
Floral and Pollen Traits of Moringa oleifera Lam. and Moringa peregrina (Forssk.) Fiori Provide Reproductive Adaptations for Arid Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Pollen Viability and Germinability Tests
2.2. Removal of the Pollenkitt
2.3. Test of Pollenkitt for Pollen Insertion into the Stylar Tube
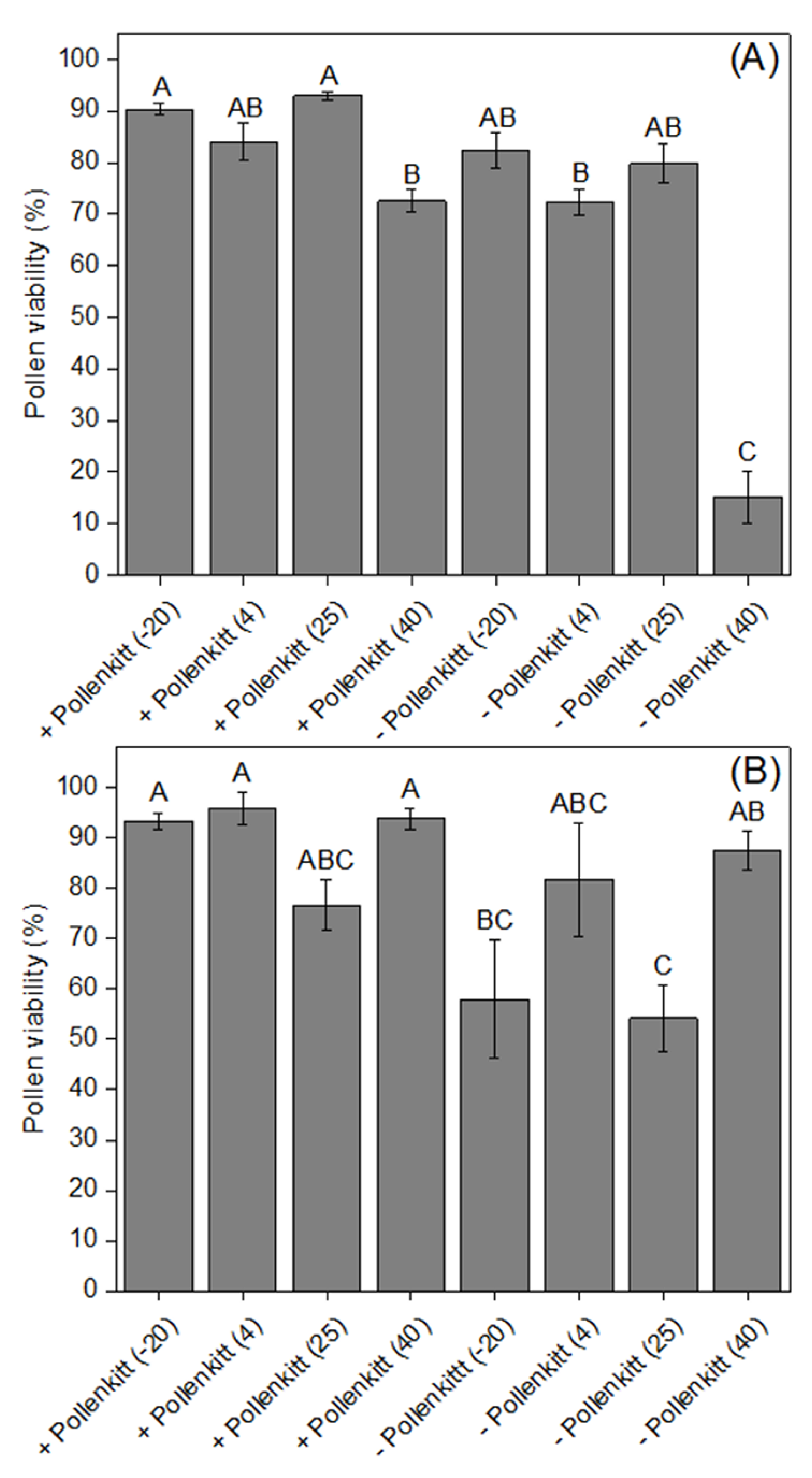
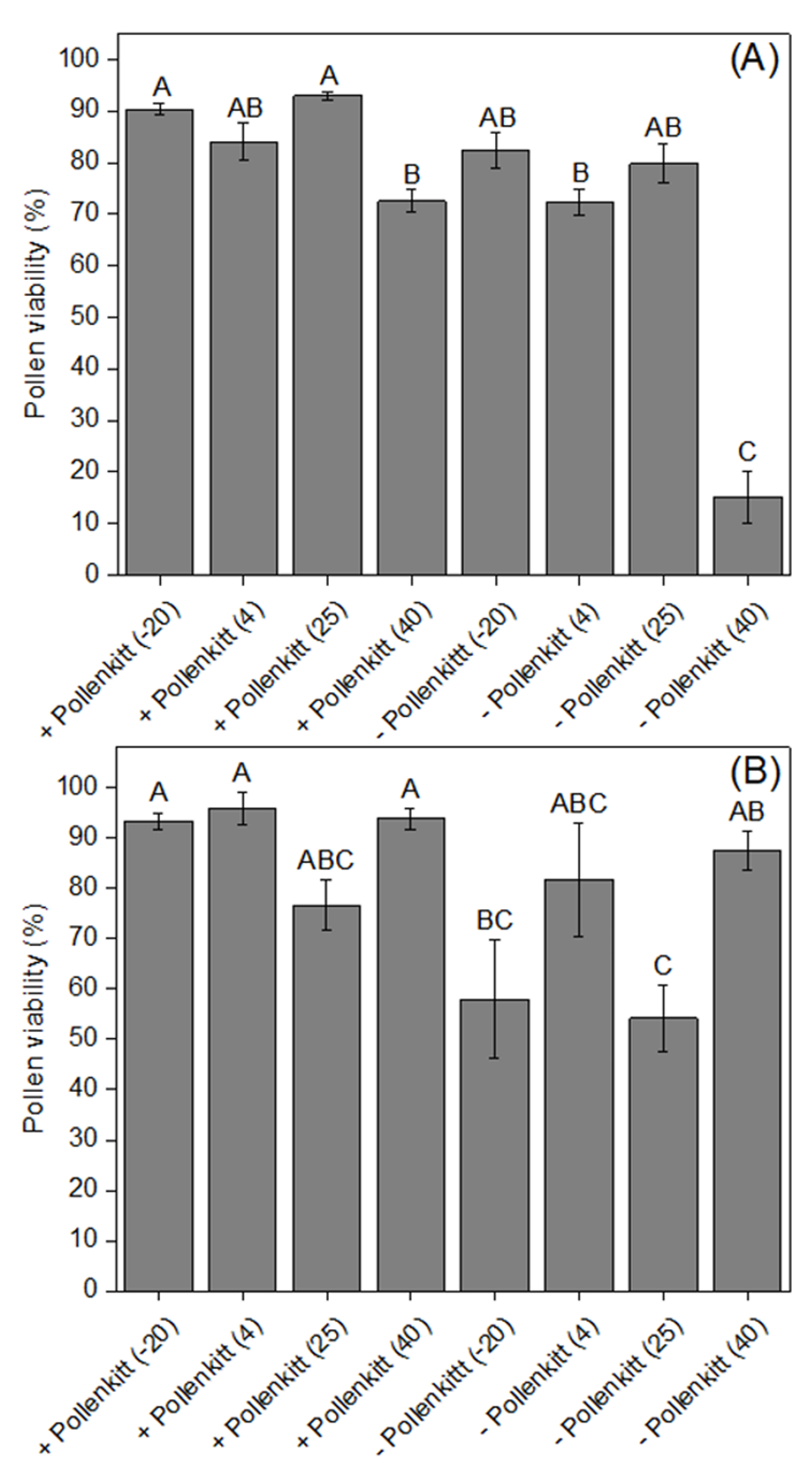
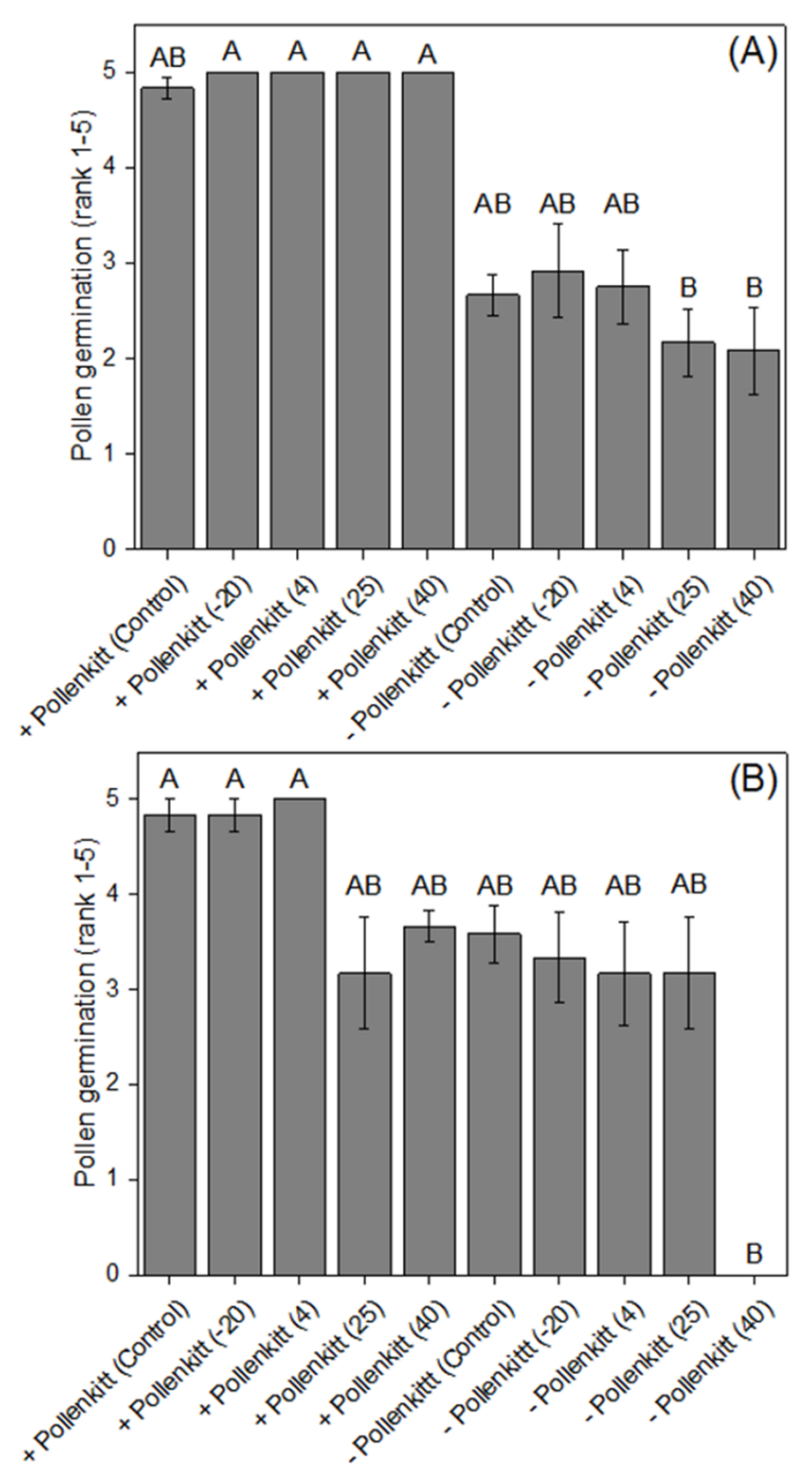
2.4. Effects of Temperature on Pollen Viability and Germinability, with or without Pollenkitt, inside the Stylar Tube
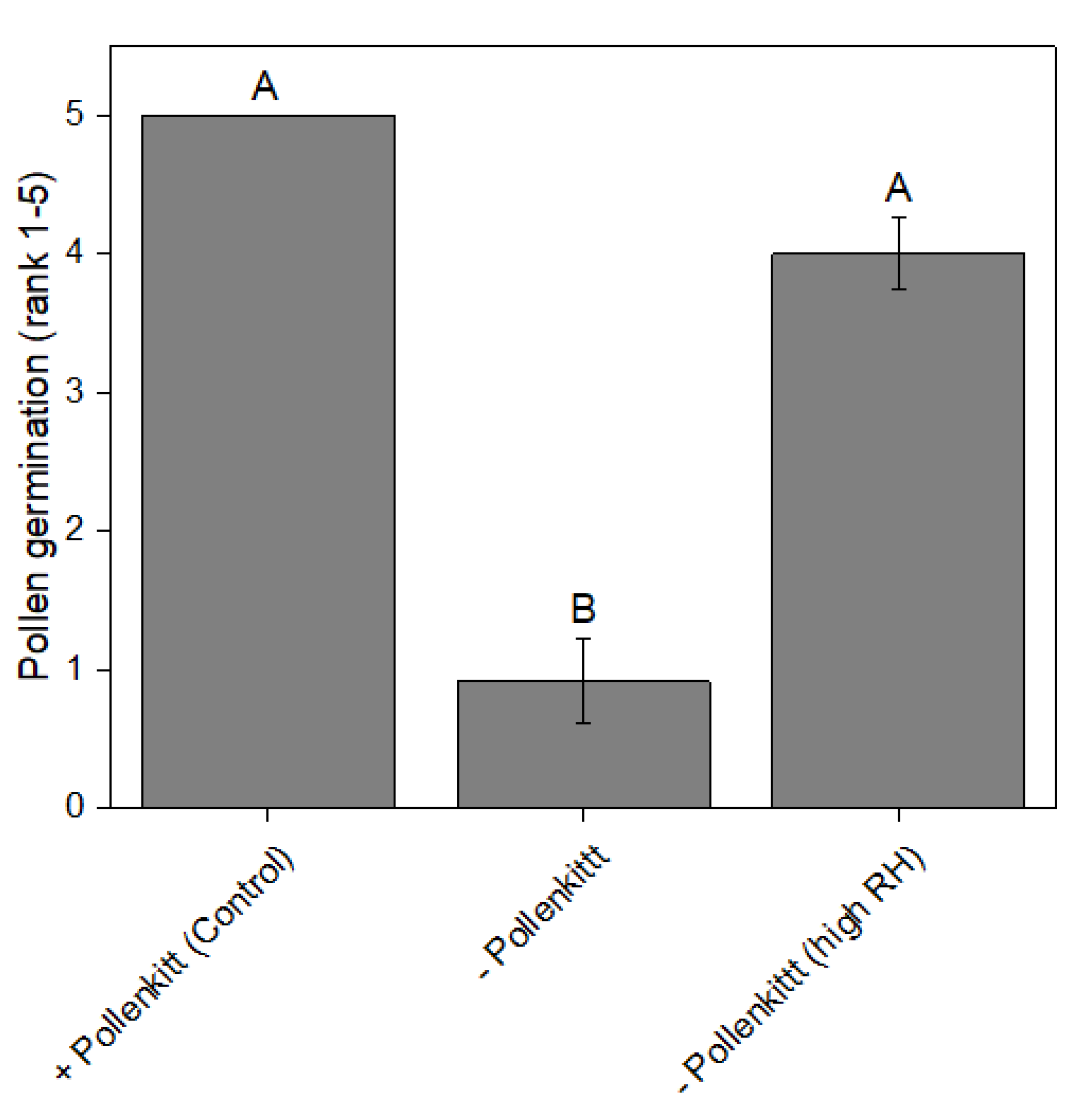
2.5. Test for Germinability under Low RH
2.6. Observation of Pollen on Anthers, Insects, and inside the Stylar Tube
2.7. Estimation of Weight Ratio of Pollenkitt to Pollen Grain
2.8. GC-MS Chemical Analysis of the Pollenkitt
2.9. Statistical Analyses
3. Results
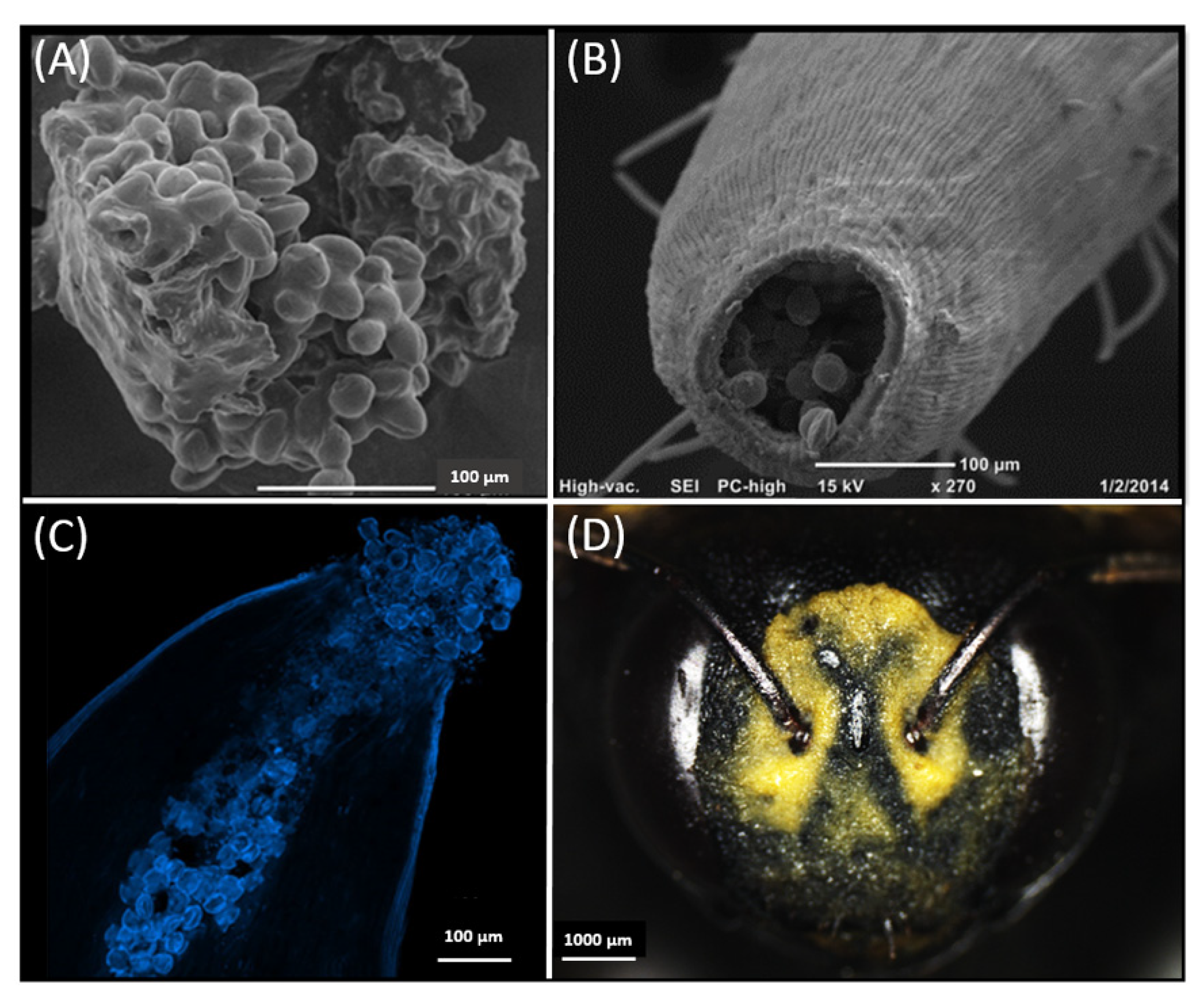
3.1. The Significance of the Pollenkitt for Pollen Insertion into the Stylar Tube
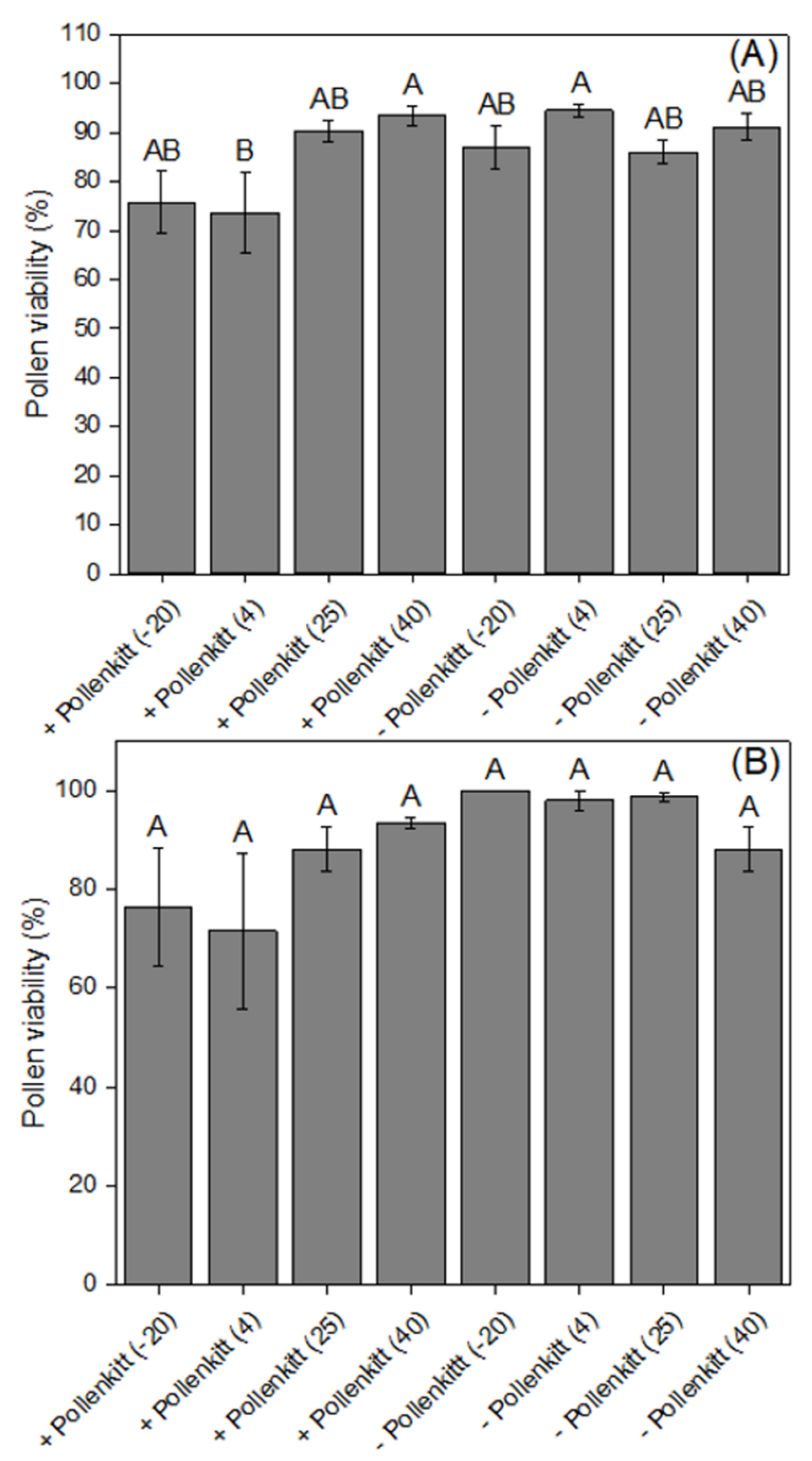
3.2. Effects of Temperatures on Pollen Viability and Pollen Germinability, with and without Pollenkitt
3.3. Effects of Extreme Temperatures on Pollen Germinability inside the Stylar Tube
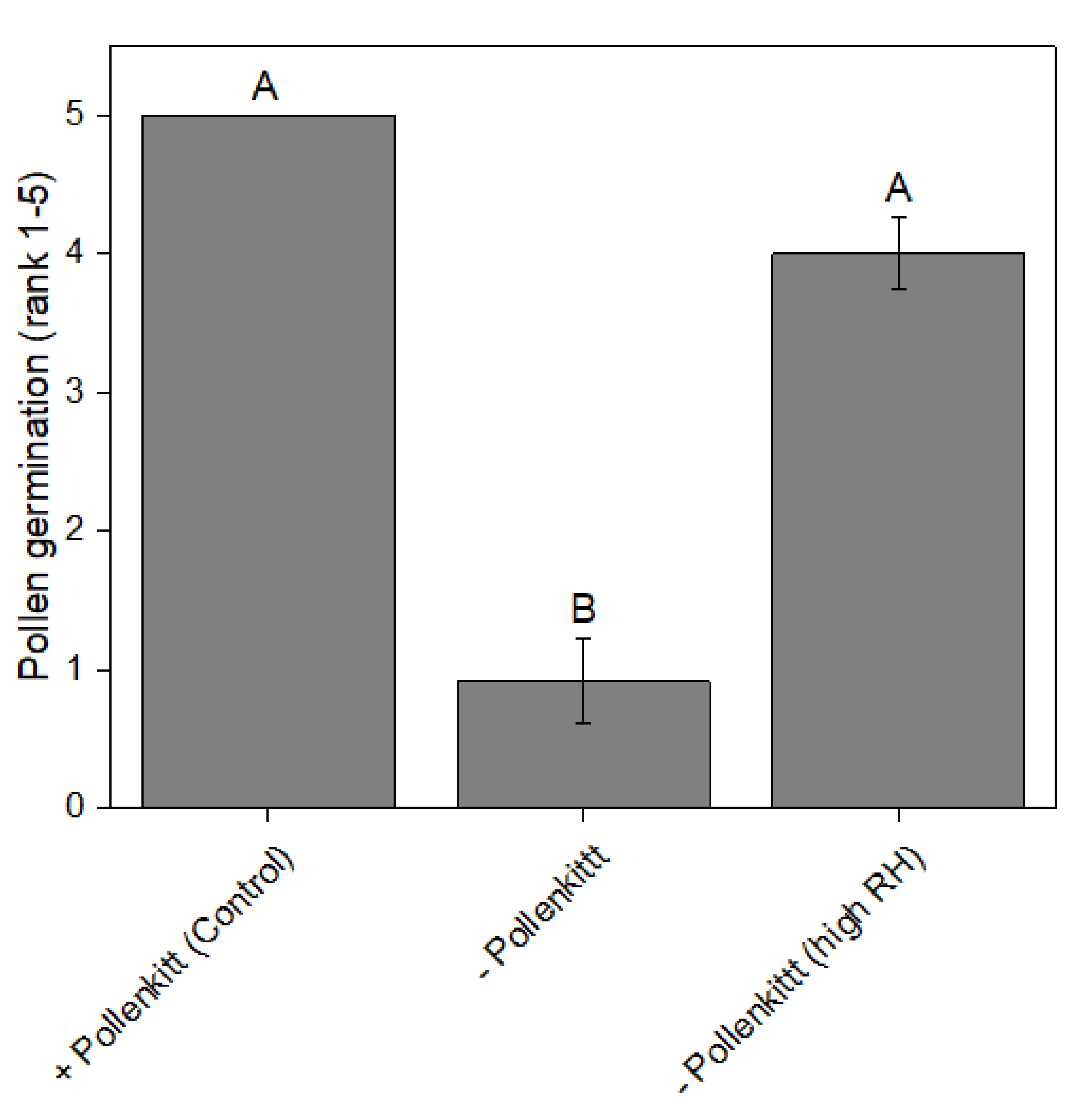
3.4. The Significance of the Pollenkitt for Germinability under Low RH
3.5. Morphological Analyses on Flowers and Insects and within the Stylar Tube
3.6. Estimation of Weight Ratio of the Pollenkitt to the Pollen Grains
3.7. GC-MS Chemical Analysis of the Pollenkitt
4. Discussion
4.1. Weight Ratio of Pollenkitt to the Pollen Grains
4.2. The Significance of the Pollenkitt for Pollen Insertion into the Stylar Tube
4.3. The Significance of the Pollenkitt for Protection against Heat and Water Stress
