clinton, S.K.; Mortazavi, A. Cruciferous vegetables, isothiocyanates, and bladder cancer prevention. Mol. Nutr. Food Res. 2018, 62, e1800079. [Google Scholar] [CrossRef]
- Kołodziejski, D.; Koss-Mikołajczyk, I.; Abdi, A.Y.; Jacob, C.; Bartoszek, A. Chemical Aspects of Biological Activity of Isothiocyanates and Indoles, the Products of Glucosinolate Decomposition. Curr. Pharm. Des. 2019, 25, 1717–1728. [Google Scholar] [CrossRef]
- Lim, Y.H.; Chun, J.H.; Lee, K.T.; Hong, S.T.; Lee, Y.H.; Kim, S.J. Changes in Composition and Content of Flavonoids by Processing Type in Rapeseed (Brassica napus) Flowers. Korean Soc. Environ. Agric. 2017, 36, 7–16. [Google Scholar] [CrossRef]
- Wu, L.H.; Luo, Y.M.; Xing, X.R.; Christie, P. EDTA-enhanced phytoremediation of heavy metal contaminated soil with Indian mustard and associated potential leaching risk. Agric. Ecosyst. Environ. 2004, 102, 307–318. [Google Scholar] [CrossRef]
- Feng, H.; Jiao, S.; Tang, Y.; Tang, P.; Liu, J. Advance studying of rape pollen. Food Ind. 2011, 9, 108–111. [Google Scholar]
- Zhu, C.; Du, L.; Gao, Y.; Qian, L.; Gu, L. Suppression effect of Brassica campestris pollen extract on the growth of prostate cancer cell in vitro. Zhejiang J. Integr. Tradit. Chin. West. Med. 2011, 21, 84–86. [Google Scholar]
- Saeidnia, S.; Gohari, A.R. Importance of Brassica napus as a medicinal food plant. J. Med. Plant Res. 2012, 6, 2700–2703. [Google Scholar] [CrossRef]
- Kaur, S.; Shri, R. Comparative study of pharmacognostic parameters, antioxidant and anticholinesterase potential of Brassica napus and Brassica oleracea var. Acephala. IJPSR 2018, 9, 3718–3724. [Google Scholar] [CrossRef]
- Kujawski, R.; Baraniak, J.; Kania, M.; Bartkowiak-Wieczorek, J.; Bogacz, A.; Ożarowski, M.; Cichocki, M.; Spoz, E.; Mikolajczak, P.L. Diet based on oil of seeds of Brassica napus. Implications for the prevention and treatment of prostate diseases. Herba Polonica 2015, 60, 78–88. [Google Scholar] [CrossRef]
- Mussury, R.M.; Fernandes, W.D. Studies of the floral biology and reproductive system of Brassica napus L. (Cruciferae). Braz. Arch. Biol. Technol. 2000, 43, 111–115. [Google Scholar] [CrossRef]
- Bertazzini, M.; Forlani, G. Intraspecific Variability of Floral Nectar Volume and Composition in Rapeseed (Brassica napus L. var. oleifera). Front. Plant Sci. 2016, 7, 288. [Google Scholar] [CrossRef]
- Latifundist Media. Top 10 Rapeseed Producing Countries in 2019. Available online: https://latifundist.com/en/rating/top-10-proizvoditelej-rapsa-v-2019-godu (accessed on 26 August 2021).
- Delifoods. Rapeseed Honey. Available online: https://delimfg.com/natural-honey/rapeseed-honey/ (accessed on 26 August 2021).
- Crops and Horticulture Division Agriculture and Agri-Food Canada. Statistical Overview of the Canadian Honey and Bee Industry. 2019. Available online: https://multimedia.agr.gc.ca/pack/pdf/honey_miel_2019-eng.pdf (accessed on 26 August 2021).
- Ruisinger, B.; Schieberle, P. Characterization of the key aroma compounds in rape honey by means of the molecular sensory science concept. J. Agric. Food Chem. 2012, 60, 4186–4194. [Google Scholar] [CrossRef]
- Kevan, P.G.; Lee, H.; Shuel, R.W. Sugar ratios in nectars of varieties of canola (Brassica napus). J. Apic. Res. 1991, 30, 99–102. [Google Scholar] [CrossRef]
- Stace, P. Protein Content and Amino Acid Profiles of Honey Bee-Collected Pollens; Bees ‘N Trees Consultants: Lismore, Australia, 1996. [Google Scholar] [CrossRef]
- Somerville, D.C. Nutritional Value of Bee Collected Pollens; RIRDC Publication No. 01/047; Rural Industries Research and Development Corporation, New South Wales Agriculture: New South Wales, Australia, 2001. [Google Scholar]
- Rosa, A.S.; Blochtein, B.; Ferreira, N.R.; Witter, S. Apis mellifera (Hymenoptera: Apidae) as a potential Brassica napus pollinator (cv. Hyola 432) (Brassicaceae), in Southern Brazil. Braz. J. Biol. 2010, 70, 1075–1081. [Google Scholar] [CrossRef]
- Rosa, A.S.; Blochtein, B.; Lima, D.K. Honey bee contribution to canola pollination in Southern Brazil. Sci. Agric. 2011, 68, 255–259. [Google Scholar] [CrossRef][Green Version]
- Xie, L.; Xu, B.; Sun, Y.; Zhao, F.; Yin, J.; Geng, Y. The effect of pollination by honeybees on yield of oilseed rape and fatty acid composition of rapeseed. J. Bee 2011, 4, 41–43. [Google Scholar]
- Steffan-Dewenter, I. Seed set of male-sterile and male-fertile oilseed rape (Brassica napus) in relation to pollinator density. Apidologie 2003, 34, 227–235. [Google Scholar] [CrossRef][Green Version]
- Sabbahi, R.; de Oliveira, D.; Marceau, J. Influence of honey bee (Hymenoptera: Apidae) density on the production of canola (Crucifera: Brassicacae). J. Econ. Entomol. 2005, 98, 367–372. [Google Scholar] [CrossRef]
- Durán, X.A.; Ulloa, R.B.; Carrillo, J.A.; Contreras, J.L.; Bastidas, M.T. Evaluation of yield component traits of honeybee-pollinated (Apis mellifera L.) rapeseed canola (Brassica napus L.). Chil. J. Agric. Res. 2010, 70, 309–314. [Google Scholar] [CrossRef]
- Rollin, O.; Garibaldi, L.A. Impacts of honeybee density on crop yield: A meta-analysis. J. Appl. Ecol. 2017, 56, 1152–1163. [Google Scholar] [CrossRef]
- Pernal, S.F.; Currie, R.W. Nectar quality in open-pollinated, pol CMS hybrid, and dominant SI hybrid oilseed summer rape. Can. J. Plant Sci. 1998, 78, 79–89. [Google Scholar] [CrossRef]
- Cutler, C.; Scott-Dupree, C.D. Exposure to Clothianidin Seed-Treated Canola Has No Long-Term Impact on Honey Bees. J. Econ. Entomol. 2007, 100, 765–772. [Google Scholar] [CrossRef]
- Fussel, G.E. History of cale (Brassica sp.). Nature 1995, 176, 48–51. [Google Scholar] [CrossRef]
- Baranyk, P.; Fábry, A. History of the Rapeseed (Brassica napus L.) Growing and Breeding from Middle Age Europe to Canberra. International Rapeseed Congress. 1999. Available online: http://www.regional.org.au/au/gcirc/4/374.htm (accessed on 28 September 2019).
