
Blue Light Upregulates Auxin Signaling and Stimulates Root Formation in Irregular Rooting of Rosemary Cuttings

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Light Irradiation and Growth Conditions
2.3. Determination of Cutting Quality
2.4. qRT-PCR Analysis
2.5. De Novo Assembly and Investigation of IAA Biosynthetic Genes
2.6. Statistical Analysis
3. Results and Discussion
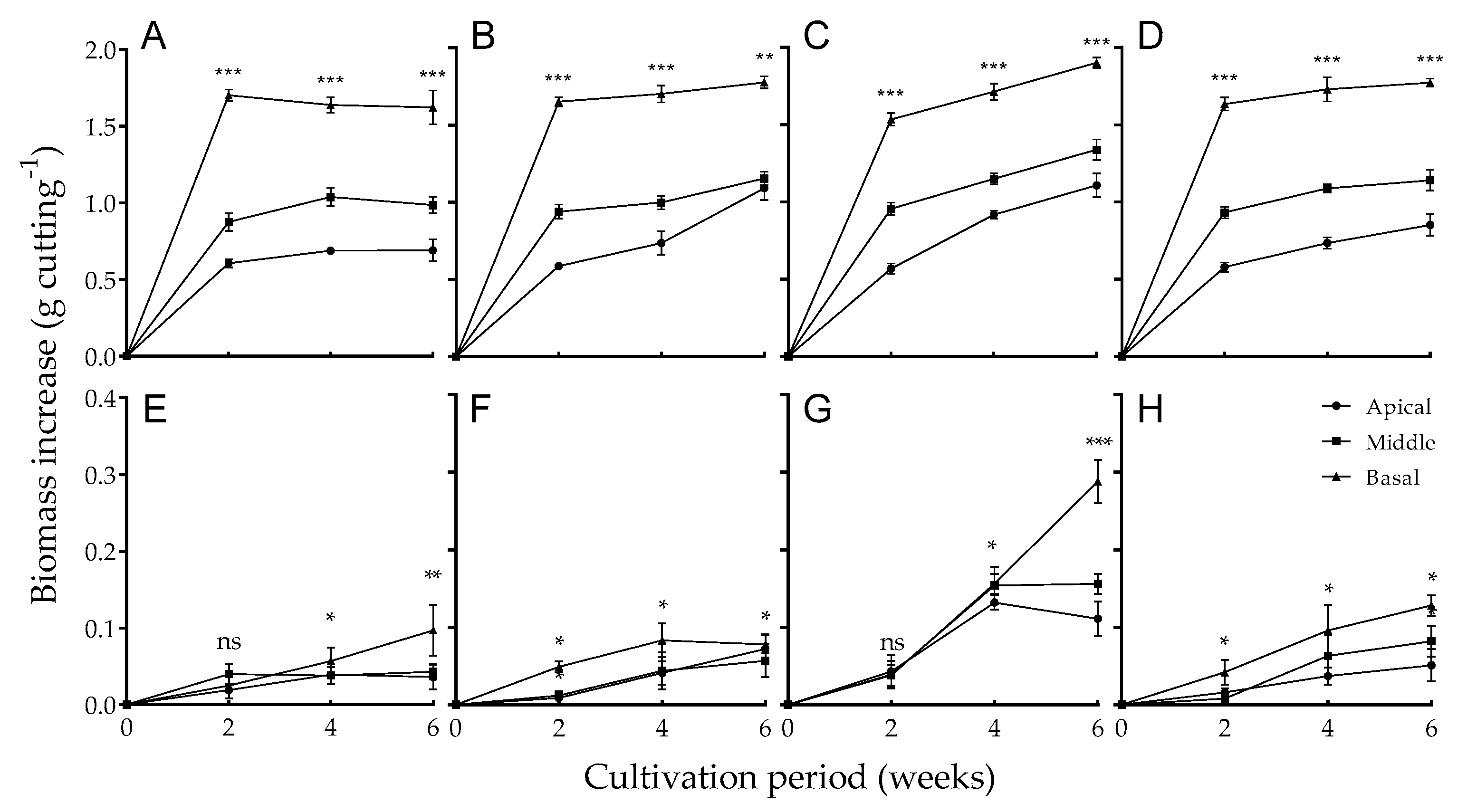
3.1. Shoot and Root Growth Differences among Stem Positions under Different Light Qualities
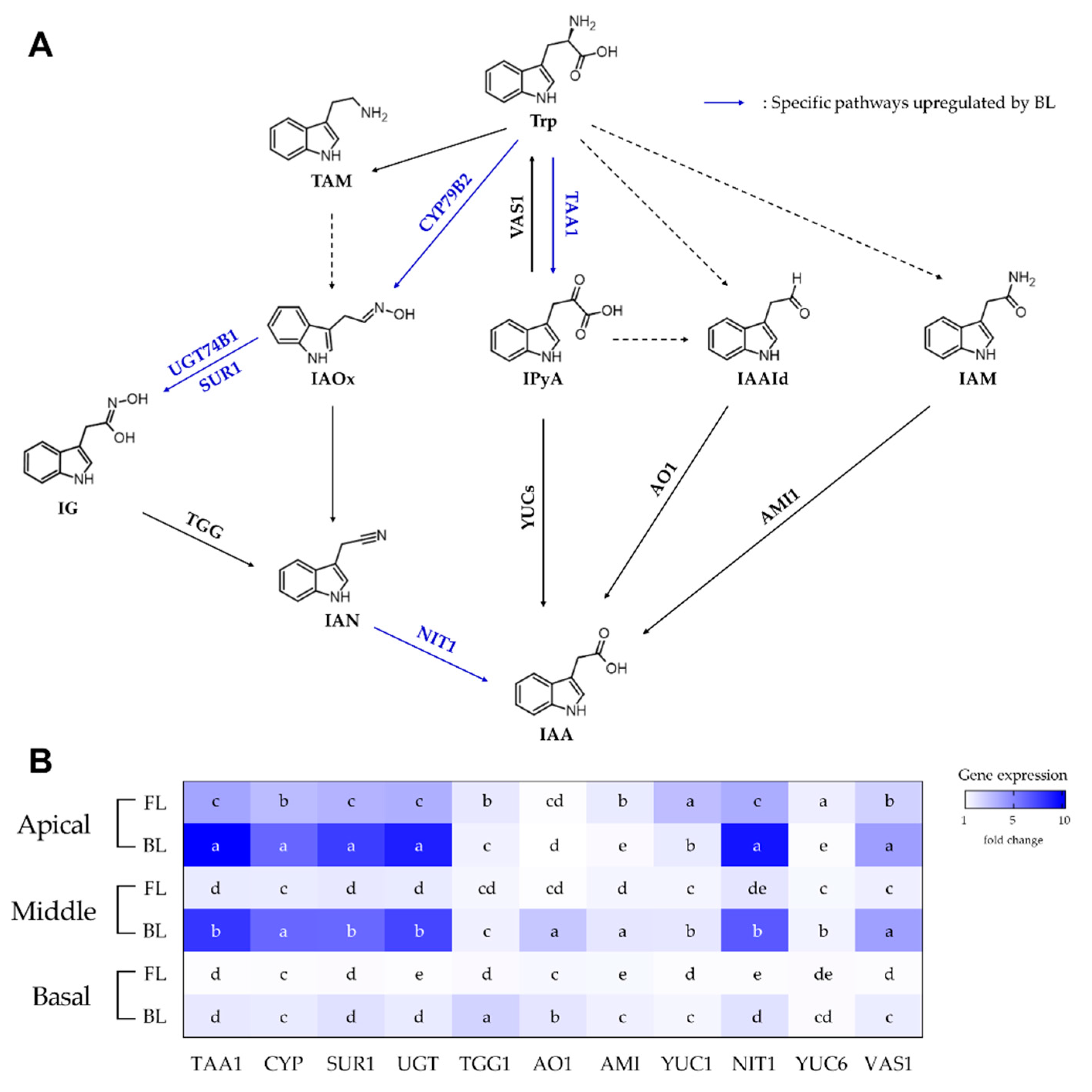
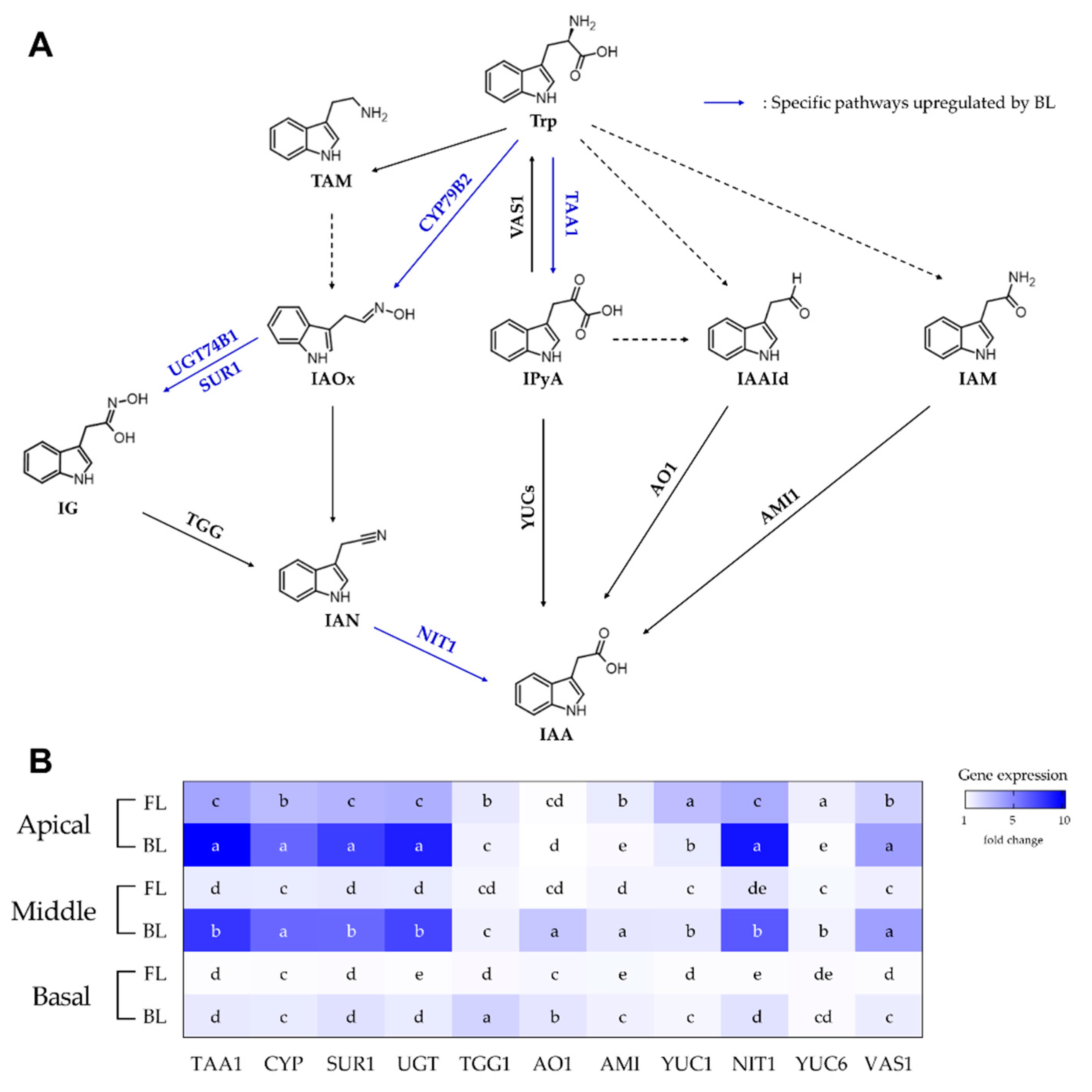
3.2. RNA Expression of IAA Biosynthetic Genes in Rosemary Cuttings
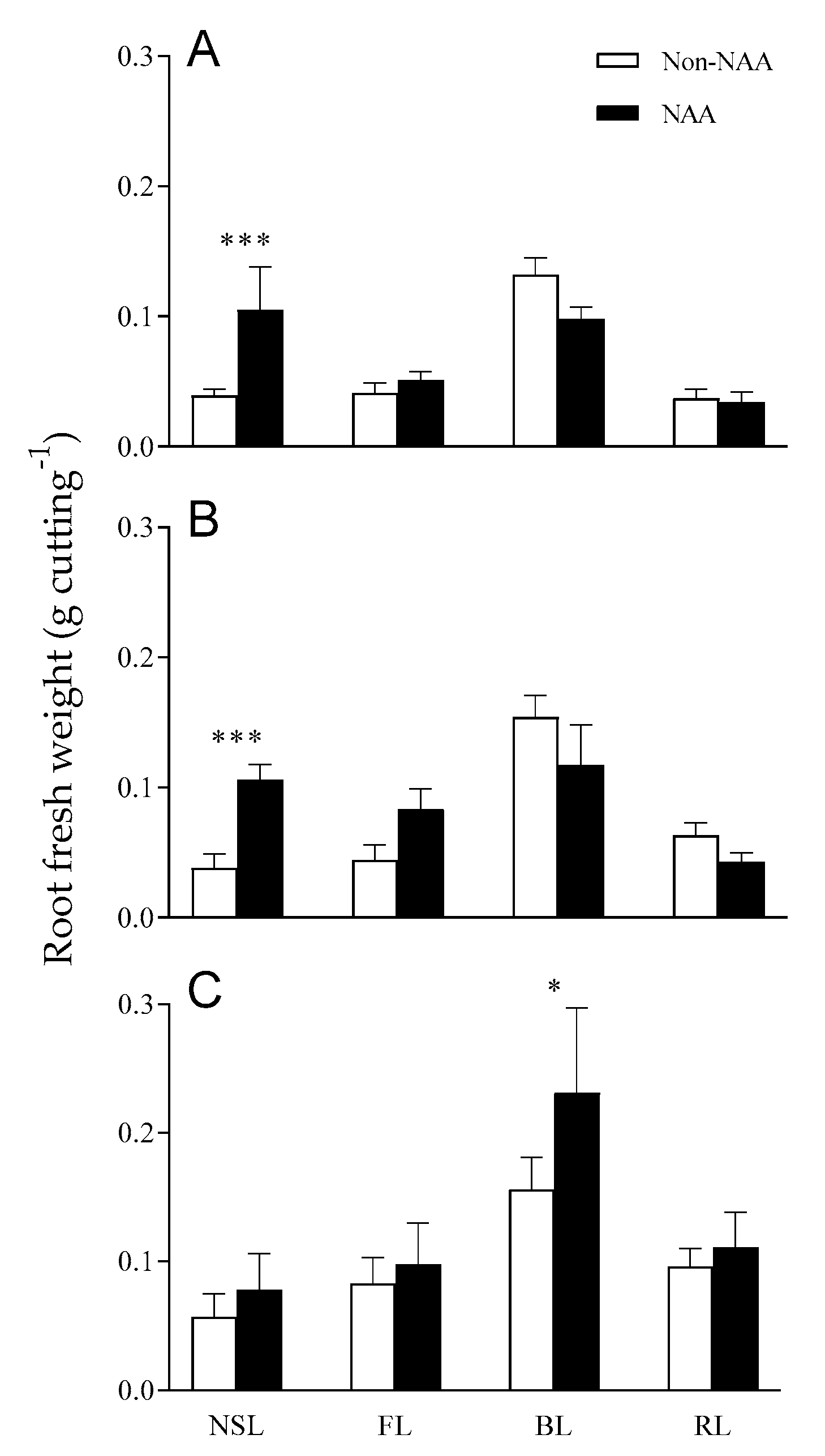
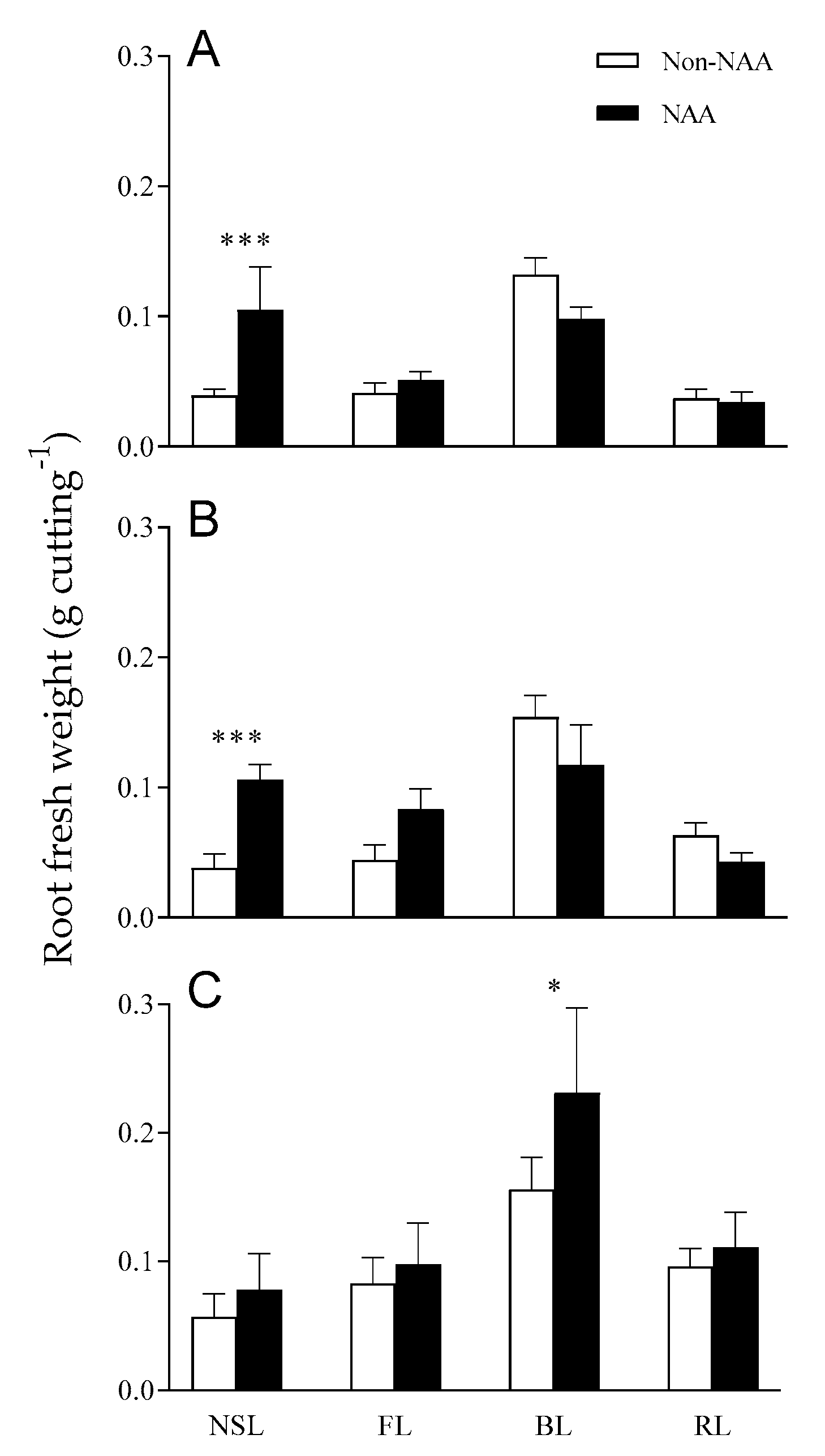
3.3. Effects of NAA Treatment on Root Growth under Different Light Qualities
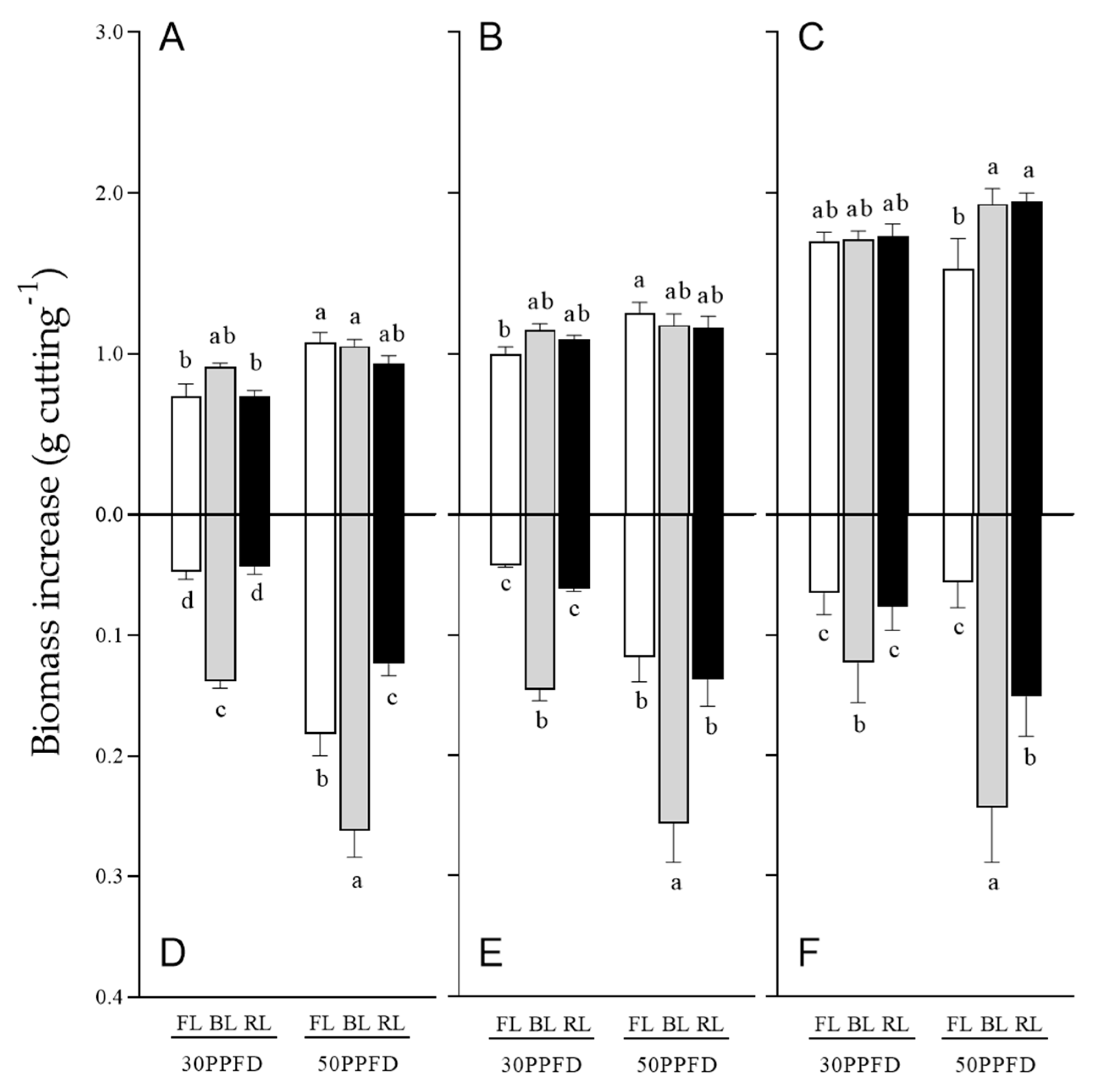
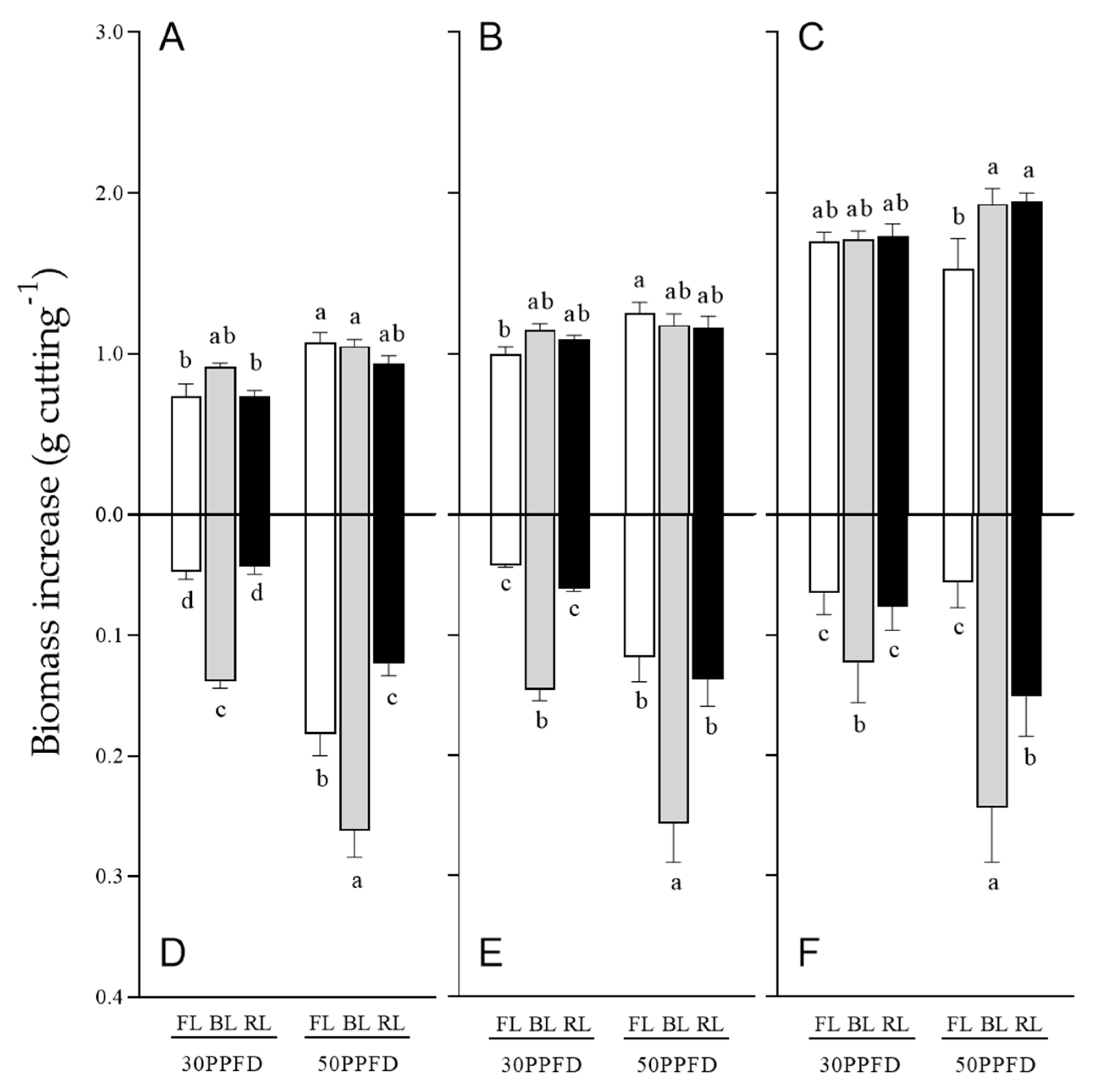
3.4. Effects of Light Intensity on Rosemary Cuttings
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMI1 | AMIDASE 1 |
| AO1 | ALDEHYDE OXIDASE 1 |
| CYP79B2 | CYTOCHROME P450 79B2 |
| IAAId | Indole-3-acetaldehyde |
| IAM | Indole-3-acetamide |
| IAN | Indole-3-acetonitrile |
| IAOx | Indole-3-acetaldoxime |
| IG | Indole glucosinolate |
| IPyA | Indole-3-pyruvic acid |
| NIT1 | NITRILASE 1 |
| SUR1 | SUPERROOT 1 |
| TAA1 | TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS 1 |
| TAM | Tryptamine |
| TGG | β-THIOGLUCOSIDE GLUCOHYDROLASE |
| Trp | L-Tryptophan |