4.4. The Significance of the Stylar Tube for Protection against Heat and Water Stress
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olson, M.E.; Carlquist, S. Stem and root anatomical correlations with life form diversity, ecology, and systematics in Moringa (Moringaceae). Bot. J. Linn. Soc. 2001, 135, 315–348. [Google Scholar] [CrossRef]
- Ramachandran, C.; Peter, K.V.; Gopalakrishnan, P.K. Drumstick (Moringa oleifera): A multipurpose Indian vegetable. Econ. Bot. 1980, 34, 276–283. [Google Scholar] [CrossRef]
- Sengupta, A.; Gupta, M.P. Studies on the seed fat composition of Moringaceae family. Fett. Wiss. Technol. 1970, 72, 6–10. [Google Scholar] [CrossRef]
- Kleiman, R.; Ashley, D.A.; Brown, J.H. Comparison of two seed oils used in cosmetics, Moringa and Marula. Ind. Crop. Prod. 2008, 28, 361–364. [Google Scholar] [CrossRef]
- Rashid, U.; Anwar, F.; Moser, B.R.; Knothe, G. Moringa oleifera oil: A possible source of biodiesel. Bioresour. Technol. 2008, 99, 8175–8179. [Google Scholar] [CrossRef]
- Caceres, A.; Cabrera, O.; Morales, O.; Mollinedo, P.; Mendia, P. Pharmacological properties of Moringa oleifera. 1: Preliminary screening for antimicrobial activity. J. Ethnopharmacol. 1991, 33, 213–216. [Google Scholar] [CrossRef]
- Holst, S. Moringa: Nature’s Medicine Cabinet; Sierra Sunrise Books: Sherman Oaks, CA, USA, 2000; Volume 14622. [Google Scholar]
- Jahn, S.A.A.; Mussad, H.A.; Burgstaller, H. The tree that purifies water: Cultivating multipurpose Moringaceae in the Sudan. Unasylva 1986, 152, 23–28. [Google Scholar]
- Hegazy, A.K.; Hammouda, O.; Lovett-Doust, J.; Gomaa, N.H. Population dynamics of Moringa peregrina along altitudinal gradient in the northwestern sector of the Red Sea. J. Arid Environ. 2008, 72, 1537–1551. [Google Scholar] [CrossRef]