- Cook, S.M.; Skellern, M.P.; Döring, T.F.; Pickett, J.A. Red oilseed rape? The potential for manipulation of petal colour in control strategies for the pollen beetle (Meligethes aeneus). Arthropod-Plant Interact. 2013, 7, 249–258. [Google Scholar] [CrossRef]
- Mushtaq, M.A.; Pan, Q.; Chen, D.; Zhang, Q.; Ge, X.; Li, Z. Comparative Leaves Transcriptome Analysis Emphasizing on Accumulation of Anthocyanins in Brassica: Molecular Regulation and Potential Interaction with Photosynthesis. Front. Plant Sci. 2016, 7, 311. [Google Scholar] [CrossRef]
- Fu, W.; Chen, D.; Pan, Q.; Li, F.; Zhao, Z.; Ge, X.; Li, Z. Production of red-flowered oilseed rape via the ectopic expression of Orychophragmus violaceus OvPAP2. Plant Biotechnol. J. 2018, 16, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Weiss, M.R. Floral color change: A widespread functional convergence. Am. J. Bot. 1995, 82, 167–185. [Google Scholar] [CrossRef]
- Mol, J.; Grotewold, E.; Koes, R. How genes paint flowers and seeds. Trends Plant Sci. 1998, 3, 212–217. [Google Scholar] [CrossRef]
- Deng, J.; Chen, S.; Yin, X.J.; Wang, K.; Liu, Y.L.; Li, S.H.; Yang, P. Systematic qualitative and quantitative assessment of anthocyanins, flavones and flavonols in the petals of 108 lotus (Nelumbo nucifera) cultivars. Food Chem. 2013, 139, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Yin, N.W.; Wang, S.X.; Jia, L.D.; Zhu, M.C.; Yang, J.; Zhou, B.J.; Yin, J.M.; Lu, K.; Wang, R.; Li, J.N.; et al. Identification and Characterization of Major Constituents in Different-Colored Rapeseed Petals by UPLC-HESI-MS/MS. J. Agric. Food Chem. 2019, 67, 11053–11065. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.; Tu, J.; Zhang, Y.; Chen, C.; Yang, Q.; Ten, H.; Niu, J.; Yi, J. Investigation and utilization of cropping forage rape after wheat in northwest area of China. Sci. Technol. West China 2004, 6, 4–7. [Google Scholar]
- Bhuiyan, K.H.; Hossain, M.M.; Bari, M.N.; Khanam, M.R. Identification of bee plants and analysis of honey collected from different plant sources. Pak. J. Biol. Sci. 2002, 5, 1199–1201. [Google Scholar] [CrossRef]
- Khan, T.S.; Mubeen, U. Wheat straw: A pragmatic overview. Curr. Res. J. Biol. Sci. 2012, 4, 673–675. [Google Scholar]
- Huang, R. Effect of Crushed and Ammoniated Rape Straw on Feeding Weining Yellow Cattle. Master’s Thesis, Jiangxi Agricultural University, Nanchang, China, 2013. [Google Scholar]
- Ayres, L.; Clements, B. Forage Brassicas—Quality Crops for Livestock Production; Agfact P2. 1. 13; NSW Agriculture: New South Wales, Australia, 2002; pp. 1–3. [Google Scholar]
- Karaosmanoglu, F.; Isigigur-Ergundenler, A.; Sever, A. Biochar from the straw-stalk of rapeseed plant. Energy Fuels 2000, 14, 336–339. [Google Scholar] [CrossRef]
- Ozcimen, D.; Karaosmanoglu, F. Production and characterization of bio-oil and biochar from rapeseed cake. Renew. Energy 2004, 29, 779–787. [Google Scholar] [CrossRef]
- Azargohar, R.; Nanda, S.; Kang, K.; Bond, T.; Dalai, A.K.; Kozinski, J.A. Effects of bio-additives on the physicochemical properties and mechanical behavior of canola hull fuel pellets. Renew. Energy 2019, 132, 296–307. [Google Scholar] [CrossRef]
- Sánchez-Monedero, M.; Cayuela, L.; Asuncion, R.; Jindo, K.; Mondini, C.; Bolan, N. Role of biochar as an additive in organic waste composting. Bioresour. Technol. 2017, 247, 1155–1164. [Google Scholar] [CrossRef]
- Raza, S.T.; Bo, Z.; Ali, Z.; Liang, T.J. Vermicomposting by Eisenia fetida is a Sustainable and Eco-Friendly Technology for Better Nutrient Recovery and Organic Waste Management in Upland Areas of China. Pak. J. Zool. 2019, 51, 1027–1034. [Google Scholar] [CrossRef]
- Salam, A.; Bashir, S.; Khan, I.; Hu, H.Q. Two years impacts of rapeseed residue and rice straw biochar on Pb and Cu immobilization and revegetation of naturally co-contaminated soil. Appl. Geochem. 2019, 105, 97–104. [Google Scholar] [CrossRef]
- Salam, A.; Bashir, S.; Khan, I.; Rizwan, M.S.; Chhajro, M.A.; Feng, X.W.; Zhu, J.; Hu, H. Biochars immobilize lead and copper in naturally contaminated soil. Environ. Eng. Sci. 2018, 35, 1349–1360. [Google Scholar] [CrossRef]
- Salam, A.; Shaheen, S.M.; Bashir, S.; Khan, I.; Wang, J.X.; Rinklebe, J.; Rehman, F.U.; Hu, H. Rice straw- and rapeseed residue-derived biochars affect the geochemical fractions and phytoavailability of Cu and Pb to maize in a contaminated soil under different moisture content. J. Environ. Manag. 2019, 237, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.W.; Gordon, A.H.; Lomax, J.A.; Chesson, A. Composition and rumen degradability of straw from three varieties of oilseed rape before and after alkali, hydrothermal and oxidative treatment. J. Sci. Food Agric. 1987, 41, 1–15. [Google Scholar] [CrossRef]