| UGT74B1 | UDP-GLUCOSYL TRANSFERASE |
| VAS1 | REVERSAL OF SAV3 PHENOTYPE 1 |
| YUCs | YUCCAs |
References
- Naiel, M.A.E.; Ismael, N.E.M.; Negm, S.S.; Ayyat, M.S.; Al-Sagheer, A.A. Rosemary leaf powder–supplemented diet enhances performance, antioxidant properties, immune status, and resistance against bacterial diseases in Nile Tilapia (Oreochromis niloticus). Aquaculture 2020, 526, 735370. [Google Scholar] [CrossRef]
- Still, S.M. Manual of Herbaceous Ornamental Plants, 4th ed.; Stipes Publishing Champaign: Champaign, IL, USA, 1994; pp. 570–571. [Google Scholar]
- Pence, V.C. Evaluating costs for the in vitro propagation and preservation of endangered plants. In Vitro Cell. Dev. Biol. 2011, 47, 176–187. [Google Scholar] [CrossRef]
- Hartmann, H.T.; Kester, D.E.; Davies, T.D.; Geneve, T.L., Jr. Plant Propagation: Principles and Practices, 6th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1997; p. 288. [Google Scholar]
- Hansen, J.; Kristensen, K. Axillary bud growth in relation to adventitious root formation in cuttings. Physiol. Plant. 1990, 79, 39–44. [Google Scholar] [CrossRef]
- Bredmose, N.; Kristiansen, K.; Nielsen, B. Propagation temperature, PPFD, auxin treatment, cutting size and cutting position affect root formation, axillary bud growth and shoot development in miniature rose (Rosa hybrida L.) plants and alter homogeneity. J. Hortic. Sci. Biotechnol. 2004, 79, 458–465. [Google Scholar] [CrossRef]
