
Genetic Diversity and Structure of Local Pear Cultivars from Mountainous Areas from Aragon (Northeastern Spain)


Abstract
:1. Introduction
2. Materials and Methods
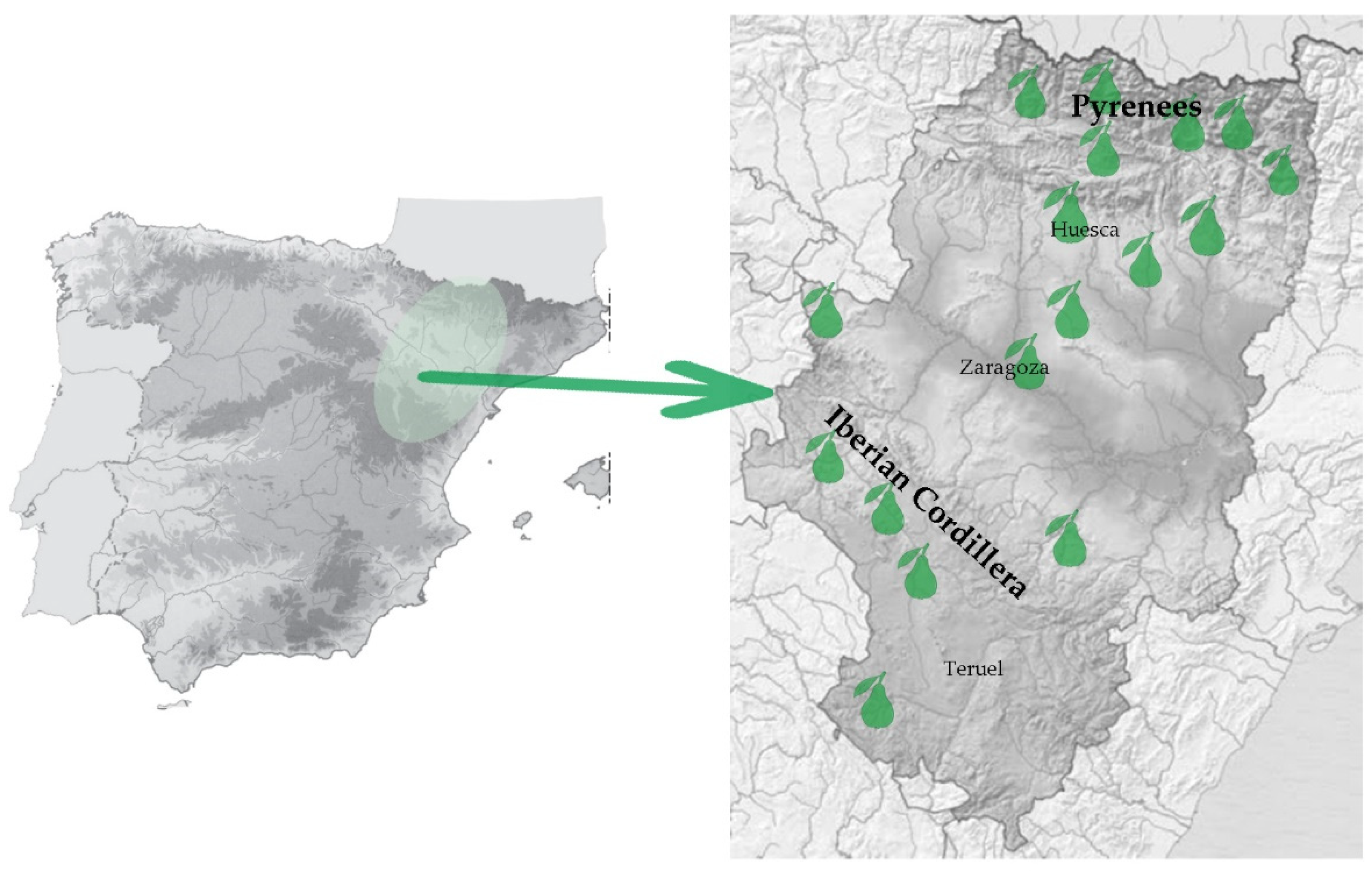
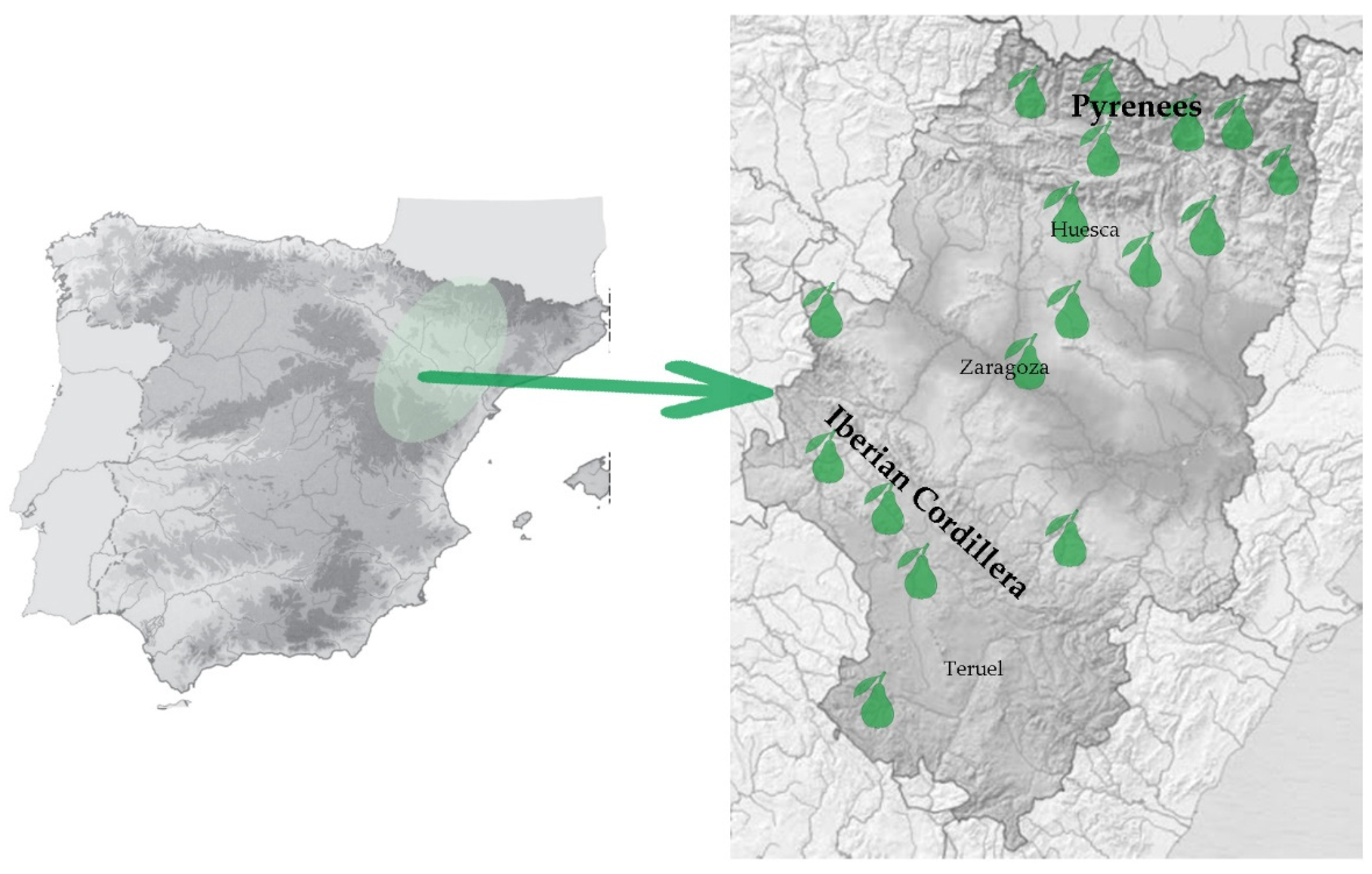
2.1. Plant Material and DNA Extraction
2.2. PCR Reactions and SSR Analysis
2.3. Genetic Diversity Evaluation
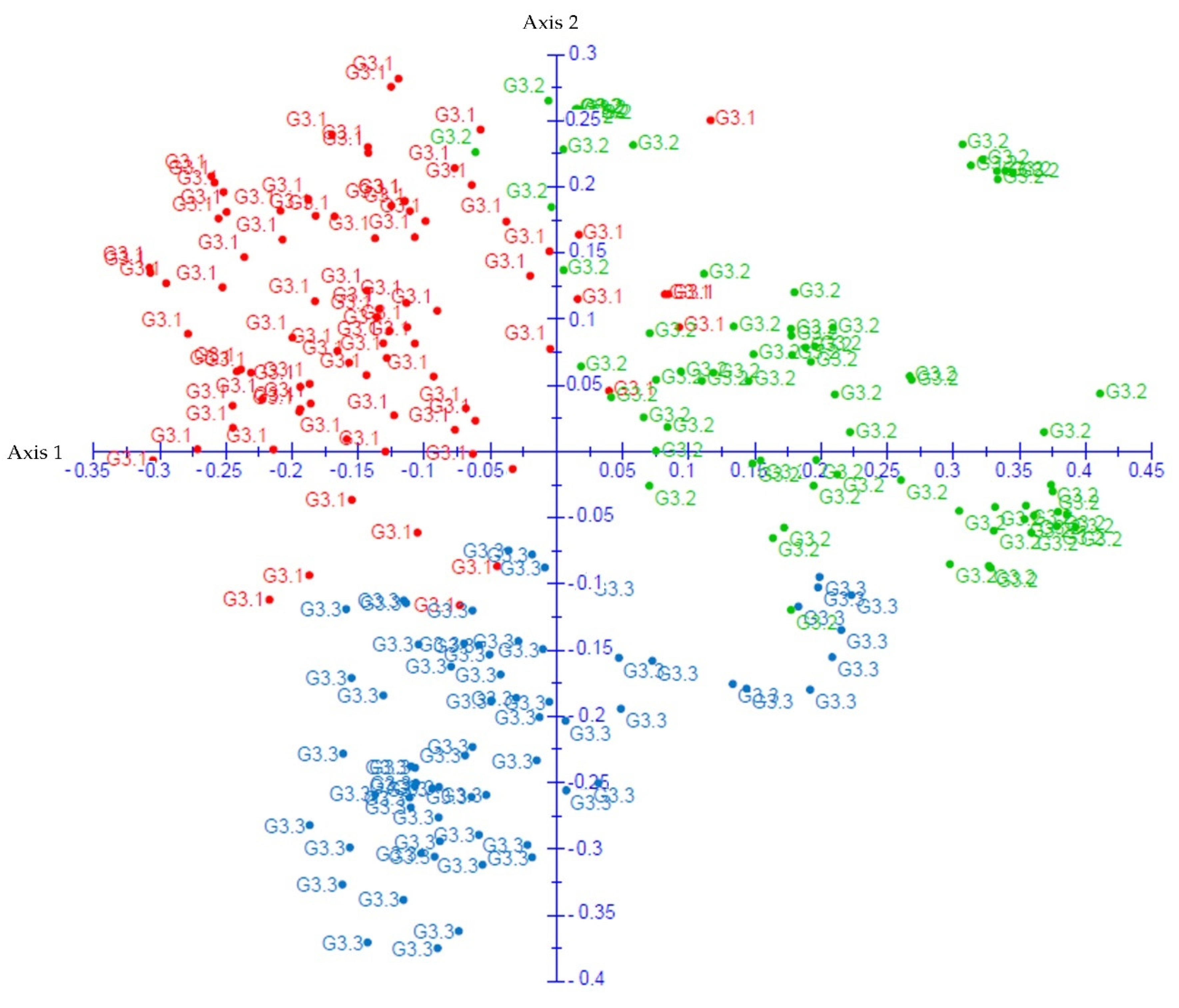
2.4. Analysis of the Genetic Structure, AMOVA and Principal Coordinates (PCoA)
3. Results
3.1. SSR Polymorphism and Genetic Diversity
3.2. Elucidation of Genetic Relationships between Local and Reference Material