- Caraiman, P.; Pohontu, C.; Soreanu, G.; Macoveanu, M.; Cretescu, I. Optimization process of cadmium and zinc removal from soil by phytoremediation using Brassica napus and Triticales sp. Environ. Eng. Manag. J. 2012, 11, 271–278. [Google Scholar] [CrossRef]
- Angelova, V.R.; Ivanova, R.I.; Todorov, J.M.; Ivanov, K. Potential of rapeseed (Brassica napus L.) for phytoremediation of soils contaminated with heavy metals. J. Environ. Prot. ecol. 2017, 18, 468–478. [Google Scholar]
- Belouchrani, A.S.; Mameri, N.; Abdi, N.; Hocine, G.; Lounici, H.; Drouiche, N. Phytoremediation of soil contaminated with Zn using canola (Brassica napus L). Ecol. Eng. 2016, 95, 43–49. [Google Scholar] [CrossRef]
- Adiloglu, S.; Sume, M.T.S.A. Chrome (Cr) pollution in agricultural areas improvement by phytoremediation method with canola (Brassica napus L.) plant growing. J. Essent. Oil Bear. Plants 2015, 18, 1180–1186. [Google Scholar] [CrossRef]
- Gasco, G.; Álvarez, M.L.; Paz-Ferreiro, J.; Mendez, A. Combining phytoextraction by Brassica napus and biochar amendment for the remediation of a mining soil in Riotinto (Spain). Chemosphere 2019, 231, 562–570. [Google Scholar] [CrossRef]
- Dhiman, S.S.; Selvaraj, C.; Li, J.; Singh, R.; Zhao, X.; Kim, D.; Kim, D.; Kim, J.Y.; Kang, Y.C.; Lee, J.K. Phytoremediation of metal-contaminated soils by the hyperaccumulator canola (Brassica napus L.) and the use of its biomass for ethanol production. Fuel 2016, 183, 107–114. [Google Scholar] [CrossRef]
- Gao, J.; Cao, W.; Li, D.; Xu, M.; Zeng, X.; Nie, J.; Zhang, W. Effects of long-term double-rice and green manure rotation on rice yield and soil organic matter in paddy field. Acta Ecol. Sin. 2011, 31, 4542–4548. [Google Scholar]
- Liu, Q.; Dai, Z.; Zhou, X.; Wang, Z.; Cong, R. Effects of rapeseed straw returning on increment of yield and supplying capacity of potassium in rice. Hubei Agric. Sci. 2014, 53, 5112–5115. [Google Scholar]
- Zhou, Y.; Wu, W.; Xu, Y.; Chen, G.; Xu, R.; Zhang, L. Effects of returned rapeseed straw on soil fertility and yield of subsequent rice. J. Yangzhou Univ. 2015, 36, 53–58. [Google Scholar]
- Grove, M.; Spencer, G.; Rohwedder, W.; Mandava, N.; Worley, J.F.; Warthen, J.D., Jr.; Steffens, G.L.; Flippen-Anderson, J.L.; Cook, J.C., Jr. Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen. Nature 1979, 281, 216–217. [Google Scholar] [CrossRef]
- Bernard, E.; Larkin, R.P.; Tavantzis, S.; Erich, M.S.; Alyokhin, A.; Sewell, G.; Lannan, A.; Gross, S.D. Compost, rapeseed rotation, and biocontrol agents significantly impact soil microbial communities in organic and conventional potato production systems. Appl. Soil Ecol. 2012, 52, 29–41. [Google Scholar] [CrossRef]
- Williams-Woodward, J.L.; Pfleger, F.L.; Fritz, A.V.; Allmaras, R.R. Green manures of oat, rape and sweet corn for reducing common root rot in pea (Pisum sativum) caused by Aphanomyces euteiches. Plant Soil 1997, 188, 43–48. [Google Scholar] [CrossRef]
- Ojaghian, M.R.; Cui, Z.Q.; Xie, G.L.; Li, B.; Zhang, J.Z. Brassica green manure rotation crops reduce potato stem rot caused by Sclerotinia sclerotium. Australas. Plant Pathol. 2012, 41, 347–349. [Google Scholar] [CrossRef]
- Cochran, K.A.; Rothrock, C.S. Brassica green manure amendments for management of Rhizoctonia solani in two annual ornamental crops in the field. Hortscience 2015, 50, 555–558. [Google Scholar] [CrossRef]
- Liu, L.; Li, J.; Ren, P.; Zhao, S.; Wang, H.; Li, Y. Effects of different brassica green manure on growth and yield of tobacco. J. Henan Agric. Sci. 2017, 46, 52–76. [Google Scholar]
- Larkin, R.P.; Lynch, R.P. Use and effects of different brassica and other rotation crops on soil borne diseases and yield of potato. Horticulturae 2018, 4, 37. [Google Scholar] [CrossRef]
- Mojtahedi, H.; Santo, G.S.; Wilson, J.H.; Hang, A.N. Managing Meloidogyne chitwoodi on potato with rapeseed as green manure. Plant Dis. 1993, 77, 42–46. [Google Scholar] [CrossRef]
- Mojtahedi, H.; Santo, G.S.; Hang, A.N.; Wilson, J.H. Suppression of root-knot nematode populations with selected rapeseed cultivars as green manure. J. Nematol. 1991, 23, 170–174. [Google Scholar]
- Riga, E.; Mojtahedi, H.; Ingham, R.E.; McGuire, A.M. Green manure amendments and management of root knot nematodes on potato in the Pacific Northwest of USA. Nematol. Monogr. Perspect. 2003, 2, 151–158. [Google Scholar]
- Boydston, R.A.; Hang, A. Rapeseed (Brassica napus) green manure crop suppresses weeds in potato (Solanum tuberosum). Weed Technol. 1995, 9, 669–675. [Google Scholar] [CrossRef]
- Mennan, H.; Ngouajio, M. Effect of brassica cover crops and hazelnut husk mulch on weed control in hazelnut orchards. Hortitechnology 2012, 22, 99–105. [Google Scholar] [CrossRef]