- Gil, C.S.; Jung, H.Y.; Lee, C.; Eom, S.H. Blue light and NAA treatment significantly improve rooting on single leaf-bud cutting of Chrysanthemum via upregulated rooting-related genes. Sci. Hort. 2020, 274, 109650. [Google Scholar] [CrossRef]
- Liu, J.; Sherif, S.M. Hormonal orchestration of bud dormancy cycle in deciduous woody perennials. Front. Plant Sci. 2019, 10, 1136. [Google Scholar] [CrossRef]
- Ersoy, N.; Kalyoncu, I.; Özer, N. Rooting of apical softwood cuttings of Cotoneaster horizontalis Dcne. with application of IBA and air humidity. Selcuk J. Agric. Food Sci. 2017, 30, 67–73. [Google Scholar]
- De Almeida, M.R.; Aumond, M.; Da Costa, C.T.; Schwambach, J.; Ruedell, C.M.; Correa, L.R.; Fett-Neto, A.G. Environmental control of adventitious rooting in Eucalyptus and Populus cuttings. Trees 2017, 31, 1377–1390. [Google Scholar] [CrossRef]
- Lim, Y.J.; Eom, S.H. Effects of different light types on root formation of Ocimum basilicum L. cuttings. Sci. Hort. 2013, 164, 552–555. [Google Scholar] [CrossRef]
- Sager, J.C.; Wheeler, R.M. Application of sunlight and lamps for plant irradiation in space bases. Adv. Space Res. 1992, 12, 133–140. [Google Scholar] [CrossRef]
- Thomas, B.; Dickinson, H.G. Evidence for two photoreceptors controlling growth in de-etiolated seedlings. Planta 1979, 146, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Blaauw, O.H.; Blaauw-Jansen, G. The phototropic responses of Avena coleoptiles. Acta Bot. Neerlandica 1970, 19, 755–763. [Google Scholar] [CrossRef]
