
Can Grazing Moderate Climatic Effects on Herbage Nutritional Quality?



{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Treatments
2.3. Vegetation Sampling
2.4. Data Analysis
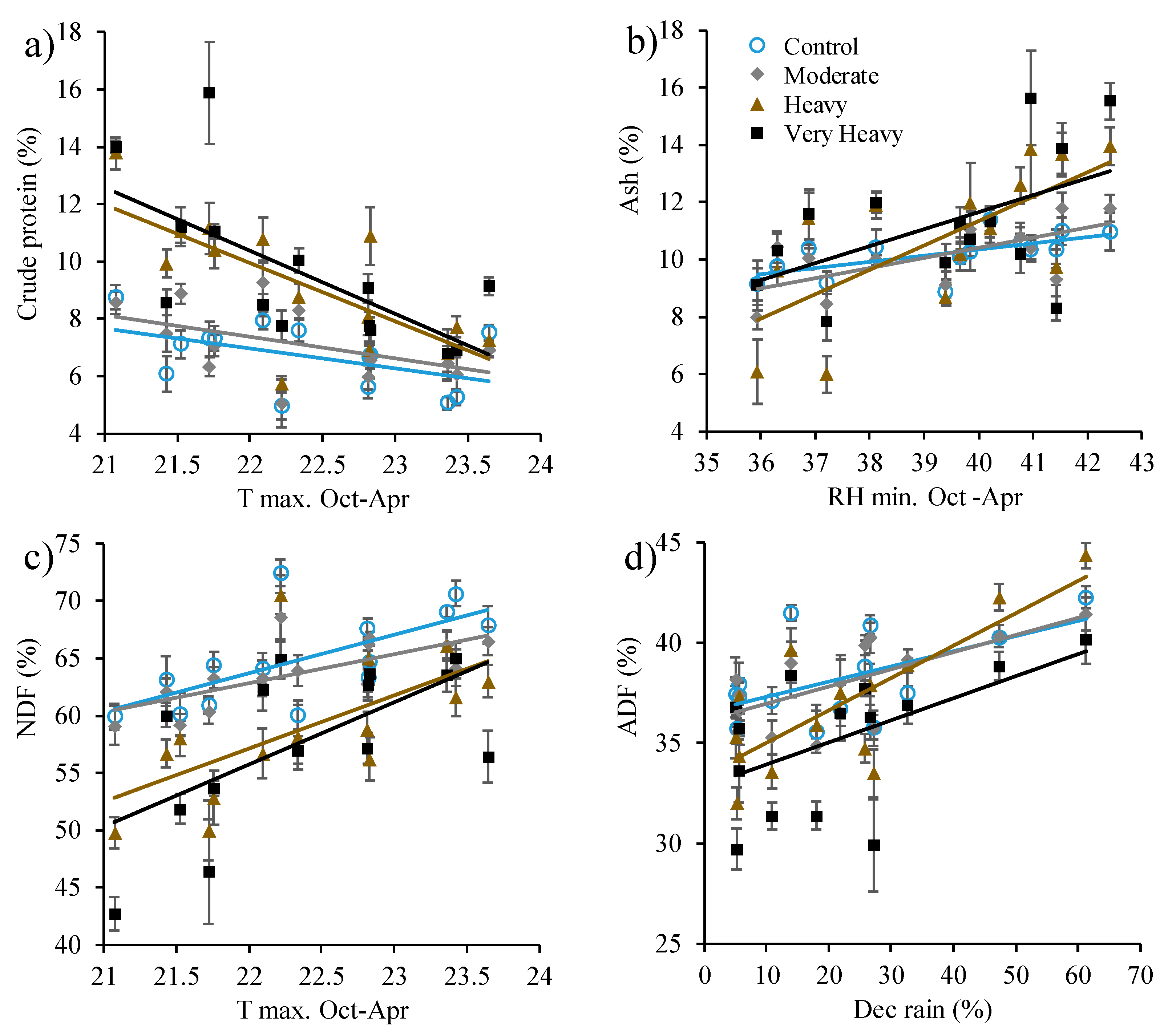
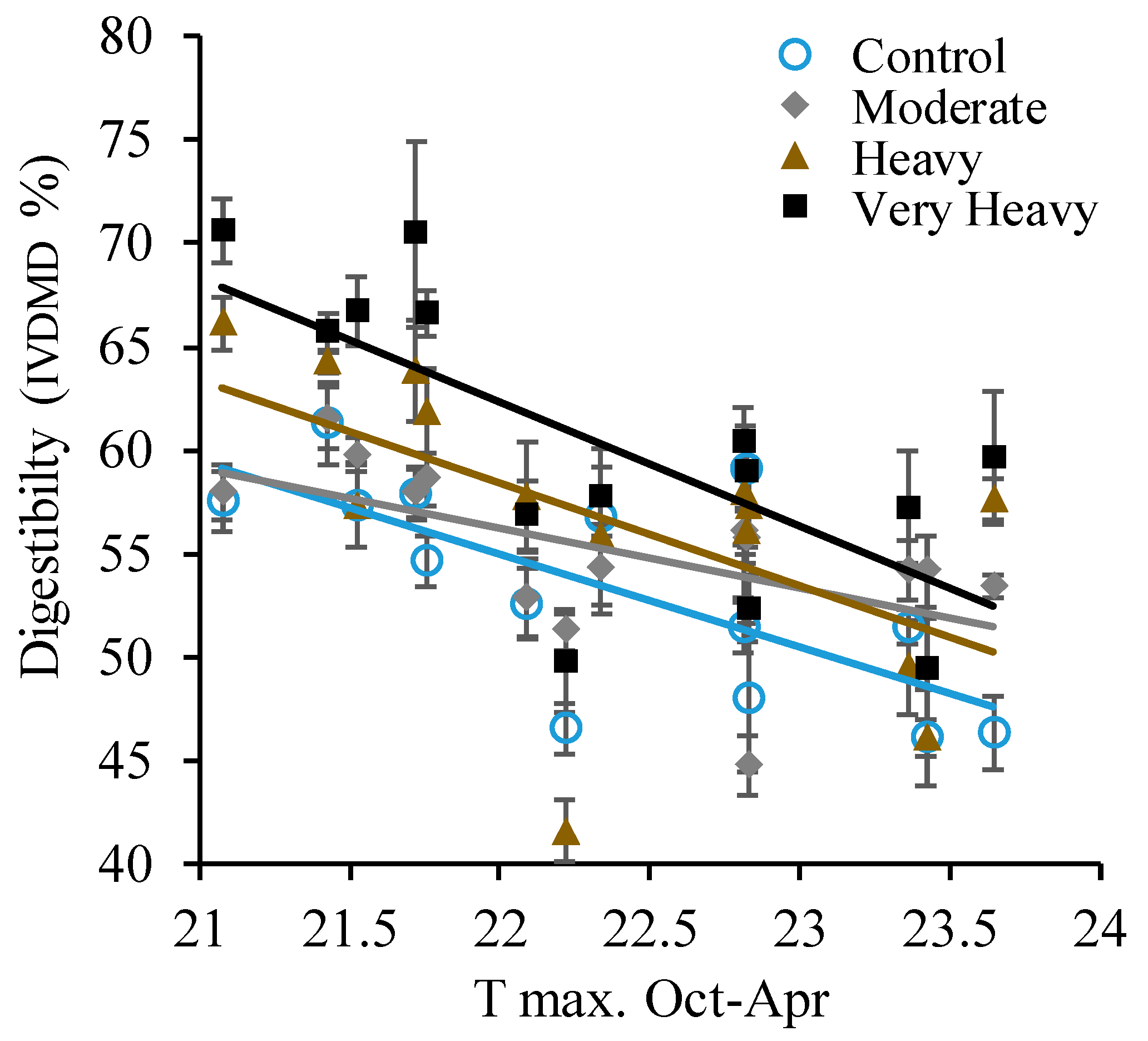
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapin, F.S., III; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Boone, R.B.; Conant, R.T.; Sircely, J.; Thornton, P.K.; Herrero, M. Climate change impacts on selected global rangeland ecosystem services. Glob. Chang. Biol. 2018, 24, 1382–1393. [Google Scholar] [CrossRef]
- Young, A.M.; Higuera, P.E.; Duffy, P.A.; Hu, F.S. Climatic thresholds shape northern high-latitude fire regimes and imply vulnerability to future climate change. Ecography 2017, 40, 606–617. [Google Scholar] [CrossRef]
- Lefebvre, G.; Redmond, L.; Germain, C.; Palazzi, E.; Terzago, S.; Willm, L.; Poulin, B. Predicting the vulnerability of seasonally-flooded wetlands to climate change across the Mediterranean Basin. Sci. Total Environ. 2019, 692, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Mouillot, F.; Rambal, S.; Joffre, R. Simulating climate change impacts on fire frequency and vegetation dynamics in a Mediterranean-type ecosystem. Glob. Chang. Biol. 2002, 8, 423–437. [Google Scholar] [CrossRef]
- Batllori, E.; Parisien, M.; Krawchuk, M.A.; Moritz, M.A. Climate change-induced shifts in fire for Mediterranean ecosystems. Glob. Ecol. Biogeogr. 2013, 22, 1118–1129. [Google Scholar] [CrossRef]
- Golluscio, R.A.; Bottaro, H.S.; Oesterheld, M. Controls of carrying capacity: Degradation, primary production, and forage quality effects in a Patagonian steppe. Rangel. Ecol. Manag. 2015, 68, 266–275. [Google Scholar] [CrossRef]
- IPCC. The Intergovernmental Panel on Climate Change Special Report; Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Kelly, A.E.; Goulden, M.L. Rapid shifts in plant distribution with recent climate change. Proc. Natl. Acad. Sci. USA 2008, 105, 11823–11826. [Google Scholar] [CrossRef]
