
Mineral and Antioxidant Attributes of Petroselinum crispum at Different Stages of Ontogeny: Microgreens vs. Baby Greens

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Growth Chamber and Experiment Arrangement
2.2. Parsley Yield, Sampling and Macronutrients Analysis
2.3. Chlorophylls Pigments, Total Ascorbic Acid and Hydrophilic Antioxidant Activity Assessment
2.4. Carotenoids and Polyphenols Extraction and Quantification
2.5. Statistical Analysis
3. Results and Discussion
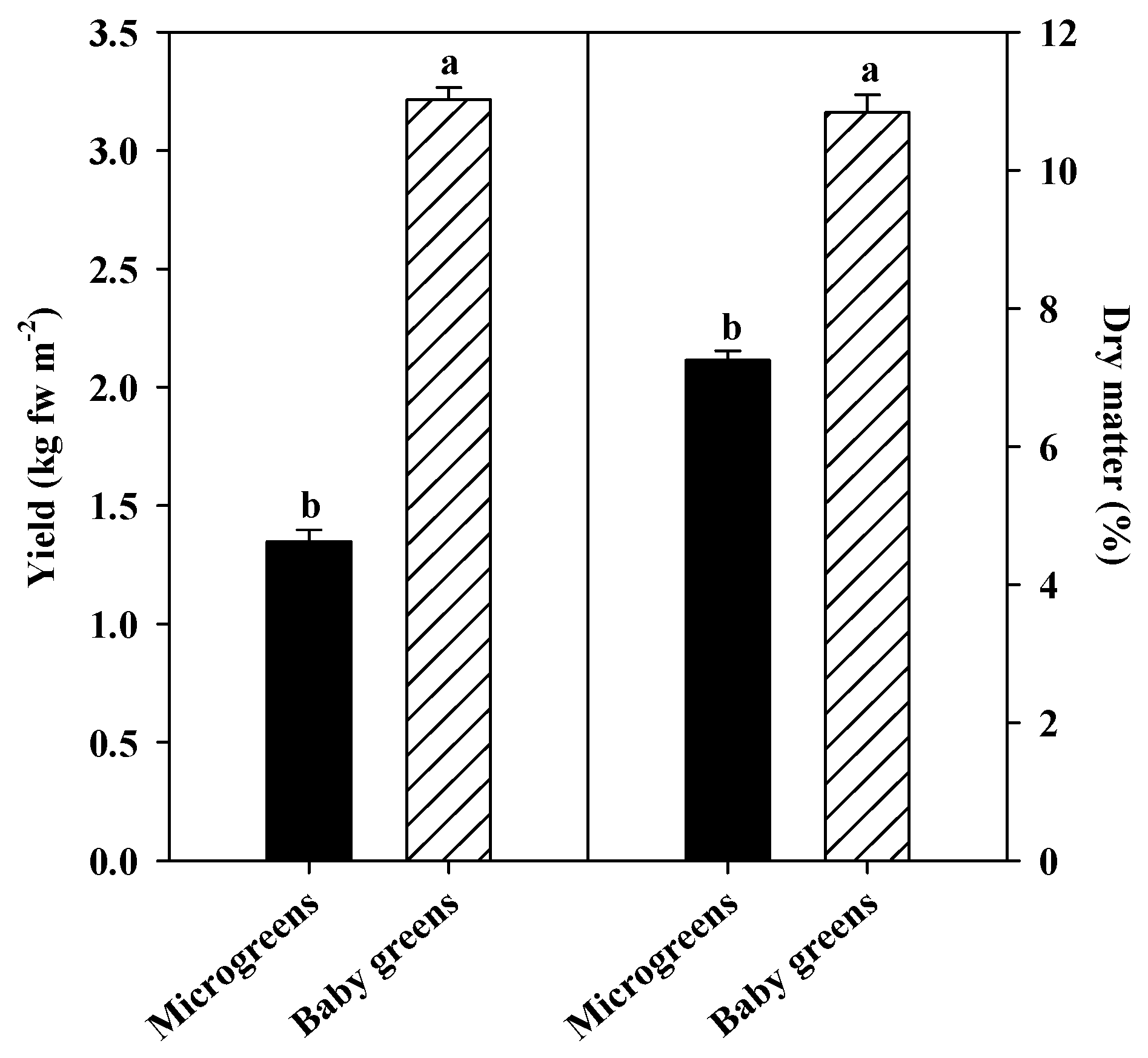
3.1. Yield, Dry Matter and Macronutrient Content
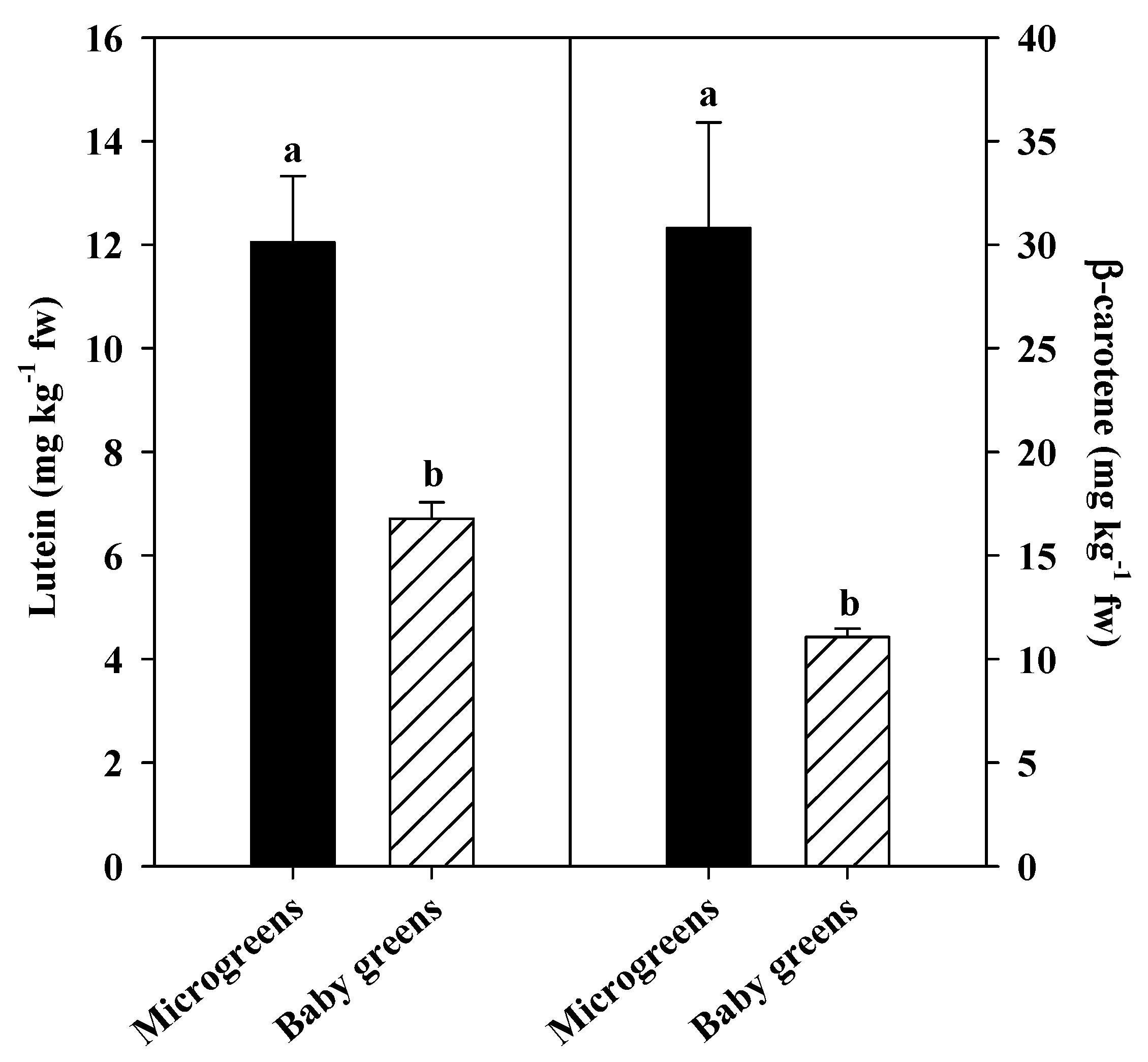
3.2. Chlorophyll and Carotenoids Pigments
3.3. Total Ascorbic Acid, Polyphenols and Hydrophilic Antioxidant Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keys, A. Mediterranean diet and public health: Personal reflections. Am. J. Clin. Nutr. 1995, 61, 1321S–1323S. [Google Scholar] [CrossRef] [PubMed]
- Bower, A.; Marquez, S.; de Mejia, E.G. The Health Benefits of Selected Culinary Herbs and Spices Found in the Traditional Mediterranean Diet. Crit. Rev. Food Sci. Nutr. 2016, 56, 2728–2746. [Google Scholar] [CrossRef] [PubMed]
- Nestle, M. Mediterranean diets: Historical and research overview. Am. J. Clin. Nutr. 1995, 61, 1313S–1320S. [Google Scholar] [CrossRef]
- Mara De Menezes Epifanio, N.; Rykiel Iglesias Cavalcanti, L.; Falcão Dos Santos, K.; Soares Coutinho Duarte, P.; Kachlicki, P.; Ozarowski, M.; Jorge Riger, C.; Siqueira De Almeida Chaves, D. Chemical characterization and in vivo antioxidant activity of parsley (Petroselinum crispum) aqueous extract. Food Funct. 2020, 11, 5346–5356. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, Â.; Polyzos, N.; Petropoulos, S.A.; Pinela, J.; Ardohain, E.; Moreira, G.; Ferreira, I.C.F.R.; Barros, L. Phytochemical composition and nutritional value of pot-grown turnip-rooted and plain and curly-leafed parsley cultivars. Agronomy 2020, 10, 1416. [Google Scholar] [CrossRef]
- Tang, E.L.H.; Rajarajeswaran, J.; Fung, S.; Kanthimathi, M.S. Petroselinum crispum has antioxidant properties, protects against DNA damage and inhibits proliferation and migration of cancer cells. J. Sci. Food Agric. 2015, 95, 2763–2771. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, F.; Wang, X.; Yao, H.Y. Evaluation of antioxidant activity of parsley (Petroselinum crispum) essential oil and identification of its antioxidant constituents. Food Res. Int. 2006, 39, 833–839. [Google Scholar] [CrossRef]