- Schwartz, A.; Zeiger, E. Metabolic energy for stomatal opening. Roles of photophosphorylation and oxidative phosphorylation. Planta 1984, 161, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, B.C.; Brown, C.S. Root-shoot interaction in the greening of wheat seedlings grown under red light. Plant Physiol. 1995, 107, 407–411. [Google Scholar] [CrossRef] [Green Version]
- Hoenecke, M.E.; Bula, R.J.; Tibbitts, T.W. Importance of ‘Blue’ photon levels for lettuce seedlings grown under red-light-emitting Diodes. HortScience 1992, 27, 427–430. [Google Scholar] [CrossRef] [Green Version]
- Di, D.W.; Zhang, C.; Luo, P.; An, C.W.; Guo, G.Q. The biosynthesis of auxin: How many paths truly lead to IAA? Plant growth regul. 2016, 78, 275–285. [Google Scholar] [CrossRef]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: Signalling, transport and the control of plant growth and development. Nat. Rev. Mol. cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Ullah, H.; Chen, J.G.; Temple, B.; Boyes, D.C.; Alonso, J.M.; Davis, K.R.; Ecker, J.R.; Jones, A.M. The β-subunit of the Arabidopsis G protein negatively regulates auxin-induced cell division and affects multiple developmental processes. Plant Cell 2003, 15, 393–409. [Google Scholar] [CrossRef] [Green Version]
- Tivendale, N.D.; Ross, J.J.; Cohen, J.D. The shifting paradigms of auxin biosynthesis. Trends Plant Sci. 2014, 19, 44–51. [Google Scholar] [CrossRef]