- Black, E. The impact of climate change on daily precipitation statistics in Jordan and Israel. Atmos. Sci. Lett. 2009, 10, 192–200. [Google Scholar] [CrossRef]
- Turco, M.; Levin, N.; Tessler, N.; Saaroni, H. Recent changes and relations among drought, vegetation and wildfires in the Eastern Mediterranean: The case of Israel. Glob. Planet. Chang. 2017, 151, 28–35. [Google Scholar] [CrossRef]
- Golodets, C.; Sternberg, M.; Kigel, J.; Boeken, B.; Henkin, Z.; Seligman, N.G.; Ungar, E.D. From desert to Mediterranean rangelands: Will increasing drought and inter-annual rainfall variability affect herbaceous annual primary productivity? Clim. Chang. 2013, 119, 785–798. [Google Scholar] [CrossRef]
- Schaub, S.; Finger, R.; Leiber, F.; Probst, S.; Kreuzer, M.; Weigelt, A.; Buchmann, N.; Scherer-Lorenzen, M. Plant diversity effects on forage quality, yield and revenues of semi-natural grasslands. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Kafle, H.K.; Bruins, H.J. Climatic trends in Israel 1970–2002: Warmer and increasing aridity inland. Clim. Chang. 2009, 96, 63–77. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Adams, H.D.; Zeppel, M.J.; Anderegg, W.R.; Hartmann, H.; Landhäusser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Dovrat, G.; Meron, E.; Shachak, M.; Golodets, C.; Osem, Y. Plant size is related to biomass partitioning and stress resistance in water-limited annual plant communities. J. Arid Environ. 2019, 165, 1–9. [Google Scholar] [CrossRef]
- De Dato, G.; Pellizzaro, G.; Cesaraccio, C.; Sirca, C.; De Angelis, P.; Duce, P.; Spano, D.; Scarascia, M.G. Effects of warmer and drier climate conditions on plant composition and biomass production in a Mediterranean shrubland community. For. Ecol. 2008, 3, 511–526. [Google Scholar] [CrossRef]
- Köchy, M.; Mathaj, M.; Jeltsch, F.; Malkinson, D. Resilience of stocking capacity to changing climate in arid to Mediterranean landscapes. Reg. Environ. Chang. 2008, 8, 73–87. [Google Scholar] [CrossRef]