- Alaklabi, A. Genetic diversity of Moringa peregrina species in Saudi Arabia with ITS sequences. Saudi J. Biol. Sci. 2015, 22, 186–190. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, A. The Garden in Ancient Egypt; Rubicon Press: London, UK, 1998. [Google Scholar]
- Somali, M.A.; Bajneid, A.; Fhainian, S.S. Chemical composition and characteristic of Moringa peregrina seeds and seed oil. J. Am. Oil Chem. Soc. 1984, 16, 85–86. [Google Scholar] [CrossRef]
- Mridha, M.A.U. Prospects of Moringa cultivation in Saudi Arabia. J. Appl. Environ. Biol. Sci. 2015, 5, 39–46. [Google Scholar]
- Abud-Archila, M.; Espinosa-Arrioja, A.K.; González-Soto, T.; Gutiérrez-Oliva, V.F.; Ruíz-Valdiviezo, V.; González-Mendoza, D.; Rodríguez-Hernández, L.; Gutiérrez-Miceli, F.A. Growth and biochemical responses of Moringa (Moringa oleifera L.) to vermicompost and phosphate rock under water stress conditions. Phyton 2018, 87, 209–215. [Google Scholar]
- Vaknin, Y.; Mishal, A. The potential of the tropical “miracle tree” Moringa oleifera and its desert relative Moringa peregrina as edible seed-oil and protein crops under Mediterranean conditions. Sci. Hort. 2017, 225, 431–437. [Google Scholar] [CrossRef]
- Decraene, L.R.; De Laet, J.; Smets, E.F. Floral development and anatomy of Moringa oleifera (Moringaceae): What is the evidence for a capparalean or sapindalean affinity? Ann. Bot. 1998, 82, 273–284. [Google Scholar] [CrossRef]
- Jyothi, P.V.; Atluri, J.B.; Reddi, C.S. Pollination ecology of Moringa oleifera (Moringaceae). Proc. Plant Sci. 1990, 100, 33–42. [Google Scholar]