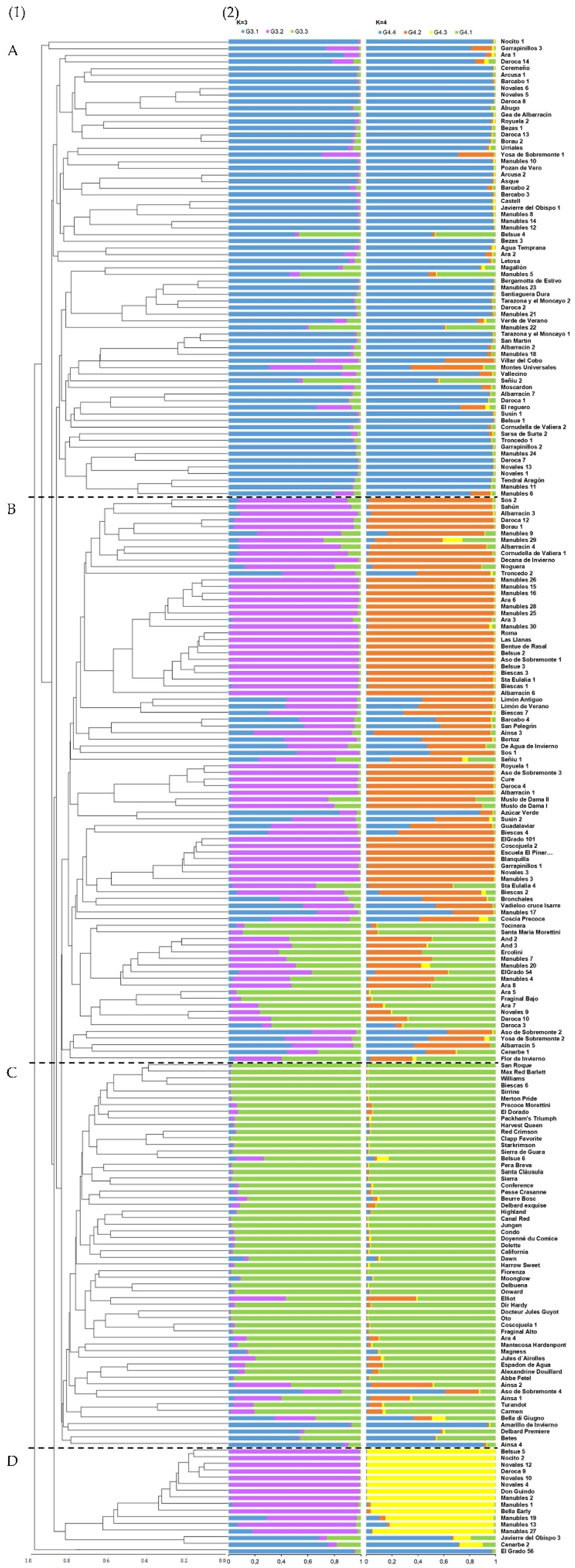
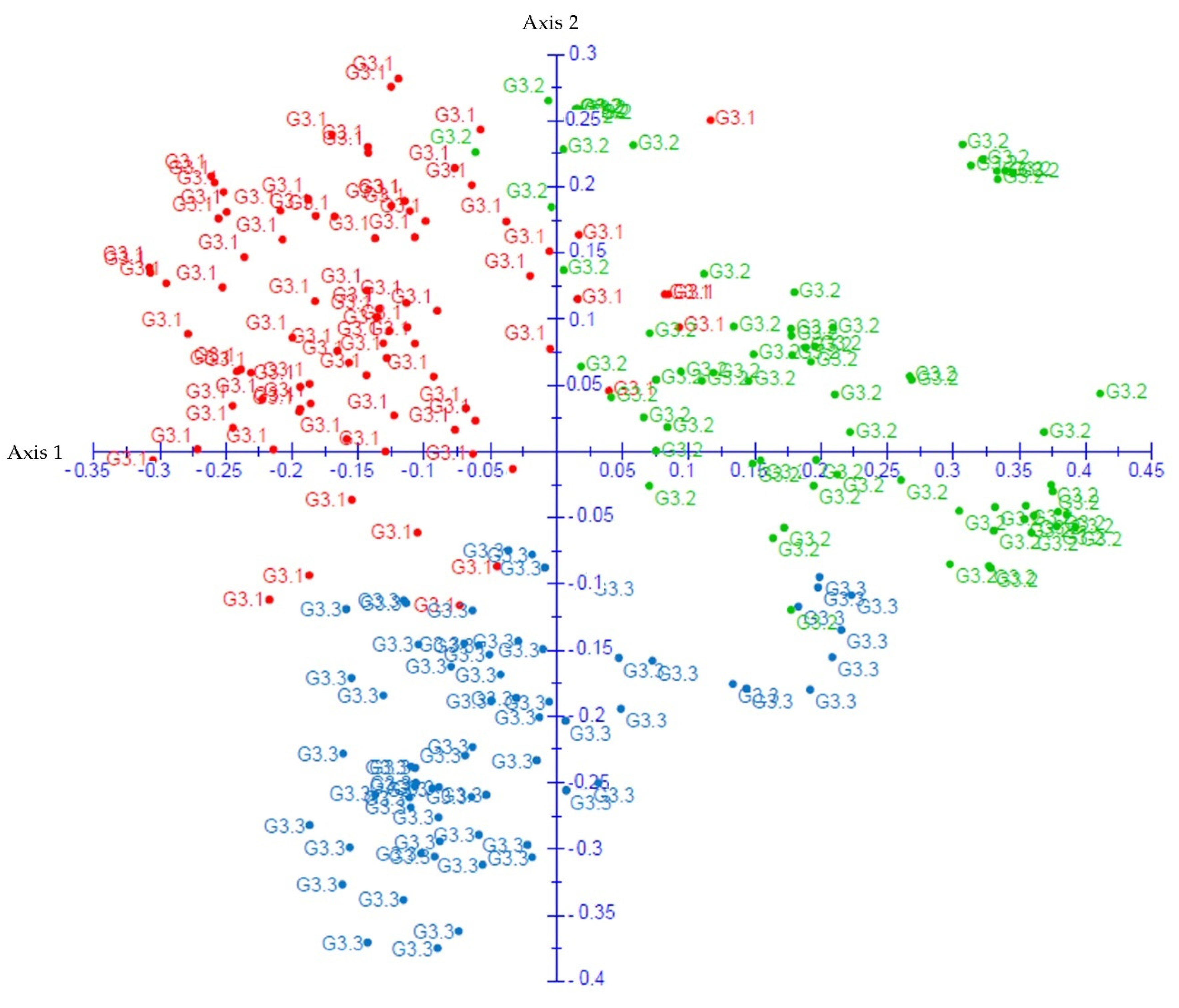
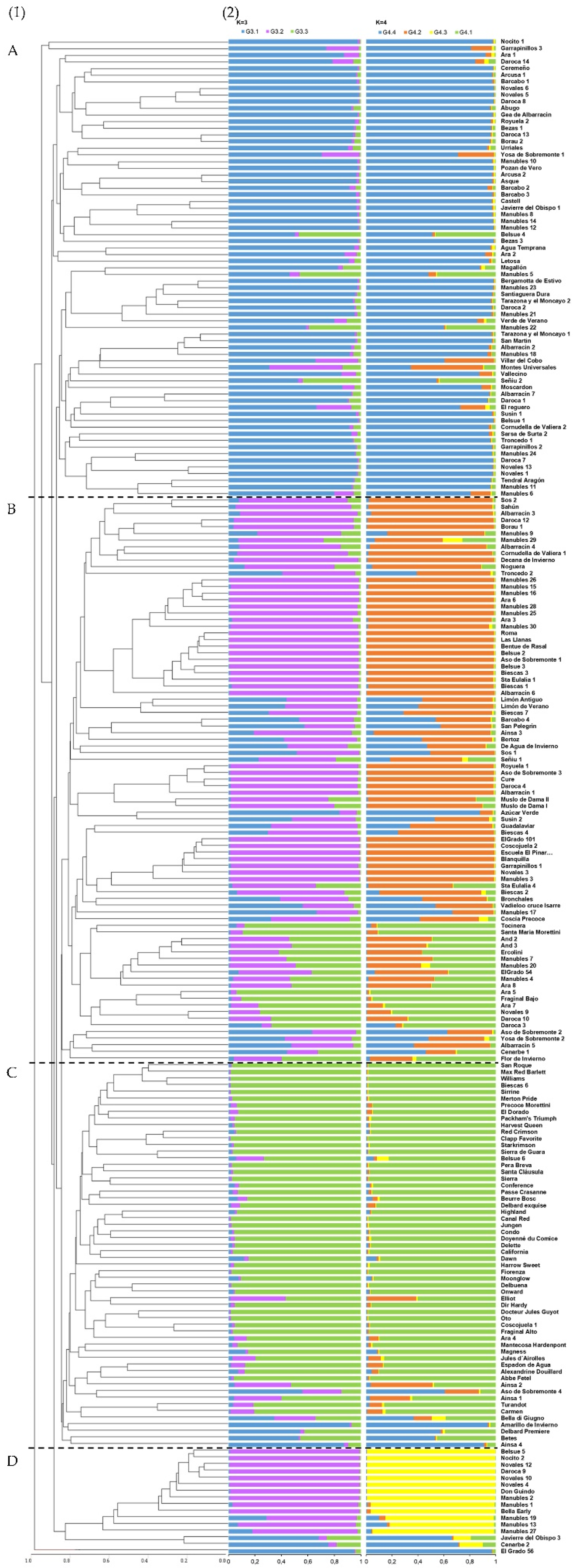
3.3. Population Genetic Structure Analysis
4. Discussion
4.1. Genetic Diversity of Pear Accessions Prospected from Mountain Areas of Aragon
4.2. Singularities of Genetic Relations among Local and Reference Material at CITA Germplasm Bank
4.3. Genetic Structure and Differentiation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Potter, D.; Eriksson, T.; Evans, R.; Oh, S.; Smedmark, J.E.E.; Morgan, D.R.; Kerr, M.; Roberston, K.R.; Arsenault, M.; Dickinson, T.A.; et al. Phylogeny and classification of Rosaceae. Plant Syst. Evol. 2007, 266, 5–43. [Google Scholar] [CrossRef]