- Lemerle, D.; Luckett, D.J.; Lockley, P.; Koetz, E.; Wu, H.W. Competitive ability of Australian canola (Brassica napus) genotypes for weed management. Crop Pasture Sci. 2014, 65, 1300–1310. [Google Scholar] [CrossRef]
- Madsen, H.; Talgre, L.; Eremeev, V.; Alaru, M.; Kauer, K.; Luik, A. Do green manures as winter cover crops impact the weediness and crop yield in an organic crop rotation? Biol. Agric. Hortic. 2016, 32, 182–191. [Google Scholar] [CrossRef]
- Mesbah, A.; Nilahyane, A.; Ghimire, B.; Beck, L.; Ghimire, R. Efficacy of cover crops on weed suppression, wheat yield, and water conservation in winter wheat-sorghum-fallow. Crop Sci. 2019, 59, 1745–1752. [Google Scholar] [CrossRef]
- Borlaug, N.E.; Dowswell, C.R. Feeding a world of ten billion people: A 21st century challenge. In Proceedings of the International Congress in the Wake of the Double Helix: From the Green Revolution to the Gene Revolution; Iowa State Press: Ames, IA, USA, 2003; pp. 27–31. [Google Scholar] [CrossRef]
- Duvick, D.N. The contribution of breeding to yield advances in maize (Zea mays L.). Adv. Agron. 2005, 86, 83–145. [Google Scholar] [CrossRef]
- Kalloo, G.; Rai, M.; Singh, M.; Kumar, S. Heterosis in Crop Plants; Researchco Book Centre: New Delhi, India, 2006. [Google Scholar]
- Riddle, N.C.; Birchler, J.A. Comparative analysis of inbred and hybrid maize at the diploid and tetraploid levels. Theor. Appl. Genet. 2008, 116, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Blum, A. Heterosis, stress, and the environment: A possible road map towards the general improvement of crop yield. J. Exp. Bot. 2013, 64, 4829–4837. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, G.; Jiang, G.; Zhang, H. Utilization of crop heterosis: A review. Euphytica 2014, 197, 161. [Google Scholar] [CrossRef]
- Munaro, E.M.; Eyherabide, G.H.; D’Andrea, K.E.; Cirilo, A.G.; Otegui, M.E. Heterosis x environment interaction in maize: What drives heterosis for grain yield? Field Crops Res. 2011, 124, 441–449. [Google Scholar] [CrossRef]
- Kaeppler, S.M. Heterosis: Many genes, many mechanisms—End the search for an undiscovered unifying theory. Int. Sch. Res. Not. Bot. 2012, 682824. [Google Scholar] [CrossRef]
- Qian, W.; Sass, O.; Meng, J.; Li, M.; Frauen, M.; Jung, C. Heterotic patterns in rapeseed (Brassica napus L.): I. Crosses between spring and Chinese semi-winter lines. Theor. Appl. Genet. 2007, 115, 27–34. [Google Scholar] [CrossRef]
- Qian, W.; Li, Q.; Noack, J.; Sass, O.; Meng, J.; Frauen, M.; Jung, C. Heterotic patterns in rapeseed (Brassica napus L.): II. Crosses between European winter and Chinese semi-winter lines. Plant Breed. 2009, 128, 466–470. [Google Scholar] [CrossRef]
- Radoev, M.; Becker, H.C.; Ecke, W. Genetic analysis of heterosis for yield and yield components in rapeseed (Brassica napus L.) by quantitative trait locus mapping. Genetics 2008, 179, 1547–1558. [Google Scholar] [CrossRef] [PubMed]
- Becker, H.C.; Engqvist, G.M.; Karlsson, B. Comparison of rapeseed cultivars and resynthesized lines based on allozyme and RFLP markers. Theor. Appl. Genet. 1995, 91, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Girke, A.; Schierholt, A.; Becker, H.C. Extending the rapeseed gene pool with resynthesized Brassica napus II: Heterosis. Theor. Appl. Genet. 2012, 124, 1017. [Google Scholar] [CrossRef]
- Sabaghnia, N.; Dehghani, H.; Alizadeh, B.; Mohghaddam, M. Genetic Analysis of Oil Yield, Seed Yield, and Yield Components in Rapeseed Using Additive Main Effects and Multiplicative Interaction Biplots. Agron. J. 2010, 102, 1361–1368. [Google Scholar] [CrossRef]
- Tian, H.Y.; Channa, S.A.; Hu, S.W. Heterotic Grouping and the Heterotic Pattern among Chinese Rapeseed (Brassica napus L.) Accessions. Agron. J. 2015, 107, 1321–1330. [Google Scholar] [CrossRef]
- Li, Q.; Zhou, Q.; Mei, J.; Zhang, Y.; Li, J.; Li, Z.; Ge, X.; Xiong, Z.; Huang, Y.; Qian, Y. Improvement of Brassica napus via interspecific hybridization between B. napus and B. oleracea. Mol. Breed. 2014, 34, 1955. [Google Scholar] [CrossRef]
- Luo, X.; Ma, C.; Yi, B.; Tu, J.; Shen, J.; Fu, T. Genetic distance revealed by genomic single nucleotide polymorphisms and their relationships with harvest index heterotic traits in rapeseed (Brassica napus L.). Euphytica 2016, 209, 41. [Google Scholar] [CrossRef]
- Shivanna, K.; Sawhney, V. Pollen biology and pollen biotechnology: An introduction. In Pollen Biotechnology for Crop Production and Improvement; Knox, R., Shivanna, K., Sawhney, V., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 1–12. [Google Scholar] [CrossRef]
- Shivanna, K.R. Self-incompatibility. In Pollen Biology and Biotechnology, 1st ed.; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar] [CrossRef]
- Shivanna, K.R. Pollen Sterility. In Pollen Biology and Biotechnology, 1st ed.; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar] [CrossRef]