- Krieg, J.; Goetze, D.; Porembski, S.; Arnold, P.; Linsenmair, K.E.; Stein, K. Floral and reproductive biology of Moringa oleifera (Moringaceae) in Burkina Faso, West Africa. Acta Hortic. 2017, 1158, 63–69. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Mandal, S. Pollination, pollen germination and stigma receptivity in Moringa oleifera Lamk. Grana 2004, 43, 48–56. [Google Scholar] [CrossRef]
- Dafni, A.; Neppi, M.; Pacini, E. Pollen and stigma biology. In Practical Pollination Biology; Dafni, A., Kevan, P.G., Husband, B., Eds.; Enviroquest: Cambridge, ON, Canada, 2005; pp. 83–147. [Google Scholar]
- Evans, D.E.; Taylor, P.E.; Singh, M.B.; Knox, R.B. Quantitative analysis of lipids and protein from the pollen of Brassica napus L. Plant Sci. 1991, 73, 117–126. [Google Scholar] [CrossRef]
- Martin, F.W. Staining and observing pollen tubes in the style by means of fluorescence. Stain Technol. 1959, 34, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Middleton, N.; Thomas, D. (Eds.) World Atlas of Desertification, 2nd ed.; Hodder Headline Group: London, UK, 1997. [Google Scholar]
- Beaumont, P. Drylands: Environmental Management and Development; Routledge: London, UK; New York, NY, USA, 2014. [Google Scholar]
- Pacini, E.; Hesse, M. Pollenkitt—Its composition, forms and functions. Flora-Morphol. Distrib. Funct. Ecol. Plants 2005, 200, 399–415. [Google Scholar] [CrossRef]
- Pacini, E.; Franchi, G.G. Some cytological, ecological and evolutionary aspects of pollination. Acta Soc. Bot. Pol. 1996, 65, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Heslop-Harrison, J. An interpretation of the hydrodynamics of pollen. Am. J. Bot. 1979, 1, 737–743. [Google Scholar] [CrossRef]