- Hummer, K.E.; Janick, J. Rosaceae: Taxonomy, Economic Importance, Genomics. In Genetics and Genomics of Rosaceae, Plant Genetics and Genomics: Crops and Models; Gardiner, S.E., Folta, K.M., Eds.; Springer: New York, NY, USA, 2009; pp. 1–17. [Google Scholar]
- Silva, G.J.; Souza, T.M.; Barbieri, R.L.; Costa de Oliveira, A. Origin, Domestication, and Dispersing of Pear (Pyrus spp.). Adv. Agric. 2014, 1–8. [Google Scholar]
- Yamamoto, T.; Terakami, S. Genomics of pear and other Rosaceae fruit trees. Breed. Sci. 2016, 66, 148–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, R.; Quamme, H.; Layne, R.; Skirvin, R. Pear. In Fruit Breeding (Volume I): Tree and Tropical Fruits; Janick, J., Moore, J.N., Eds.; John Wiley and Sons: New York, NY, USA, 1996; Volume I, pp. 441–514. [Google Scholar]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World:The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin, 4th ed.; Oxford University Press: Oxford, UK, 2012; ISBN 9780199549061. [Google Scholar]
- MAPA. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/agricultura/encuestas-plantaciones-arboles-frutales/ (accessed on 10 May 2021).