- Shirke, P.A. Leaf photosynthesis, dark respiration and fluorescence as influenced by leaf age in an evergreen tree, Prosopis juliflora. Photosynthetica 2001, 39, 305–311. [Google Scholar] [CrossRef]
- Christie, J.M. Phototropin blue-light receptors. Annu. Rev. Plant Biol. 2007, 58, 21–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goins, G.D.; Yorio, N.C.; Sanwo-Lewandowski, M.M.; Brown, C.S. Life cycle experiments with Arabidopsis grown under red light-emitting diodes (LEDs). Life Support Biosphere Sci. 1998, 5, 143–149. [Google Scholar]
- Johnson, D.M.; Smith, W.K.; Vogelmann, T.C.; Brodersen, C.R. Leaf architecture and direction of incident light influence mesophyll fluorescence profiles. Am. J. Bot. 2005, 92, 1425–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, W.K.; Vogelmann, T.C.; DeLucia, E.H.; Bell, D.T.; Shepherd, K.A. Leaf form and photosynthesis. Bioscience 1997, 47, 785–793. [Google Scholar] [CrossRef]
- Amri, E.; Lyaruu, H.V.M.; Nyomora, A.S.; Kanyeka, Z.L. Vegetative propagation of African Blackwood (Dalbergia melanoxylon Guill. & Perr.): Effects of age of donor plant, IBA treatment and cutting position on rooting ability of stem cuttings. New For. 2010, 39, 183–194. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Hall, R.B.; Bauer, E.O.; Riemenschneider, D.E. Shoot position affects root initiation and growth of dormant unrooted cuttings of Populus. Silvae Genet. 2003, 52, 273–279. [Google Scholar]