- Haisheng, L.; Gomez, I.; Meredith, J.C. Pollenkitt wetting mechanism enables species-specific tunable pollen adhesion. Langmuir 2013, 29, 3012–3023. [Google Scholar]
- Galati, B.G.; Rosenfeld, S. The pollen development in Ceiba insignis (Kunth) Gibbs & Semir ex Chorisia speciosa St. Hil. (Bombacaceae). Phytomorphology 1998, 48, 121–130. [Google Scholar]
- Hodge, W.H.; Sineath, H.H. The Mexican candelilla plant and its wax. Econ. Bot. 1956, 10, 134–154. [Google Scholar] [CrossRef]
- Saradhi, P.P.; Ram, H.M. Some Aspects of floral biology of Cassia fistula Linn. (The Indian laburnum)—Part I. Curr. Sci. 1981, 50, 802–805. [Google Scholar]
- Bourdenx, B.; Bernard, A.; Domergue, F.; Pascal, S.; Léger, A.; Roby, D.; Pervent, M.; Vile, D.; Haslam, R.P.; Napier, J.A.; et al. Overexpression of Arabidopsis ECERIFERUM1 promotes wax very-long-chain alkane biosynthesis and influences plant response to biotic and abiotic stresses. Plant Physiol. 2011, 156, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Richards, R.A.; Rawson, H.M.; Johnson, D.A. Glaucousness in wheat: Its development and effect on water—Use efficiency, gas exchange and photosynthetic tissue temperatures. Aust. J. Plant Physiol. 1986, 13, 465–473. [Google Scholar] [CrossRef]
- Bennett, D.; Izanloo, A.; Edwards, J.; Kuchel, H.; Chalmers, K.; Tester, M.; Reynolds, M.P.; Schnurbusch, T.; Langridge, P. Identification of novel quantitative trait loci for days to ear emergence and flag leaf glaucousness in bread wheat (Triticum aestivum L.) population adapted to southern Australian conditions. Theor. Appl. Genet. 2012, 124, 697–711. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Mason, R.E.; Huggins, T.; Hays, D.B. QTL on wheat (Triticum aestivum L.) chromosomes 1B, 3D and 5A are associated with constitutive production of leaf cuticular wax and may contribute to lower leaf temperatures under heat stress. Euphytica 2015, 201, 123–130. [Google Scholar] [CrossRef]