- Singh, S.; Dey, S.S.; Bhatia, R.; Kumar, R.; Behera, T.K. Current understanding of male sterility systems in vegetable Brassicas and their exploitation in hybrid breeding. Plant Reprod. 2019, 32, 231–256. [Google Scholar] [CrossRef] [PubMed]
- Engelke, T.; Hirsche, J.; Roitsch, T. Metabolically engineered male sterility in rapeseed (Brassica napus L.). Theor. Appl. Genet. 2011, 122, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Ding, Y.; Chen, Z.; Mei, J.; Qian, W. Improvement of the resistance against Sclerotinia sclerotiorum in Ogu CMS restorer in Brassica napus using wild B. oleracea as donor. Sci. Agric. Sin. 2020, 53, 1950–1958. [Google Scholar]
- Li, P.; Kang, L.; Wang, A.; Cui, C.; Jiang, L.; Guo, S.; Ge, X.; Li, Z. Development of a fertility restorer for inap CMS (Isatis indigotica) Brassica napus through genetic introgression of one alien addition. Front. Plant Sci. 2019, 10, 257. [Google Scholar] [CrossRef]
- Heath, D.W.; Earle, E.D. Synthesis of Ogura male sterile rapeseed (Brassica napus L.) with cold tolerance by protoplast fusion and effects of atrazine resistance on seed yield. Plant Cell Rep. 1996, 15, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Kilian, B.; Graner, A. NGS technologies for analyzing germplasm diversity in genebanks. Brief. Funct. Genomics 2012, 11, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Álvarez, N.P.; Khoury, C.K.; Achicanoy, H.A.; Bernau, V.; Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Harker, R.H.; Jarvis, A.; Maxted, N.; et al. Global conservation priorities for crop wild relatives. Nat. Plants 2016, 2, 16022. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Hu, S.; Gardner, C.; Lübberstedt, T. Emerging Avenues for Utilization of Exotic Germplasm. Trends Plant Sci. 2017, 22, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Gorjanc, G.; Jenko, J.; Hearne, S.J.; Hickey, J.M. Initiating maize pre-breeding programs using genomic selection to harness polygenic variation from landrace populations. BMC Genomics 2016, 17, 30. [Google Scholar] [CrossRef]
- Seyis, F.; Snowdon, R.J.; Lühs, W.; Friedt, W. Molecular characterisation of novel resynthesised rapeseed (Brassica napus L.) lines and analysis of their genetic diversity in comparison to spring rapeseed cultivars. Plant Breed. 2003, 122, 473–478. [Google Scholar] [CrossRef]
- Quijada, P.A.; Udall, J.A.; Lambert, B.; Osborn, T.C. Quantitative trait analysis of seed yield and other complex traits in hybrid spring rapeseed (Brassica napus L.): 1. Identification of genomic regions from winter germplasm. Theor. Appl. Genet. 2006, 113, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Udall, J.A.; Quijada, P.A.; Lambert, B.; Osborn, T.C. Quantitative trait analysis of seed yield and other complex traits in hybrid spring rapeseed (Brassica napus L.): 2. Identification of alleles from unadapted germplasm. Theor. Appl. Genet. 2006, 113, 597–609. [Google Scholar] [CrossRef]
- Ahmad, M.A.; Munir, I.; Ali, W.; Swati, Z.A.; Khattak, M.S.; Sohail, Q.; Sohail, Q.; Khan, I. Assessment of Genetic Diversity of Local and Exotic Brassica napus Germplasm. Pak. J. Biol. Sci. 2007, 10, 2490–2494. [Google Scholar] [CrossRef]
- Cowling, W.A.; Buirchell, B.J.; Falk, D.E. A model for incorporating novel alleles from the primary gene pool into elite crop breeding programs while reselecting major genes for domestication or adaptation. Crop Pasture Sci. 2009, 60, 1009–1015. [Google Scholar] [CrossRef]
- Kramer, C.C.; Polewicz, H.; Osborn, T.C. Evaluation of QTL alleles from exotic sources for hybrid seed yield in the original and different genetic backgrounds of spring-type Brassica napus L. Mol. Breed. 2009, 24, 419431. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Long, Y.; Liu, L.; Zhao, Y.; Tian, J.; Zhao, W.; Li, B.; Chen, L.; Chao, H.; et al. Dynamic and comparative QTL analysis for plant height in different developmental stages of Brassica napus L. Theor. Appl. Genet. 2015, 128, 1175–1192. [Google Scholar] [CrossRef]
- Guo, D.; Zhang, J.; Wang, X.; Han, X.; Wei, B.; Wang, J.; Li, B.; Yu, H.; Huang, Q.; Gu, H.; et al. The WRKY Transcription Factor WRKY71/EXB1 Controls Shoot Branching by Transcriptionally Regulating RAX Genes in Arabidopsis. Plant Cell 2015, 27, 3112–3127. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.L.; Ding, G.D.; Xu, F.S.; Cai, H.M.; Zou, J.; Ye, X.S. Genotype differences in photosynthetic characteristics and nitrogen efficiency of new-type oilseed rape responding to low nitrogen stress. J. Agric. Sci. 2015, 153, 1–14. [Google Scholar] [CrossRef]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Würschum, T.; Abel, S.; Zhao, Y. Potential of genomic selection in rapeseed (Brassica napus L.) breeding. Plant Breed. 2014, 133, 45–51. [Google Scholar] [CrossRef]