- Moon, H.K.; Park, S.Y.; Kim, Y.W.; Kim, C.S. Growth of Tsuru-rindo (Tripterospermum japonicum) cultured in vitro under various sources of light-emitting diode (LED) irradiation. J. Plant Biol. 2006, 49, 174–179. [Google Scholar] [CrossRef]
- Alallaq, S.; Ranjan, A.; Brunoni, F.; Novák, O.; Lakehal, A.; Bellini, C. Red light controls adventitious root regeneration by modulating hormone homeostasis in Picea abies seedlings. Front. Plant Sci. 2020, 11, 1397. [Google Scholar] [CrossRef]
- Sæbø, A.; Skjeseth, G.; Appelgren, M. Light quality of the in vitro stage affects the subsequent rooting and field performance of Betula pendula (Roth). Scand. J. For. Res. 1995, 10, 155–160. [Google Scholar] [CrossRef]
- Quittenden, L.J.; Davies, N.W.; Smith, J.A.; Molesworth, P.P.; Tivendale, N.D.; Ross, J.J. Auxin biosynthesis in pea: Characterization of the tryptamine pathway. Plant Physiol. 2009, 151, 1130–1138. [Google Scholar] [CrossRef] [Green Version]
- Normanly, J.; Slovin, J.P.; Cohen, J.D. Auxin biosynthesis and metabolism. In Plant Horm; Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 36–62. [Google Scholar]
- Li, R.; Jiang, J.; Jia, S.; Zhu, X.; Su, H.; Li, J. Overexpressing broccoli tryptophan biosynthetic genes BoTSB1 and BoTSB2 promotes biosynthesis of IAA and indole glucosinolates. Physiol. Plant. 2020, 168, 174–187. [Google Scholar] [CrossRef]