- Gefen, E.; Talal, S.; Brendzel, O.; Dror, A.; Fishman, A. Variation in quantity and composition of cuticular hydrocarbons in the scorpion Buthus occitanus (Buthidae) in response to acute exposure to desiccation stress. Comp. Biochem. Phys. A 2015, 182, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Krupp, J.J.; Nayal, K.; Wong, A.; Millar, J.G.; Levine, J.D. Desiccation resistance is an adaptive life-history trait dependent upon cuticular hydrocarbons, and influenced by mating status and temperature in D. melanogaster. J. Insect Physiol. 2020, 121, 103990. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, S.; Ewalda, A.; Hellwig, F. The Anatomy of the stigma and style from Cyclamen persicum (Mill.) cv. “pure white” and its relation to pollination success. Plant Biol. 2007, 9, 158–162. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Name | Molecular Formula | Retention Time | M. oleifera | M. peregrina |
|---|---|---|---|---|
| Trans-nerolidol | C15H26O | 12.321 | 1.22 ± 0.31 | --- |
| 9-methyl-nonadecane | C20H32 | 14.859 | --- | 0.54 ± 0.29 |
| Palmitic acid | C16H32O2 | 15.138 | 1.32 ± 0.38 | 1.51 ± 0.75 |
| Oleic acid | C18H34O2 | 16.268 | --- | 0.42 |
| Tricosane | C23H48 | 17.228 | 3.48 ± 0.12 | 5.54 ± 16. |
| Tetracosane | C24H50 | 17.78 | 1.22 ± 0.21 | 1.8 ± 0.18 |
| N.I, long chain | Aldehyde | 18.129 | --- | 0.65 ± 0.15 |
| Pentacosane * | C25H52 | 18.327 | 23.74 ± 0.05 | 27.98 ± 3.87 |
| N.I, long chain | Aldehyde | 18.432 | --- | 0.45 |