- Li, J.; Xu, H.H.; Liu, W.C.; Zhang, X.W.; Lu, Y.T. Ethylene Inhibits Root Elongation during Alkaline Stress through AUXIN1 and Associated Changes in Auxin Accumulation. Plant Physiol. 2015, 168, 1777–1791. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Zhao, Y.; Liu, P.; Shi, L.; Wang, X.; Wang, M.; Meng, J.; Reif, J.C. Seed Quality Traits Can Be Predicted with High Accuracy in Brassica napus Using Genomic Data. PLoS ONE 2016, 11, e0166624. [Google Scholar] [CrossRef]
- Yu, J.; Holland, J.B.; McMullen, M.D.; Buckler, E.S. Genetic Design and Statistical Power of Nested Association Mapping in Maize. Genetics 2008, 178, 539–551. [Google Scholar] [CrossRef] [PubMed]
- McMullen, M.D.; Kresovich, S.; Villeda, H.S.; Bradbury, P.; Li, H.; Sun, Q.; Flint-Garcia, S.; Thornsberry, J.; Acharya, C.; Bottoms, C.; et al. Genetic properties of the maize nested association mapping population. Science 2009, 325, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Samans, B.; Snowdon, R.J.; Scholz, U.; Weise, S.; von Wiren, N.; Ordon, F.; Scholz, U.; Weise, S.; von Wirén, N.; Nagel, K.A.; et al. PreBreed-Yield: Population Genomics Resources for Nested Association Mapping and Precision Breeding in Winter Oilseed Rape. In Proceedings of the International Plant and Animal Genome Conference XXI, San Diego, CA, USA, 12–16 January 2013. [Google Scholar] [CrossRef]
- Shunmugam, A.S.K.; Soolanayakanahally, R.; Yang, L.; Horner, K.; Higgins, E.; Lewis, M.; Parkin, I.; Vail, S.; Robinson, S. Characterizing growth physiology, flowering phenology, yield components and seed quality attributes in founder lines of a spring Brassica napus Nested Association Mapping population. In Proceedings of the 14th International Rapeseed Congress, Saskatoon, SK, Canada, 5–9 July 2015. [Google Scholar]
- Li, J.; Bus, A.; Spamer, V.; Stich, B. Comparison of statistical models for nested association mapping in rapeseed (Brassica napus L.) through computer simulations. BMC Plant Biol. 2016, 16, 26. [Google Scholar] [CrossRef]
- Huang, B.E.; Verbyla, K.L.; Verbyla, A.P.; Raghavan, C.; Singh, V.K.; Gaur, P.; Leung, H.; Varshney, R.K.; Cavanagh, C.R. MAGIC populations in crops: Current status and future prospects. Theor. Appl. Genet. 2015, 128, 999. [Google Scholar] [CrossRef]
- Zhao, F.; Zhao, H.; Wang, X.; Li, X. Construction and application potential of MAGIC population on genetic breeding of rapeseed (Brassica napus L.). Chin. J. Oil Crop Sci. 2017, 39, 145–151. [Google Scholar]
- Feuillet, C.; Leach, J.E.; Rogers, J.; Schnable, P.S.; Eversole, K. Crop genome sequencing: Lessons and rationales. Trends Plant Sci. 2011, 16, 77–88. [Google Scholar] [CrossRef]
- Bentley, D.R. Whole-genome re-sequencing. Curr. Opin. Genet. Dev. 2006, 16, 545–552. [Google Scholar] [CrossRef]
- Stratton, M. Genome resequencing and genetic Variation. Nat. Biotechnol. 2008, 26, 65–66. [Google Scholar] [CrossRef]
- Ng, P.C.; Kirkness, E.F. Whole Genome Sequencing. Methods Mol. Biol. 2010, 628, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Bayer, P.E.; Hurgobin, B.; Golicz, A.A.; Chan, C.K.; Yuan, Y.; Lee, H.; Renton, M.; Meng, J.; Li, R.; Long, Y.; et al. Assembly and comparison of two closely related Brassica napus genomes. Plant Biotechnol. J. 2017, 15, 1602–1610. [Google Scholar] [CrossRef]
- Sun, F.; Fan, G.; Hu, Q.; Zhou, Y.; Guan, M.; Tong, C.; Li, J.; Du, D.; Qi, C.; Jiang, L.; et al. The high-quality genome of Brassica napus cultivar ‘ZS11’ reveals the introgression history in semi-winter morphotype. Plant J. 2017, 92, 452–468. [Google Scholar] [CrossRef] [PubMed]
- Song, J.M.; Liu, D.X.; Xie, W.Z.; Yang, Q.Y.; Chen, L.L. BnPIR: Brassica napus pan-genome information resource for 1689 accessions. Plant Biotechnol. J. 2021, 19, 412–414. [Google Scholar] [CrossRef]
- Huang, S.; Deng, L.; Guan, M.; Li, J.; Lu, K.; Wang, H.; Fu, D.; Mason, A.S.; Liu, S.; Hua, W. Identification of genome-wide single nucleotide polymorphisms in allopolyploid crop Brassica napus. BMC Genom. 2013, 14, 717. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Zhou, T.; Ding, G.; Yang, Q.; Shi, L.; Xu, F. Physiological, genomic and transcriptional diversity in responses to boron deficiency in rapeseed genotypes. J. Exp. Bot. 2016, 67, 5769–5784. [Google Scholar] [CrossRef]
- Li, K.; Yao, Y.; Xiao, L.; Zhao, Z.; Guo, S.; Fu, Z.; Du, D. Fine mapping of the Brassica napus Bnsdt1 gene associated with determinate growth habit. Theor. Appl. Genet. 2018, 131, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Li, K.; Liu, H.; Duncan, R.W.; Guo, S.; Xiao, L.; Du, D. Whole-genome re-sequencing and fine mapping of an orange petal color gene (Bnpc1) in spring Brassica napus L. to a 151-kb region. Euphytica 2017, 213, 165. [Google Scholar] [CrossRef]