- Kreydiyyeh, S.I.; Usta, J.; Kaouk, I.; Al-Sadi, R. The mechanism underlying the laxative properties of parsley extract. Phytomedicine 2001, 8, 382–388. [Google Scholar] [CrossRef]
- Kilic, C.; Duyar, H. A Research on Production of Baby Leaf Vegetables in Floating System. Hung. Agric. Eng. 2016, 29, 24–27. [Google Scholar]
- Jadczak, D.; Bojko, K.; Wysocka, G.; Szymańska, M. Yield and biological properties of leaf parsley (Petroselinum crispum (mill.) Nym. Ex A.W. Hillc Convar. crispum). J. Elem. 2019, 24, 803–815. [Google Scholar] [CrossRef]
- Dorman, H.J.D.; Lantto, T.A.; Raasmaja, A.; Hiltunen, R. Antioxidant, pro-oxidant and cytotoxic properties of parsley. Food Funct. 2011, 2, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Samuoliene, G.; Brazaityte, A.; Viršile, A.; Jankauskiene, J.; Sakalauskiene, S.; Duchovskis, P. Red light-dose or wavelength-dependent photoresponse of antioxidants in herb microgreens. PLoS ONE 2016, 11, e0163405. [Google Scholar] [CrossRef] [PubMed]
- Saffaryazdi, A.; Ganjeali, A.; Farhoosh, R.; Cheniany, M. Variation in phenolic compounds, α-linolenic acid and linoleic acid contents and antioxidant activity of purslane (Portulaca oleracea L.) during phenological growth stages. Physiol. Mol. Biol. Plants 2020, 26, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Buturi, C.V.; Mauro, R.P.; Fogliano, V.; Leonardi, C.; Giuffrida, F. Mineral Biofortification of Vegetables as a Tool to Improve Human Diet. Foods 2021, 10, 223. [Google Scholar] [CrossRef]
- Di Gioia, F.; Renna, M.; Santamaria, P. Sprouts, Microgreens and “Baby Leaf” Vegetables; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Lenzi, A.; Orlandini, A.; Bulgari, R.; Ferrante, A.; Bruschi, P. Antioxidant and mineral composition of three wild leafy species: A comparison between microgreens and baby greens. Foods 2019, 8, 487. [Google Scholar] [CrossRef]
- Di Bella, M.C.; Niklas, A.; Toscano, S.; Picchi, V.; Romano, D.; Lo Scalzo, R.; Branca, F. Morphometric characteristics, polyphenols and ascorbic acid variation in Brassica oleracea L. novel foods: Sprouts, microgreens and baby leaves. Agronomy 2020, 10, 782. [Google Scholar] [CrossRef]