| Hexacosane | C26H54 | 18.828 | 2.31 ± 0.31 | 2.27 ± 0.13 |
| N.I, long chain | Aldehyde | 19.134 | --- | 0.58 ± 0.18 |
| Heptacosane * | C27H56 | 19.392 | 24.34 ± 0.08 | 20.22 ± 1.39 |
| Octacosane | C28H58 | 19.975 | 1.23 ± 1.26 | 0.68 |
| N.I* long chain | Aldehyde | 20.085 | --- | 3.97 ± 1.48 |
| Nonacosane * | C29H60 | 20.678 | 18.50 ± 0.02 | 17.08 ± 0.61 |
| Triacontane | C30H62 | 21.455 | 1.33 ± 0.88 | 0.94 ± 0.12 |
| γ-Tocopherol | C28H48O2 | 21.995 | 1.41 ± 0.06 | 0.36 |
| Hentriacontane * | C31H64 | 22.414 | 15.95 ± 0.1 | 14.92 ± 2.15 |
| α-Tocopherol | C29H50O2 | 22.788 | 1.37 ± 0.68 | --- |
| Tritriacontane | C33H68 | 24.865 | 1.91 ± 0.13 | --- |
| γ-Sitosterol | C29H50O | 25.297 | 0.68 ± 0.26 | --- |
| Total | 100 (82.53 *) | 100 (80.2 *) |
| Chemical Name | Molecular Formula | Retention Time | M. oleifera | M. peregrina |
|---|---|---|---|---|
| Nonanal | C9H18O | 8.791 | √ | √ |
| Trans-2-nonenal | C9H16O | 9.377 | √ | ----- |
| 3′-Methylacetophenone | C9H10O | 9.707 | √ | ----- |
| α-Terpineol | C10H18O | 9.791 | ----- | √ |
| Decanal | C10H20O | 9.831 | √ | √ |
| α-Ionol | C13H22O | 10.256 | √ | ----- |
| 3-Heptylacrolein | C10H18O | 10.386 | √ | √ |
| Undecanal | C11H22O | 10.8 | √ | √ |
| 2,4-Decadienal | C10H16O | 10.928 | √ | √ |
| 2-Undecenal | C11H20O | 11.324 | √ | √ |
| Myrcene | C10H16 | 11.402 | √ | ----- |
| Methyl cinnamate | C10H10O2 | 11.561 | √ | √ |
| Dodecyl aldehyde | C12H24O | 11.71 | √ | √ |
| β-Bisabolene | C15H24 | 12.987 | √ | √ |
| Tetra decyl aldehyde | C14H28O | 13.379 | √ | √ |
| 2-undecanol | C14H28O | 13.969 | √ | √ |
| Pentadecanal | C15H30O | 14.148 | √ | √ |
| 9-methylnonadecane | C20H42 | 15.691 | √ | √ |
| Palmitic acid | C16H32O2 | 15.797 | √ | √ |