- Dondini, L.; Sansavini, S. European pear. In Fruit Breeding; Badenes, L., Byrne, D.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 369–413. [Google Scholar]
- Iglesias, I.; Alegre, S. Peral. In La Fruticultura del Siglo XXI en España; Caja Mar Rural: Almería, Spain, 2014; pp. 57–86. [Google Scholar]
- Lasanta-Martínez, T.; Vicente-Serrano, S.M.; Cuadrat-Prats, J.M. Mountain Mediterranean landscape evolution caused by the abandonment of traditional primary activities: A study of the Spanish Central Pyrenees. Appl. Geogr. 2005, 25, 47–56. [Google Scholar] [CrossRef]
- Garcia-Ruiz, J. La evolución de la agricultura de montaña y sus efectos sobre la dinámica del paisaje. Rev. Estud. Agrosoc. 1988, 146, 7–37. [Google Scholar]
- Arnaez, J.; Lasanta, T.; Errea, M.P.; Ortigosa, L. Land abandonment, landscape evolution, and soil erosion in a spanish mediterranean mountain region: The case of Camero viejo. Land Degrad. Develop. 2001, 22, 537–550. [Google Scholar] [CrossRef]
- Ferreira dos Santos, A.R.; Ramos-Cabrer, A.M.; Díaz-Hernández, M.B.; Pereira-Lorenzo, S. Genetic variability and diversification process in local pear cultivars from northwestern Spain using microsatellites. Tree Genet. Genomes 2011, 7, 1041–1056. [Google Scholar] [CrossRef]
- Miranda, C.; Urrestarazu, J.; Santesteban, L.G.; Royo, J.B.; Urbina, V. Genetic Diversity and Structure in a Collection of Ancient Spanish Pear Cultivars Assessed by Microsatellite Markers. J. Am. Soc. Hortic. Sci. 2010, 135, 428–437. [Google Scholar] [CrossRef] [Green Version]
- Wünsch, A.; Hormaza, J.I. Characterization of variability and genetic similarity of European pear using microsatellite loci developed in apple. Sci. Hortic. 2007, 113, 37–43. [Google Scholar] [CrossRef]
- Harlan, J.R. Our vanishing genetic resources. Science 1975, 188, 618–621. [Google Scholar] [CrossRef]
- Zeven, A.C. Landraces: A review of definitions and classifications. Euphytica 1998, 104, 127–139. [Google Scholar] [CrossRef]
- Alonso Segura, J.M.; Espiau Ramírez, M.T.; Pina Sobrino, A.; Rubio-Cabetas, M.J.; Fernández i Martí, Á. Genetic diversity of the Spanish Pear Germplasm Collection assessed by SSRs. Acta Hortic. 2021, 1303, 37–44. [Google Scholar] [CrossRef]
- Herrero, J. Cartografía de Frutales de Hueso y Pepita. Pomona Hispánica. An. Estac. Exp. Aula Dei 1964. [Google Scholar]
- Herrero, J.; Iturrioz, M. Variedades de Peral en España. An. Estac. Exp. Aula Dei 1971, 11, 200–266. [Google Scholar]
- Schlötterer, C. The evolution of molecular markers-just a matter of fashion? Nat. Rev. Genet. 2004, 5, 63–69. [Google Scholar] [CrossRef]
- Katayama, H.; Uematsu, C. Comparative analysis of chloroplast DNA in Pyrus species: Physical map and gene localization. Theor. Appl. Genet. 2003, 106, 303–310. [Google Scholar] [CrossRef]
- Teng, Y. Genetic relationships of Pyrus species and cultivars native to East Asia revealed by Randomly Amplified Polymorphic DNA markers. J. Amer. Soc. Hort. Sci. 2002, 127, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Bao, L.; Chen, K.; Zhang, D.; Cao, Y.; Yamamoto, T.; Teng, Y. Genetic diversity and similarity of pear (Pyrus L.) cultivars native to East Asia revealed by SSR (simple sequence repeat) markers. Genet. Resour. Crop Evol. 2007, 54, 959. [Google Scholar] [CrossRef]
- Bassil, N.; Postman, J.D. Identification of European and Asian pears using EST-SSRs from Pyrus. Genet. Resour. Crop Evol. 2010, 57, 357–370. [Google Scholar] [CrossRef]
- Wu, J.; Li, L.-T.; Li, M.; Khan, M.A.; Li, X.-G.; Chen, H.; Yin, H.; Zhang, S.-L. High-density genetic linkage map construction and identification of fruit-related QTLs in pear using SNP and SSR markers. J. Exp. Bot. 2014, 65, 5771–5781. [Google Scholar] [CrossRef]
- Evans, K.; Fernández-Fernández, F.; Govan, K. Harmonising fingerprinting protocols to allow camparisons between germplasm collections—Pyrus. Acta Hortic 2009, 814, 103–106. [Google Scholar] [CrossRef]
- Urrestarazu, J.; Royo, J.B.; Santesteban, L.G.; Miranda, C. Evaluating the Influence of the Microsatellite Marker Set on the Genetic Structure Inferred in Pyrus communis L. PLoS ONE 2015, 10, e0138417. [Google Scholar] [CrossRef] [Green Version]
- Ferradini, N.; Lancioni, H.; Torricelli, R.; Russi, L.; Dalla Ragione, I.; Cardinali, I.; Marconi, G.; Gramaccia, M.; Concezzi, L.; Achilli, A.; et al. Characterization and Phylogenetic Analysis of Ancient Italian Landraces of Pear. Front. Plant Sci. 2017, 8, 751. [Google Scholar] [CrossRef] [Green Version]
- Kocsisné, G.M.; Bolla, D.; Anhalt-Brüderl, U.C.M.; Forneck, A.; Taller, J.; Kocsis, L. Genetic diversity and similarity of pear (Pyrus communis L.) cultivars in Central Europe revealed by SSR markers. Genet. Resour. Crop Evol. 2020, 67, 1755–1763. [Google Scholar] [CrossRef]
- Sehic, J.; Garkava-Gustavsson, L.; Fernández-Fernández, F.; Nybom, H. Genetic diversity in a collection of European pear (Pyrus communis) cultivars determined with SSR markers chosen by ECPGR. Sci. Hortic. 2012, 145, 39–45. [Google Scholar] [CrossRef]
- Gasi, F.; Kurtovic, M.; Kalamujic, B.; Pojskic, N.; Grahic, J.; Kaiser, C.; Meland, M. Assessment of European pear (Pyrus communis L.) genetic resources in Bosnia and Herzegovina using microsatellite markers. Sci. Hortic. 2013, 157, 74–83. [Google Scholar] [CrossRef]