- Schiessl, S.V.; Huettel, B.; Kuehn, D.; Reinhardt, R.; Snowdon, R.J. Flowering Time Gene Variation in Brassica Species Shows Evolutionary Principles. Front. Plant Sci. 2017, 8, 1742. [Google Scholar] [CrossRef]
- Liu, S.; Fan, C.; Li, J.; Cai, G.; Yang, Q.; Wu, J.; Yi, X.; Zhang, C.; Zhou, Y. A genome wide association study reveals novel elite allelic variations in seed oil content of Brassica napus. Theor. Appl. Genet. 2016, 129, 1203–1215. [Google Scholar] [CrossRef]
- Zhang, K.; Raboanatahiry, N.; Zhu, B.; Li, M. Progress in Genome Editing Technology and Its Application in Plants. Front. Plant Sci. 2017, 8, 177. [Google Scholar] [CrossRef]
- Schiml, S.; Fauser, F.; Puchta, H. The CRISPR/Cas system can be used as nuclease for in planta gene targeting and as paired nickases for directed mutagenesis in Arabidopsis resulting in heritable progeny. Plant J. 2014, 80, 1139–1150. [Google Scholar] [CrossRef]
- Georges, F.; Ray, H. Genome editing of crops: A renewed opportunity for food security. GM Crops Food 2017, 8, 1–12. [Google Scholar] [CrossRef]
- Rodríguez-Leal, D.; Lemmon, Z.H.; Man, J.; Bartlett, M.E.; Lippman, Z.B. Engineering Quantitative Trait Variation for Crop Improvement by Genome Editing. Cell 2017, 171, 470–480. [Google Scholar] [CrossRef]
- Zhang, K.; Nie, L.; Cheng, Q.; Yin, Y.; Chen, K.; Qi, F.; Zou, D.; Liu, H.; Zhao, W.; Wang, B.; et al. Effective editing for lysophosphatidic acid acyltransferase 2/5 in allotetraploid rapeseed (Brassica napus L.) using CRISPR-Cas9 system. Biotechnol. Biofuels 2019, 12, 225. [Google Scholar] [CrossRef]
- Sun, Q.; Lin, L.; Liu, D.; Wu, D.; Fang, Y.; Wu, J.; Wang, Y. CRISPR/Cas9-Mediated Multiplex Genome Editing of the BnWRKY11 and BnWRKY70 Genes in Brassica napus L. Int. J. Mol. Sci. 2018, 19, 2716. [Google Scholar] [CrossRef]
- Sriboon, S.; Li, H.; Guo, C.; Senkhamwong, T.; Dai, C.; Liu, K. Knock-out of TERMINAL FLOWER 1 genes altered flowering time and plant architecture in Brassica napus. BMC Genet. 2020, 21, 52. [Google Scholar] [CrossRef]
- Braatz, J.; Harloff, H.J.; Mascher, M.; Stein, N.; Himmelbach, A.; Jung, C. CRISPR-Cas9 Targeted Mutagenesis Leads to Simultaneous Modification of Different Homoeologous Gene Copies in Polyploid Oilseed Rape (Brassica napus). Plant Physiol. 2017, 174, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wu, J.J.; Tang, T.; Liu, K.D.; Dai, C. CRISPR/Cas9-mediated genome editing efficiently creates specific mutations at multiple loci using one sgRNA in Brassica napus. Sci. Rep. 2017, 7, 7489. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhang, K.; Tang, M.; Zhou, W.; Chen, L.; Chen, Z.; Li, M. CRISPR-based genome editing technology and its applications in oil crops. Oil Crop Sci. 2021, 6, 105–113. [Google Scholar] [CrossRef]





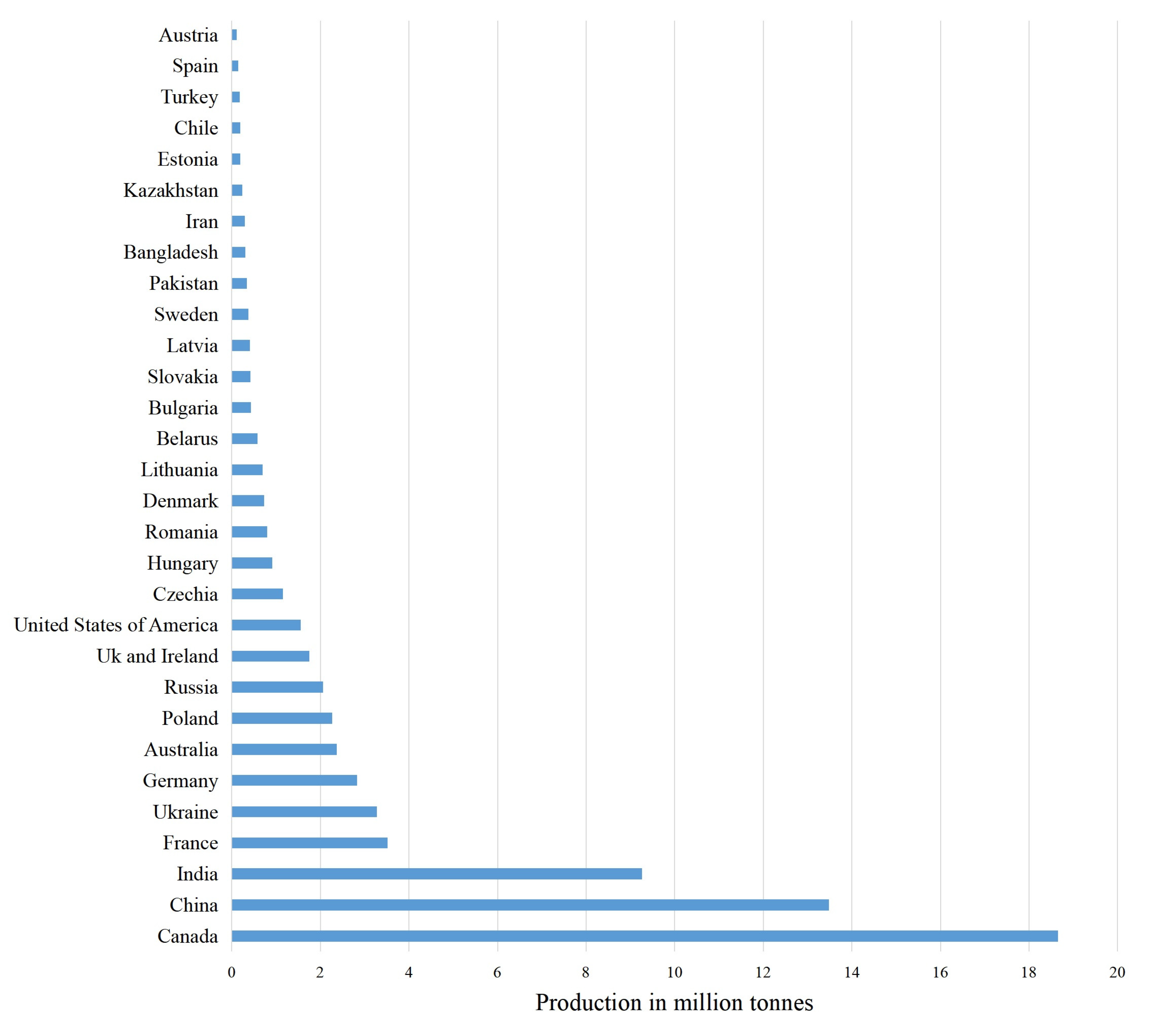{kind=link}
{kind=link}
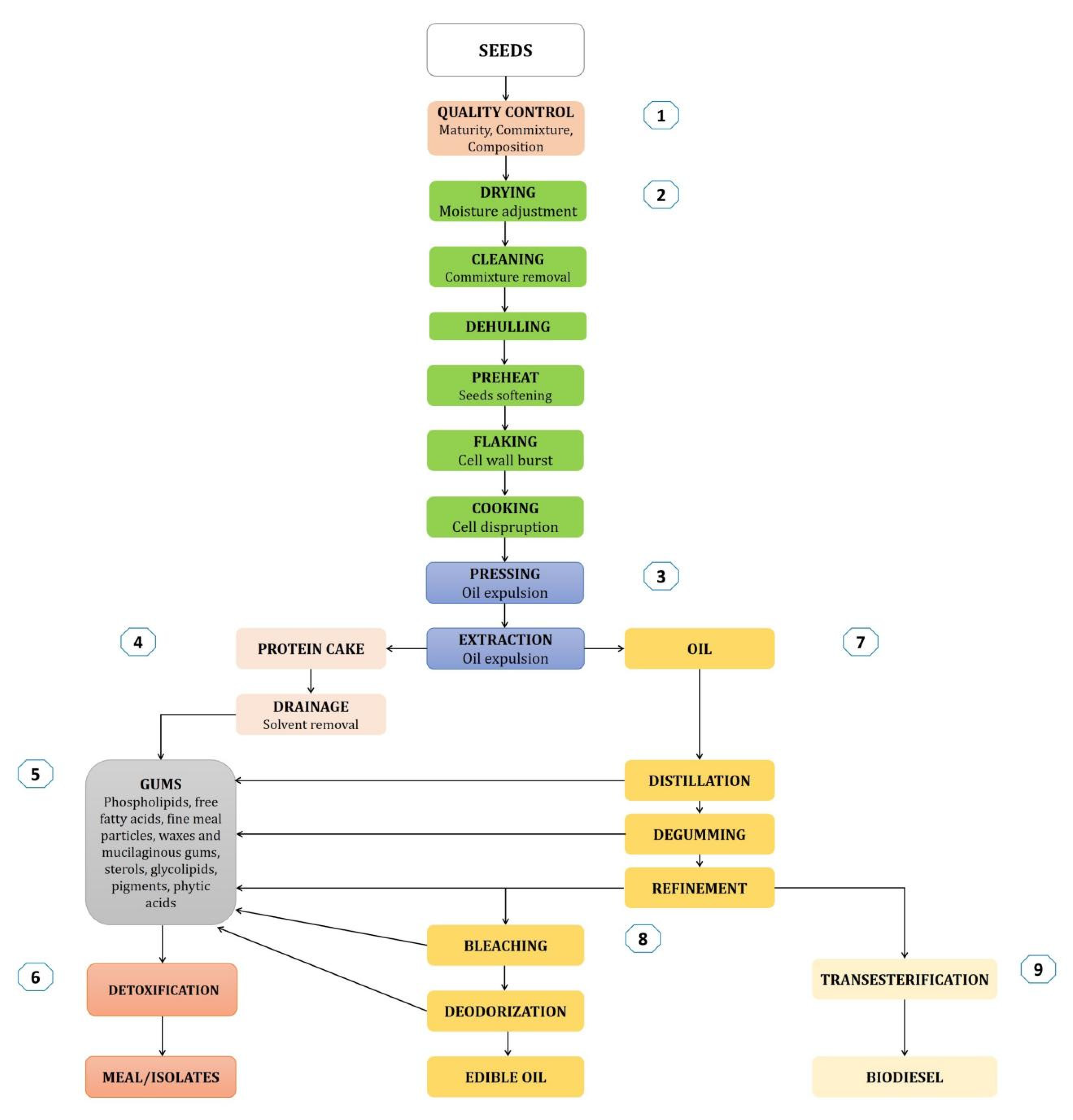{kind=link}
{kind=link}
| Principle | Outcome | Edibility | Advantages | Limitations | References | |
|---|---|---|---|---|---|---|
| Solvent Extraction | Seed compression and usage of hexane to extract the oil | High oil yield | Low toxicity and digestible at hexane, 2% of total food | Can be performed on industrial scale | Possible health issues | [15,16,17,18] |
| Hexane Free Oil Extraction | Enzymatic reaction, usage of gaz, heat or ultrasound | – | Safe for consumption, better taste, and color | Safer for consumption, less time-consuming, better oxidative stability and shelf-life, preserved or improved beneficial compounds, better taste and odor | Not tested on an industrial scale | [18,19] |
| Aqueous Extraction | Water-based extraction, usage of enzyme protease | High oil and protein quality, high oil yield | Safe for consumption | Preserved oil and protein quality, water leftover can be removed by drying. | Not tested on industrial an scale | [20,21,22,23] |
| Heat Treatments | Heat treatments for oil refinement | – | Safe for consumption, but less nutritious | Less glucosinolates compared to the cold pressed treatment. | Denaturation of proteins, formation of transfat, loss of beneficial compounds, not tested on an industrial scale. | [24,25,26,27,28,29,30] |
| Chemicals Free Extraction | Dry fractionation, sieving, ultrafiltration/diafltration, and precipitation | No lignocellulosic fiber, enriched protein and phenolic fractions, less phenolic acids and condensed tannins | Safe for consumption, better taste and color. | Chemical-free, improved isolates taste and color. | Not tested on an industrial scale | [31,32,33,34,35,36,37,38,39] |
| . | Rapeseed | Corn | Olive | Peanut | Soybean | Sunflower | |
|---|---|---|---|---|---|---|---|
| Fatty Acid | SFA | 7% | 13% | 14% | 17% | 16% | 9% |
| MUFA | 62% | 27% | 71% | 45% | 23% | 57% | |