- Treadwell, D.; Hochmuth, R.; Landrum, L.; Laughlin, W. Microgreens: A New Specialty Crop. Edis 2020, 2020, hs1164–hs2020. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Soteriou, G.A.; Giordano, M.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Genotype-Specific Modulatory Effects of Select Spectral Bandwidths on the Nutritive and Phytochemical Composition of Microgreens. Front. Plant Sci. 2019, 10, 1501. [Google Scholar] [CrossRef] [PubMed]
- Pannico, A.; El-Nakhel, C.; Kyriacou, M.C.; Giordano, M.; Stazi, S.R.; De Pascale, S.; Rouphael, Y. Combating Micronutrient Deficiency and Enhancing Food Functional Quality Through Selenium Fortification of Select Lettuce Genotypes Grown in a Closed Soilless System. Front. Plant Sci. 2019, 10, 1495. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Van Montagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef]
- Kim, H.J.; Fonseca, J.M.; Choi, J.H.; Kubota, C.; Dae, Y.K. Salt in irrigation water affects the nutritional and visual properties of romaine lettuce (Lactuca sativa L.). J. Agric. Food Chem. 2008, 56, 3772–3776. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef]
- Duyar, H.; Kiliç, C.C. A Research on Production of Rocket and Parsley in Floating System. J. Agric. Sci. 2016, 8, 54. [Google Scholar] [CrossRef]
- Waterland, N.L.; Moon, Y.; Tou, J.C.; Kim, M.J.; Pena-Yewtukhiw, E.M.; Park, S. Mineral Content Differs among Microgreen, Baby Leaf, and Adult Stages in Three Cultivars of Kale. Hort Sci. 2017, 52, 566–571. [Google Scholar] [CrossRef]
- Vaštakaitė-Kairienė, V.; Brazaitytė, A.; Viršilė, A.; Samuolienė, G.; Miliauskienė, J.; Jankauskienė, J.; Novičkovas, A.; Duchovskis, P. The nutritional value of Brassica leafy greens in different growth stages. Acta Hortic. 2020, 1271, 455–464. [Google Scholar] [CrossRef]
- Kopsell, D.E.; Kopsell, D.A.; Sams, C.E.; Barickman, T.C. Ratio of Calcium To Magnesium Influences Biomass, Elemental Accumulations, and Pigment Concentrations in Kale. J. Plant Nutr. 2013, 36, 2154–2165. [Google Scholar] [CrossRef]