- Queiroz, Á.; Bagoin Guimarães, J.; Sánchez, C.; Simões, F.; Maia de Sousa, R.; Viegas, W.; Veloso, M.M. Genetic Diversity and Structure of the Portuguese Pear (Pyrus communis L.) Germplasm. Sustaninability 2019, 11, 5340. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, A.; Assunção, A.; Ramadas, I.; Viegas, W.; Veloso, M.M. Molecular characterization of Portuguese pear landraces (Pyrus communis L.) using SSR markers. Sci. Hortic. 2015, 183, 72–76. [Google Scholar] [CrossRef]
- Pina, A.; Urrestarazu, J.; Errea, P. Analysis of the genetic diversity of local apple cultivars from mountainous areas from Aragon (Northeastern Spain). Sci. Hortic. 2014, 174, 1–9. [Google Scholar] [CrossRef]
- Urrestarazu, J.; Errea, P.; Miranda, C.; Santesteban, L.G.; Pina, A. Genetic diversity of Spanish Prunus domestica L. germplasm reveals a complex genetic structure underlying. PLoS ONE 2018, 13, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennici, S.; Las Casas, G.; Distefano, G.; Di Guardo, M.; Continella, A.; Ferlito, F.; Gentile, A.; Malfa, S. La Elucidating the contribution of wild related species on autochthonous pear germplasm: A case study from Mount Etna. PLoS ONE 2018, 13, e0198512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hormaza, J.I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theor. Appl. Genet. 2002, 104, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, F.; Harvey, N.G.; James, C.M. Isolation and characterization of polymorphic microsatellite markers from European pear (Pyrus communis L.). Mol. Ecol. Notes 2006, 6, 1039–1041. [Google Scholar] [CrossRef]
- Garkava-Gustavsson, L.; Kolodinska Brantestam, A.; Sehic, J.; Nybom, H. Molecular characterisation of indigenous Swedish apple cultivars based on SSR and S-allele analysis. Hereditas 2008, 145, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Liebhard, R.; Gianfranceschi, L.; Koller, B.; Ryder, C.D.; Tarchini, R.; Van De Weg, E.; Gessler, C. Development and characterisation of 140 new microsatellites in apple (Malus x domestica Borkh.). Mol. Breed. 2002, 10, 217–241. [Google Scholar] [CrossRef]
- Hokanson, S.C.; Szewc-McFadden, A.K.; Lamboy, W.F.; McFerson, J.R. Microsatellite (SSR) markers reveal genetic identities, genetic diversity and relationships in a Malus×domestica borkh. core subset collection. Theor. Appl. Genet. 1998, 97, 671–683. [Google Scholar] [CrossRef]
- Gianfranceschi, L.; Seglias, N.; Tarchini, R.; Komjanc, M.; Gessler, C. Simple sequence repeats for the genetic analysis of apple. Theor. Appl. Genet. 1998, 96, 1069–1076. [Google Scholar] [CrossRef]
- Vinatzer, B.A.; Patocchi, A.; Tartarini, S.; Gianfranceschi, L.; Sansavini, S.; Gessler, C. Isolation of two microsatellite markers from BAC clones of the Vf scab resistance region and molecular characterization of scab-resistant accessions in Malus germplasm*. Plant Breed. 2004, 123, 321–326. [Google Scholar] [CrossRef]
- Hardy, O.J.; Vekemans, X. spagedi: A versatile computer program to analyse spatial genetic structure at the individual or population levels. Mol. Ecol. Notes 2002, 2, 618–620. [Google Scholar] [CrossRef] [Green Version]
- Morgante, M.; Vogel, J. Compound microsatellite primers for the detection of genetic polymorphisms. U.S. Patent Application 08/326456, 1994. [Google Scholar]
- Tessier, C.; David, J.; This, P.; Boursiquot, J.M.; Charrier, A. Optimization of the choice of molecular markers for varietal identification in Vitis vinifera L. Theor. Appl. Genet. 1999, 98, 171–177. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Enviroment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org (accessed on 10 May 2021).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Urrestarazu, J.; Miranda, C.; Santesteban, L.G.; Royo, J.B. Genetic diversity and structure of local apple cultivars from Northeastern Spain assessed by microsatellite markers. Tree Genet. Genomes 2012, 8, 1163–1180. [Google Scholar] [CrossRef]
- Jacobs, M.M.; Smulders, M.J.; van den Berg, R.G.; Vosman, B. What’s in a name; Genetic structure in Solanum section Petota studied using population-genetic tools. BMC Evol. Biol. 2011, 11, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, R.; Vershinin, A.; Grzebyta, J.; Shaw, P.; Smýkal, P.; Marshall, D.; Ambrose, M.J.; Ellis, T.N.; Flavell, A.J. The genetic diversity and evolution of field pea (Pisum) studied by high throughput retrotransposon based insertion polymorphism (RBIP) marker analysis. BMC Evol. Biol. 2010, 10, 44. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; von Holdt, B.M. Structure Harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Breton, C.; Pinatel, C.; Médail, F.; Bonhomme, F.; Bervillé, A. Comparison between classical and Bayesian methods to investigate the history of olive cultivars using SSR-polymorphisms. Plant Sci. 2008, 175, 524–532. [Google Scholar] [CrossRef]
- Meirmans, P.G.; Van Tienderen, P.H. genotype and genodive: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. Notes 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin Software. 2006. Available online: http://darwin.cirad.fr/ (accessed on 10 May 2021).