| PUFA | 28% | 54% | 11% | 32% | 57% | 29% | |
| Vitamin * | Alpha–tocopherol | 17.3 mg | 22.6 mg | 20.9 mg | 15.2 mg | 12.2 mg | 68.5 mg |
| Alpha–tocotrienol | – | 1.49 mg | <20.7 mg | <21.5 mg | – | <20.6 mg | |
| Beta–tocopherol | – | 1.1 mg | <10.3 mg | <9.97 mg | 1.8 mg | 2.54 mg | |
| Beta–tocotrienol | 8.07 mg | 4.4 mg | 20.7 mg | <21.5 mg | 1.6 mg | <20.6 mg | |
| Delta–tocopherol | 1.48 mg | 2.78 mg | <10.3 mg | <10.8 mg | 22 mg | <10.3 mg | |
| Delta–tocotrienol | – | – | <20.7 mg | <21.5 mg | – | <20.6 mg | |
| Gamma–tocopherol | 41.3 mg | 60.9 mg | 1.78 mg | 13.4 mg | 70.4 mg | <10.3 mg | |
| Gamma–tocotrienol | – | 1.6 mg | <20.7 mg | <21.5 mg | – | <20.6 mg | |
| Phylloquinone | – | – | 26 µg | 4.2 µg | – | 6.9 µg | |
| Phytosterols * | Stigmasterol | 2.38 mg | 55.3 mg | 1.45 mg | 17.8 mg | 55.4 mg | 29.2 mg |
| Campesterol | 260 mg | 153 mg | 5.34 mg | 33 mg | 58 mg | 33.9 mg | |
| Brassicasterol | 368 mg | 0.414 mg | <0.5 mg | <0.5 mg | – | <0.5 mg | |
| Beta–sitosterol | 368 mg | 538 mg | 128 mg | 122 mg | 153 mg | 205 mg | |
| Delta–5–avenasterol | 24.7 mg | 25.9 mg | 12.1 mg | 16.4 mg | 10.5 mg | 11.4 mg | |
| Campestanol | – | 12.9 mg | <0.5 mg | 0.14 mg | 2.15 mg | <0.5 mg | |
| Beta–sitostanol | – | 30.4 mg | 3.26 mg | 3.58 mg | 5.62 mg | 3.36 mg | |
| Stigmastadiene | – | – | <1 mg | <1 mg | – | <1 mg | |
| Delta–7–Stigmastenol | – | – | 22.4 mg | 8.2 mg | – | 59.1 mg | |
| Other phytosterols | 14.7 mg | 17.8 mg | – | – | 20.8 mg | – |
| Meal Composition | Palatability/Feed Intake | Effect | Animal Performance | References | ||
|---|---|---|---|---|---|---|
| Mammals | Cattle and steer | Rapeseed meals with or without wheat dried grains | Weight gain | Improved milk yield of 1 L/day | [171,172,173,174,176,177] | |
| Pig and sow | 25% of rapeseed meals mixed with wheat, soybean or corn | No change | Better feed efficiency, protein digestibility, and microbial community | Better energy | [38,151,180,181,182] | |
| 40% of cold–pressed rapeseed meals with corn or soybean meal | Reduced | Reduced growth, increased liver and thyroid gland weight, and decreased serum tetraiodothyronine | [183] | |||
| High fiber meals | Reduced digestibility | Reduced energy | [184] | |||
| Dehulled meals | Better P digestibility, no change in body weight | [185] | ||||
| 300 g/kg of rapeseed meals | Better lactic acid bacteria, no impact on digestibility | No change in energy | [186] | |||
| Sheep | Rapeseed meals and other diets | No change on growth and weight | No change on milk yield | [187,188,189] | ||
| Goat | Rapeseed meals and other diets | Weight gain | [190,191] | |||
| Horse | 15% rapeseed meals | No change | [192] | |||
| Rabbit | Rapeseed meals and other diets | Improved growth | [193] | |||
| Poultry | Ostriches | Rapeseed meals and other diets | [194] | |||
| Chicken | Up to 30% of rapeseed diets formulated on digestible amino acid | No change | Improved weight gain | No change in egg production | [195,196,197,198,199] | |
| Rapeseed meals as partial replacement of soybean meals | No change | No bad effects | No change | [200,201] | ||
| 150 g/kg of yellow seed rapeseed meals | Small weight gain | [205,206] | ||||
| Quail | 12% rapeseed meals as partial replacement of soybean meals | Decrease of 17.5% rapeseed meal | No bad effects | No change in meat quality | [202,203] | |
| Turkey | 15% of raw or fermented rapeseed meals with other diets | Improved bodyweight, antioxidant activity, and intestinal histomorphology | Improved meat quality | [148,204] | ||
| 200 g/kg of yellow seed rapeseed meals | No change | No change in amino acid digestibility and bodyweight | [205,206] | |||
| Aquatic Species | Fishes | Rapeseed meals with other diets | No change | [209,210,211,212,213,214] | ||
| Shrimp | 30% rapeseed meals with other diets | No change | No change in growth | No change | [209,210,211,212,213,214,215] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raboanatahiry, N.; Li, H.; Yu, L.; Li, M. Rapeseed (Brassica napus): Processing, Utilization, and Genetic Improvement. Agronomy 2021, 11, 1776. https://doi.org/10.3390/agronomy11091776
Raboanatahiry N, Li H, Yu L, Li M. Rapeseed (Brassica napus): Processing, Utilization, and Genetic Improvement. Agronomy. 2021; 11(9):1776. https://doi.org/10.3390/agronomy11091776
Chicago/Turabian StyleRaboanatahiry, Nadia, Huaixin Li, Longjiang Yu, and Maoteng Li. 2021. "Rapeseed (Brassica napus): Processing, Utilization, and Genetic Improvement" Agronomy 11, no. 9: 1776. https://doi.org/10.3390/agronomy11091776
APA StyleRaboanatahiry, N., Li, H., Yu, L., & Li, M. (2021). Rapeseed (Brassica napus): Processing, Utilization, and Genetic Improvement. Agronomy, 11(9), 1776. https://doi.org/10.3390/agronomy11091776