- McGill, B.J.; Etienne, R.S.; Gray, J.S.; Alonso, D.; Anderson, M.J.; Benecha, H.K.; Dornelas, M.; Enquist, B.J.; Green, J.L.; He, F.; et al. Species abundance distributions: Moving beyond single prediction theories to integration within an ecological framework. Ecol. Lett. 2007, 10, 995–1015. [Google Scholar] [CrossRef]
- Le Roux, P.C.; McGeoch, M.A. Rapid range expansion and community reorganization in response to warming. Glob. Chang. Biol. 2008, 14, 2950–2962. [Google Scholar] [CrossRef]
- Ellis, E.C.; Klein, G.K.; Siebert, S.; Lightman, D.; Ramankutty, N. Anthropogenic transformation of the biomes, 1700 to 2000. Glob. Ecol. Biogeogr. 2010, 19, 589–606. [Google Scholar] [CrossRef]
- Landau, S.Y.; Dvash, L.; Yehuda, Y.; Muklada, H.; Peleg, G.; Henkin, Z.; Voet, H.; Ungar, E.D. Impact of animal density on cattle nutrition in dry Mediterranean rangelands: A faecal NIRS-aided study. Animal 2018, 12, 265–274. [Google Scholar] [CrossRef]
- Ungar, E.D. Perspectives on the concept of rangeland carrying capacity, and their exploration by means of Noy-Meir’s two-function model. Agric. Syst. 2019, 173, 403–413. [Google Scholar] [CrossRef]
- Miranda, J.D.; Armas, C.; Padilla, F.M.; Pugnaire, F.I. Climatic change and rainfall patterns: Effects on semi-arid plant communities of the Iberian Southeast. J. Arid Environ. 2011, 75, 1302–1309. [Google Scholar] [CrossRef]
- Klein, J.A.; Harte, J.; Zhao, X. Experimental warming, not grazing, decreases rangeland quality on the Tibetan Plateau. Ecol. Appl. 2007, 17, 541–557. [Google Scholar] [CrossRef] [PubMed]
- Osoro, K.; Ferreira, L.; García, U.; Martínez, A.; Celaya, R. Forage intake, digestibility and performance of cattle, horses, sheep and goats grazing together on an improved heathland. Anim. Prod. Sci. 2017, 57, 102–109. [Google Scholar] [CrossRef]
- George, M.; Nader, G.; Dunbar, J. Balancing Beef Cow Nutrient Requirements and Seasonal Forage Quality on Annual Rangeland; Division of Agriculture and Natural Resources, University of California: Oakland, CA, USA, 2001. [Google Scholar]
- Henkin, Z.; Landau, S.; Ungar, E.; Perevolotsky, A.; Yehuda, Y.; Sternberg, M. Effect of timing and intensity of grazing on the herbage quality of a Mediterranean rangeland. J. Anim. Feed Sci. 2007, 16, 318–322. [Google Scholar] [CrossRef]