- Bindraban, P.S.; Dimkpa, C.; Nagarajan, L.; Roy, A.; Rabbinge, R. Revisiting fertilisers and fertilisation strategies for improved nutrient uptake by plants. Biol. Fertil. Soils 2015, 51, 897–911. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Variation in macronutrient content, phytochemical constitution and in vitro antioxidant capacity of green and red butterhead lettuce dictated by different developmental stages of harvest maturity. Antioxidants 2020, 9, 300. [Google Scholar] [CrossRef]
- Kizhedath, A.; Suneetha, V. Estimation of chlorophyll content in common household medicinal leaves and their utilization to avail health benefits of chlorophyll. J. Pharm. Res. 2011, 4, 1412–1413. [Google Scholar]
- Kimura, M.; Rodriguez-Amaya, D.B. Carotenoid composition of hydroponic leafy vegetables. J. Agric. Food Chem. 2003, 51, 2603–2607. [Google Scholar] [CrossRef]
- Pannico, A.; Graziani, G.; El-Nakhel, C.; Giordano, M.; Ritieni, A.; Kyriacou, M.C.; Rouphael, Y. Nutritional stress suppresses nitrate content and positively impacts ascorbic acid concentration and phenolic acids profile of lettuce microgreens. Italus Hortus 2020, 27, 41–52. [Google Scholar] [CrossRef]
- Hirschmann, J.V.; Raugi, G.J. Adult scurvy. J. Am. Acad. Dermatol. 1999, 41, 895–910. [Google Scholar] [CrossRef]
- Al-Juhaimi, F.; Ghafoor, K. Total phenols and antioxidant activities of leaf and stem extracts from coriander, mint and parsley grown in Saudi Arabia. Pak. J. Bot. 2011, 43, 2235–2237. [Google Scholar]



{kind=link}
{kind=link}
| Harvest Stage | Nitrate | P | K | Ca | Mg | S | Na |
|---|---|---|---|---|---|---|---|
| (mg kg−1 fw) | (mg kg−1 fw) | (mg kg−1 fw) | (mg kg−1 fw) | (mg kg−1 fw) | (mg kg−1 fw) | (mg kg−1 fw) | |
| Microgreens | 3362 ± 240 | 393.6 ± 1.44 | 3843 ± 26.9 | 467.7 ± 11.8 | 373.6 ± 4.31 | 249.1 ± 11.1 | 474.1 ± 12.4 |
| Baby greens | 1161 ± 27 | 270.9 ± 2.47 | 3677 ± 35.7 | 689.7 ± 19.6 | 597.0 ± 28.1 | 252.2 ± 6.51 | 920.8 ± 23.6 |
| Significance | *** | *** | * | *** | *** | ns | *** |
| Harvest Stage | HAA | TAA | Chlorophyll a | Chlorophyll b | Total Chlorophylls |
|---|---|---|---|---|---|
| (mmol AA eq. 100 g−1 fw) | (mg AA 100 g−1 fw) | (mg g−1 fw) | (mg g−1 fw) | (mg g−1 fw) | |
| Microgreens | 0.91 ± 0.03 | 20.26 ± 0.63 | 10.68 ± 0.18 | 2.91 ± 0.05 | 13.59 ± 0.13 |
| Baby greens | 2.46 ± 0.09 | 57.45 ± 10.75 | 14.04 ± 0.80 | 4.32 ± 0.31 | 18.36 ± 0.88 |