- Casanova-Sáez, R.; Voß, U. Auxin metabolism controls developmental decisions in land plants. Trends Plant Sci. 2019, 24, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Normanly, J.; Grisafi, P.; Fink, G.R.; Bartel, B. Arabidopsis mutants resistant to the auxin effects of indole-3-acetonitrile are defective in the nitrilase encoded by the NIT1 gene. Plant Cell 1997, 9, 1781–1790. [Google Scholar] [PubMed]
- Seo, M.; Akaba, S.; Oritani, T.; Delarue, M.; Bellini, C.; Caboche, M.; Koshiba, T. Higher activity of an aldehyde oxidase in the auxin-overproducing superroot1 mutant of Arabidopsis thaliana. Plant Physiol. 1998, 116, 687–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H.; et al. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Li, T.; Zhou, M.; Li, M.; Zhao, Y.; Xu, J.; Hu, F.; Li, H. Caenorhabditis elegans extracts stimulate IAA biosynthesis in Arthrobacter pascens ZZ21 via the indole-3-pyruvic acid pathway. Microorganisms 2021, 9, 970. [Google Scholar] [CrossRef]
- Singh, K.K.; Tomar, Y.K. Effect of planting time and indole butyric acid levels on rooting of woody cuttings of phalsa (Grewia asiatica L.). HortFlora Res. Spect. 2015, 4, 39–43. [Google Scholar]
- Singh, K.K.; Chauhan, J.S.; Rawat, J.M.S.; Rana, D.K. Effect of different growing conditions and various concentrations of IBA on the rooting and shooting of hardwood cutting of phalsa (Grewia asetica L.) under valley condition of Garhwal Himalayas. Plant Arch. 2015, 15, 131–136. [Google Scholar]
- Braha, S.; Rama, P. The effects of indol butyric acid and naphthalene acetic acid of adventitious root formation to green cuttings in blueberry cv. Vaccinium corymbosum L. Int. J. Sci. Res. 2016, 5, 876–879. [Google Scholar]