- Smart, A.J.; Derner, J.D.; Hendrickson, J.R.; Gillen, R.L.; Dunn, B.H.; Mousel, E.M.; Johnson, P.S.; Gates, R.N.; Sedivec, K.K.; Harmoney, K.R.; et al. Effects of grazing pressure on efficiency of grazing on North American Great Plains rangelands. Rangel. Ecol. Manag. 2010, 63, 397–406. [Google Scholar] [CrossRef]
- Hanley, M.E.; Lamont, B.B.; Fairbanks, M.M.; Rafferty, C.M. Plant structural traits and their role in anti-herbivore defense. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 157–178. [Google Scholar] [CrossRef]
- Dovrat, G.; Meron, E.; Shachak, M.; Golodets, C.; Osem, Y. Functional reorganization and productivity of a water-limited annual plant community. Plant Ecol. 2020, 221, 191–204. [Google Scholar] [CrossRef]
- Henkin, Z.; Ungar, E.D.; Dvash, L.; Perevolotsky, A.; Yehuda, Y.; Sternberg, M.; Voet, H.; Landau, S.Y. Effects of cattle grazing on herbage quality in a herbaceous Mediterranean rangeland. Grass Forage Sci. 2011, 66, 516–525. [Google Scholar] [CrossRef]
- Ryser, P. The importance of tissue density for growth and life span of leaves and roots: A comparison of five ecologically contrasting grasses. Funct. Ecol. 1996, 1, 717–723. [Google Scholar] [CrossRef]
- Diaz, S.; Lavorel, S.; McIntyre, S.U.E.; Falczuk, V.; Casanoves, F.; Milchunas, D.G.; Skarpe, C.; Rusch, G.; Sternberg, M.; Landsberg, J.; et al. Plant trait responses to grazing–a global synthesis. Glob. Chang. Biol. 2007, 13, 313–341. [Google Scholar] [CrossRef]
- Dovrat, G.; Meron, E.; Shachak, M.; Golodets, C.; Osem, Y. The relative contributions of functional diversity and functional identity to ecosystem function in water-limited environments. J. Veg. Sci. 2019, 30, 427–437. [Google Scholar] [CrossRef]
- Landau, S.Y.; Glasser, T.; Dvash, L. Monitoring nutrition in small ruminants with the aid of near infrared reflectance spectroscopy (NIRS) technology: A review. Small Rumin. Res. 2006, 61, 1–11. [Google Scholar] [CrossRef]