| Significance | *** | * | * | * | ** |
| Polyphenols (µg g−1 fw) | Harvest Stage | Significance | |
|---|---|---|---|
| Microgreens | Baby Greens | ||
| Apigenin | 0.15 ± 0.00 | 0.22 ± 0.01 | *** |
| Apigenin-7-apiosyl-glucoside | 369.4 ± 3.52 | 537.8 ± 3.54 | *** |
| Apigenin-7-O-glucoside | 8.56 ± 0.20 | 24.61 ± 0.88 | *** |
| Apigenin-malonyl-apiosyl-glucoside | 363.6 ± 3.31 | 650.0 ± 30.9 | *** |
| Caffeic acid | 0.28 ± 0.00 | 0.41 ± 0.00 | *** |
| Chlorogenic acid | 1.01 ± 0.00 | 0.58 ± 0.01 | *** |
| Chrysoeriol | 3.47 ± 0.24 | 8.22 ± 0.19 | *** |
| Coumaric acid | 2.27 ± 0.01 | 3.50 ± 0.02 | *** |
| Ferulic acid | 0.26 ± 0.00 | 0.39 ± 0.00 | *** |
| Kaempferol-7-O-glucoside | 0.17 ± 0.02 | 0.15 ± 0.02 | ns |
| Luteolin-7-O-glucoside | 0.53 ± 0.01 | 1.86 ± 0.07 | *** |
| Quercetin-3-O-galactoside | 0.47 ± 0.03 | 4.38 ± 0.12 | *** |
| Total polyphenols | 750.2 ± 2.05 | 1232.0 ± 33.7 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Nakhel, C.; Pannico, A.; Graziani, G.; Giordano, M.; Kyriacou, M.C.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Mineral and Antioxidant Attributes of Petroselinum crispum at Different Stages of Ontogeny: Microgreens vs. Baby Greens. Agronomy 2021, 11, 857. https://doi.org/10.3390/agronomy11050857
El-Nakhel C, Pannico A, Graziani G, Giordano M, Kyriacou MC, Ritieni A, De Pascale S, Rouphael Y. Mineral and Antioxidant Attributes of Petroselinum crispum at Different Stages of Ontogeny: Microgreens vs. Baby Greens. Agronomy. 2021; 11(5):857. https://doi.org/10.3390/agronomy11050857
Chicago/Turabian StyleEl-Nakhel, Christophe, Antonio Pannico, Giulia Graziani, Maria Giordano, Marios C. Kyriacou, Alberto Ritieni, Stefania De Pascale, and Youssef Rouphael. 2021. "Mineral and Antioxidant Attributes of Petroselinum crispum at Different Stages of Ontogeny: Microgreens vs. Baby Greens" Agronomy 11, no. 5: 857. https://doi.org/10.3390/agronomy11050857
APA StyleEl-Nakhel, C., Pannico, A., Graziani, G., Giordano, M., Kyriacou, M. C., Ritieni, A., De Pascale, S., & Rouphael, Y. (2021). Mineral and Antioxidant Attributes of Petroselinum crispum at Different Stages of Ontogeny: Microgreens vs. Baby Greens. Agronomy, 11(5), 857. https://doi.org/10.3390/agronomy11050857