| δ-tetradecalactone | C14H26O2 | 16.99 | √ | ----- |
| Stearic acid | C18H36O2 | 17.06 | √ | √ |
| Bisphenol A | C15H16O2 | 17.239 | √ | √ |
| Tricosane | C23H48 | 18.028 | √ | √ |
| δ-Stearolactone | C18H34O2 | 18.223 | √ | ----- |
| Tetracosane | C24H50 | 18.441 | √ | √ |
| Pentacosane | C25H52 | 18.971 | √ | √ |
| Hexacosane | C26H54 | 19.485 | √ | √ |
| Heptacosane | C27H56 | 20.029 | √ | √ |
| Octacosane | C28H58 | 20.63 | √ | √ |
| Nonacosane | C29H60 | 21.312 | √ | √ |
| Triacontane | C30H62 | 22.106 | √ | ----- |
| Hentriacontane | C31H64 | 23.04 | √ | ----- |
| Dotriacontane | C32H66 | 24.156 | √ | ----- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaknin, Y.; Eisikowitch, D.; Mishal, A. Floral and Pollen Traits of Moringa oleifera Lam. and Moringa peregrina (Forssk.) Fiori Provide Reproductive Adaptations for Arid Conditions. Agronomy 2021, 11, 1090. https://doi.org/10.3390/agronomy11061090
Vaknin Y, Eisikowitch D, Mishal A. Floral and Pollen Traits of Moringa oleifera Lam. and Moringa peregrina (Forssk.) Fiori Provide Reproductive Adaptations for Arid Conditions. Agronomy. 2021; 11(6):1090. https://doi.org/10.3390/agronomy11061090
Chicago/Turabian StyleVaknin, Yiftach, Dan Eisikowitch, and Adina Mishal. 2021. "Floral and Pollen Traits of Moringa oleifera Lam. and Moringa peregrina (Forssk.) Fiori Provide Reproductive Adaptations for Arid Conditions" Agronomy 11, no. 6: 1090. https://doi.org/10.3390/agronomy11061090
APA StyleVaknin, Y., Eisikowitch, D., & Mishal, A. (2021). Floral and Pollen Traits of Moringa oleifera Lam. and Moringa peregrina (Forssk.) Fiori Provide Reproductive Adaptations for Arid Conditions. Agronomy, 11(6), 1090. https://doi.org/10.3390/agronomy11061090