- Pierantoni, L.; Cho, K.-H.; Shin, I.-S.; Chiodini, R.; Tartarini, S.; Dondini, L.; Kang, S.-J.; Sansavini, S. Characterisation and transferability of apple SSRs to two European pear F1 populations. Theor. Appl. Genet. 2004, 109, 1519–1524. [Google Scholar] [CrossRef]
- Brooks, R.; Olmo, H. The Brooks and Olmo Register of Fruit & Nut Varieties, 3rd ed.; American Society for Horticultural Science: Alexandria, VA, USA, 1997. [Google Scholar]
- Baccichet, I.; Foria, S.; Messina, R.; Peccol, E.; Losa, A.; Fabro, M.; Gori, G.; Zandigiacomo, P.; Cipriani, G.; Testolin, R. Genetic and ploidy diversity of pear (Pyrus spp.) germplasm of Friuli Venezia Giulia, Italy. Genet. Resour. Crop Evol. 2020, 67, 83–96. [Google Scholar] [CrossRef]
- Puskás, M.; Höfer, M.; Sestraş, R.E.; Peil, A.; Sestraş, A.F.; Hanke, M.V.; Flachowsky, H. Molecular and flow cytometric evaluation of pear (Pyrus L.) genetic resources of the German and Romanian national fruit collections. Genet. Resour. Crop Evol. 2016, 63, 1023–1033. [Google Scholar] [CrossRef]
- Venturi, S.; Dondini, L.; Donini, P.; Sansavini, S. Retrotransposon characterisation and fingerprinting of apple clones by S-SAP markers. Theor. Appl. Genet. 2006, 112, 440–444. [Google Scholar] [CrossRef]
- Pereira-Lorenzo, S.; Urrestarazu, J.; Ramos-Cabrer, A.M.; Miranda, C.; Pina, A.; Dapena, E.; Moreno, M.A.; Errea, P.; Llamero, N.; Díaz-Hernández, M.B.; et al. Analysis of the genetic diversity and structure of the Spanish apple genetic resources suggests the existence of an Iberian genepool. Ann. Appl. Biol. 2017, 171, 424–440. [Google Scholar] [CrossRef]
- Tatari, M.; Ghasemi, A.; Mousavi, A. Diversity of Local and Wild Pear Germplasm in Central Regions of Iran. Int. J. Fruit Sci. 2020, 20, 432–447. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| MM | Locus | Dye | LG | Allelic Range | μM | Primer Sequence (5′-3′) | Reference | |
|---|---|---|---|---|---|---|---|---|
| A | CH01d08 | PET | 15 | 242–305 | 0.2 | F | CTC CGC CGC TAT AAC ACT TC | [41] |
| R | TAC TCT GGA GGG TAT GTC AAA G | |||||||
| A | CH01d09 | PET | 12 | 120–176 | 0.2 | F | GCC ATC TGA ACA GAA TGT GC | [41] |
| R | CCC TTC ATT CAC ATT TCC AG | |||||||
| A | CH03g07 | VIC | 3 | 204–288 | 0.4 | F | AAT AAG CAT TCA AAG CAA TCC G | [41] |
| R | TTT TTC CAA ATC GAG TTT CGT T | |||||||
| A | CH05c06 | FAM | 16 | 82–118 | 0.2 | F | ATT GGA ACT CTC CGT ATT GTG C | [41] |
| R | ATC AAC AGT AGT GGT AGC CGG T | |||||||
| A | GD142 | FAM | 9 | 126–184 | 0.2 | F | GGCACCCAAGCCCCTAA | [42] |
| R | GGAACCTACGACAGCAAAGTTACA | |||||||
| B | CH02b10 | PET | 2 | 120–162 | 0.6 | F | CAA GGA AAT CAT CAA AGA TTC AAG | [43] |
| R | CAA GTG GCT TCG GAT AGT TG | |||||||
| B | CH02c11 | PET | 10 | 203–249 | 0.2 | F | TGA AGG CAA TCA CTC TGT GC | [41] |
| R | TTC CGA GAA TCC TCT TCG AC | |||||||
| B | CH03d12 | NED | 6 | 91–158 | 0.2 | F | GCC CAG AAG CAA TAA GTA AAC C | [41] |
| R | ATT GCT CCA TGC ATA AAG GG | |||||||
| B | CH-Vf1 | VIC | 1 | 129–172 | 0.2 | F | ATCACCACCAGCAGCAAAG | [44] |
| R | CATACAAATCAAAGCACAACCC | |||||||
| C | GD147 | PET | 13 | 121–167 | 0.2 | F | TCCCGCCATTTCTCTGC | [42] |
| R | AAACCGCTGCTGCTGAAC | |||||||
| C | CH01f07 | NED | 10 | 175–211 | 0.2 | F | CCC TAC ACA GTT TCT CAA CCC | [41] |
| R | CGT TTT TGG AGC GTA GGA AC | |||||||
| C | EMPc11 | VIC | 11 | 135–157 | 0.2 | F | GCGAT TA A AGATCA ATA A ACCCATA | [39] |
| R | AAGCAGCTGGT TGGTGA A AT | |||||||
| C | EMPc117 | FAM | 7 | 88–140 | 0.2 | F | GT TCTATCTACCAAGCCACGCT | [39] |
| R | CGT T TGTGTGTTTTACGTGT TG | |||||||
| C | CH04e03 | FAM | 5 | 175–207 | 0.2 | F | TTG AAG ATG TTT GGC TGT GC | [41] |
| R | TGC ATG TCT GTC TCC TCC AT |
| Overall Set (Ind = 228) | Set of Local Material (Ind = 156) | Set of Reference Material (Ind = 72) | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LOCUS | NA | Ae | Rare Alleles | He | Ho | PD | Fis | NA | Ae | Rare Alleles | He | Ho | PD | Fis | NA | Ae | Rare Alleles | He | Ho | PD | Fis |
| CH03d12 | 15 | 6.14 | 9 | 0.84 | 0.83 | 0.95 | −0.026 | 14 | 6.43 | 8 | 0.84 | 0.84 | 0.95 | −0.022 | 10 | 4.6 | 5 | 0.78 | 0.81 | 0.89 | −0.035 |
| CH-Vf1 | 15 | 7.21 | 8 | 0.86 | 0.87 | 0.96 | −0.048 | 15 | 7.23 | 7 | 0.86 | 0.89 | 0.96 | −0.055 | 9 | 5.84 | 3 | 0.83 | 0.83 | 0.94 | −0.029 |