- Gonin, M.; Bergougnoux, V.; Nguyen, T.D.; Gantet, P.; Champion, A. What makes adventitious roots? Plants 2019, 8, 240. [Google Scholar] [CrossRef] [Green Version]
- Eliasson, L.; Brunes, L. Light effects on root formation in aspen and willow cuttings. Physiol. Plantarum 1980, 48, 261–265. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gil, C.-S.; Kwon, S.-J.; Jeong, H.-Y.; Lee, C.; Lee, O.-J.; Eom, S.-H. Blue Light Upregulates Auxin Signaling and Stimulates Root Formation in Irregular Rooting of Rosemary Cuttings. Agronomy 2021, 11, 1725. https://doi.org/10.3390/agronomy11091725
Gil C-S, Kwon S-J, Jeong H-Y, Lee C, Lee O-J, Eom S-H. Blue Light Upregulates Auxin Signaling and Stimulates Root Formation in Irregular Rooting of Rosemary Cuttings. Agronomy. 2021; 11(9):1725. https://doi.org/10.3390/agronomy11091725
Chicago/Turabian StyleGil, Chan-Saem, Soon-Jae Kwon, Ho-Young Jeong, Chanhui Lee, Oak-Jin Lee, and Seok-Hyun Eom. 2021. "Blue Light Upregulates Auxin Signaling and Stimulates Root Formation in Irregular Rooting of Rosemary Cuttings" Agronomy 11, no. 9: 1725. https://doi.org/10.3390/agronomy11091725
APA StyleGil, C.-S., Kwon, S.-J., Jeong, H.-Y., Lee, C., Lee, O.-J., & Eom, S.-H. (2021). Blue Light Upregulates Auxin Signaling and Stimulates Root Formation in Irregular Rooting of Rosemary Cuttings. Agronomy, 11(9), 1725. https://doi.org/10.3390/agronomy11091725