| Ch01d08 | 17 | 7.02 | 10 | 0.86 | 0.88 | 0.96 | −0.045 | 16 | 6.42 | 8 | 0.84 | 0.87 | 0.95 | −0.043 | 12 | 7.37 | 6 | 0.86 | 0.9 | 0.94 | −0.050 |
| GD142 | 23 | 7.99 | 17 | 0.87 | 0.9 | 0.97 | −0.045 | 23 | 7.85 | 17 | 0.87 | 0.93 | 0.96 | −0.072 | 18 | 7.18 | 12 | 0.86 | 0.85 | 0.96 | 0.015 |
| CH03g07 | 25 | 6.99 | 19 | 0.86 | 0.83 | 0.96 | −0.010 | 22 | 8.01 | 16 | 0.88 | 0.87 | 0.97 | −0.011 | 15 | 4.44 | 9 | 0.77 | 0.76 | 0.92 | −0.005 |
| CH01f07 | 18 | 9.76 | 9 | 0.9 | 0.86 | 0.97 | 0.017 | 18 | 10.63 | 9 | 0.91 | 0.86 | 0.97 | 0.032 | 15 | 7.18 | 8 | 0.86 | 0.87 | 0.94 | −0.019 |
| CH05c06 | 16 | 5.35 | 10 | 0.81 | 0.81 | 0.93 | −0.016 | 15 | 5.53 | 10 | 0.82 | 0.88 | 0.92 | −0.079 | 11 | 4.68 | 6 | 0.79 | 0.67 | 0.91 | 0.147 |
| CH02b10 | 17 | 7.72 | 10 | 0.87 | 0.85 | 0.97 | 0.007 | 17 | 7.6 | 9 | 0.87 | 0.83 | 0.96 | 0.031 | 13 | 6.47 | 7 | 0.85 | 0.88 | 0.94 | −0.045 |
| CH04e03 | 11 | 1.83 | 9 | 0.45 | 0.44 | 0.65 | 0.001 | 10 | 1.67 | 7 | 0.4 | 0.37 | 0.59 | 0.063 | 8 | 2.16 | 5 | 0.54 | 0.58 | 0.72 | −0.097 |
| EMPc117 | 22 | 6.04 | 16 | 0.83 | 0.79 | 0.95 | −0.002 | 22 | 7.44 | 16 | 0.87 | 0.88 | 0.96 | −0.041 | 15 | 3.66 | 12 | 0.73 | 0.61 | 0.89 | 0.133 |
| CH01d09 | 25 | 12.38 | 16 | 0.92 | 0.91 | 0.98 | −0.008 | 23 | 10.81 | 15 | 0.91 | 0.9 | 0.97 | 0.003 | 19 | 10.33 | 12 | 0.9 | 0.93 | 0.97 | −0.033 |
| GD147 | 16 | 3.99 | 12 | 0.75 | 0.74 | 0.91 | −0.014 | 16 | 4.43 | 12 | 0.77 | 0.76 | 0.92 | 0.000 | 13 | 3.06 | 9 | 0.67 | 0.69 | 0.87 | −0.048 |
| CH02c11 | 19 | 7.42 | 12 | 0.87 | 0.88 | 0.96 | −0.030 | 17 | 7.2 | 11 | 0.86 | 0.89 | 0.94 | −0.041 | 13 | 6.94 | 6 | 0.86 | 0.85 | 0.95 | −0.007 |
| EMPc11 | 12 | 5.74 | 6 | 0.83 | 0.86 | 0.95 | −0.082 | 11 | 6.43 | 6 | 0.84 | 0.91 | 0.95 | −0.092 | 11 | 3.69 | 5 | 0.73 | 0.74 | 0.88 | −0.052 |
| Mean | 17.93 | 6.83 | 11.64 | 0.82 | 0.82 | 0.93 | −0.022 | 17.07 | 6.98 | 10.79 | 0.82 | 0.83 | 0.93 | −0.027 | 13 | 5.54 | 7.5 | 0.79 | 0.78 | 0.91 | −0.008 |
| Group | Varieties |
|---|---|
| 1 | Aso de Sobremonte 4–Novales 11 |
| 2 | Albarracin 9–San Martin–Daroca 6 |
| 3 | Susin 2–Biescas 5 |
| 4 | Andorra 2–Andorra 4 |
| 5 | Cornudella de Valiera 1–Cornudella de Valiera 3 |
| 6 | Tarazona y el Moncayo 2–Tarazona y el Moncayo 3 |
| 7 | Troncedo 1–Troncedo 3 |
| 8 | Daroca 3–Daroca 5 |
| 9 | Nocito 2–Albarracin 8 |
| 10 | Caldearenas–Santa Eulalia 2–Novales 12 |
| 11 | Novales 1–Novales 2 |
| 12 | Tendral Aragon (REF)–Tendral Valencia (REF) |
| 13 | Ainsa 3–Sarsa de Surta 1 |
| 14 | Roma (REF)–Santa Eulalia 3–Isarre–Novales 7–Novales 8 |
| 15 | Bezas 1–Bezas 2 |
| 16 | El Grado 101–El Grado 04–Andorra 5 |
| 17 | Tarazona y el Moncayo 4–Coscojuela 2 |
| 18 | Malacara (REF)–Magallon (REF) |
| Genetic Group | n (qI > 0.80 a) | nR (qI > 0.80 a) | % qI > 0.8 | NA | NA/Locus | He | Ho |
|---|---|---|---|---|---|---|---|
| K = 3 | |||||||
| G3.1 | 86 (62) | 13 (10) | 72.09% | 136 | 9.71 | 0.81 | 0.77 |
| G3.2 | 75 (53) | 12 (6) | 70.67% | 126 | 9.00 | 0.78 | 0.90 |
| G3.3 | 67 (51) | 47 (43) | 76.11% | 123 | 8.79 | 0.74 | 0.78 |
| K = 4 | |||||||
| G4.1 | 65 (54) | 48 (44) | 83.07% | 146 | 10.42 | 0.74 | 0.78 |
| G4.2 | 64 (42) | 9 (6) | 65.62% | 138 | 9.85 | 0.75 | 0.89 |
| G4.3 | 13 (13) | 2 (2) | 100% | 56 | 4.00 | 0.62 | 0.95 |
| G4.4 | 86 (65) | 13 (11) | 75.58% | 226 | 16.14 | 0.81 | 0.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bielsa, F.J.; Irisarri, P.; Errea, P.; Pina, A. Genetic Diversity and Structure of Local Pear Cultivars from Mountainous Areas from Aragon (Northeastern Spain). Agronomy 2021, 11, 1778. https://doi.org/10.3390/agronomy11091778
Bielsa FJ, Irisarri P, Errea P, Pina A. Genetic Diversity and Structure of Local Pear Cultivars from Mountainous Areas from Aragon (Northeastern Spain). Agronomy. 2021; 11(9):1778. https://doi.org/10.3390/agronomy11091778
Chicago/Turabian StyleBielsa, Francisco Javier, Patricia Irisarri, Pilar Errea, and Ana Pina. 2021. "Genetic Diversity and Structure of Local Pear Cultivars from Mountainous Areas from Aragon (Northeastern Spain)" Agronomy 11, no. 9: 1778. https://doi.org/10.3390/agronomy11091778
APA StyleBielsa, F. J., Irisarri, P., Errea, P., & Pina, A. (2021). Genetic Diversity and Structure of Local Pear Cultivars from Mountainous Areas from Aragon (Northeastern Spain). Agronomy, 11(9), 1778. https://doi.org/10.3390/agronomy11091778