- Gutman, M.; Seligman, N.G. Grazing management of Mediterranean foothill range in the upper Jordan River valley. J. Range Manag. 1979, 32, 86–92. [Google Scholar] [CrossRef]
- Sternberg, M.; Gutman, M.; Perevolotsky, A.; Ungar, E.; Kigel, J. Vegetation response to grazing management in a Mediterranean herbaceous community: A functional group approach. J. Appl. Ecol. 2000, 37, 224–237. [Google Scholar] [CrossRef]
- Henkin, Z.; Ungar, E.D.; Perevolotsky, A.; Gutman, M.; Yehuda, Y.; Dolev, A.; Landau, S.Y.; Sternberg, M.; Seligman, N.G. Long-term trade-offs between herbage growth, animal production and supplementary feeding in heavily grazed Mediterranean grassland. Rangel. Ecol. Manag. 2015, 68, 332–340. [Google Scholar] [CrossRef]
- Tilley, J.M.; Terry, R.A. A two-stage technique for the in vitro digestion of forage crops. Grass Forage Sci. 1963, 18, 104–111. [Google Scholar] [CrossRef]
- Cunniff, P. Official Methods of Analysis of AOAC International, 16th ed.; AOAC International: Arlington, VA, USA, 1995; Volume 1. [Google Scholar]
- Goering, H.K.; Van Soest, P.J. Forage Fiber Analyses, Apparatus, Reagents, Procedures, and Some Applications. Agriculture Handbook; No. 379; ARS-USDA: Washington, DC, USA, 1970. [Google Scholar]
- Murphy, L. Likelihood: Methods for Maximum Likelihood Estimation, R Package. Version 1.4; 2012. Available online: http://www.sortie-nd.org/lme/lme_R_code_tutorials.html (accessed on 2 January 2020).
- Edwards, A.W. Likelihood, Expanded Edition; Johns Hopkins University Press: Baltimore, MD, USA, 1992. [Google Scholar]
- Fay, P.A.; Carlisle, J.D.; Knapp, A.K.; Blair, J.M.; Collins, S.L. Productivity responses to altered rainfall patterns in a C4 dominated grassland. Oecologia 2003, 137, 245–251. [Google Scholar] [CrossRef]
- de Aldana, B.R.V.; García Criado, B. Interannual variations of above-ground biomass and nutritional quality of Mediterranean grasslands in Western Spain over a 20-year period. Aust. J. Agric. Res. 2008, 59, 769–779. [Google Scholar] [CrossRef]
- Elgersma, A.; Søegaard, K. Changes in nutritive value and herbage yield during extended growth intervals in grass–legume mixtures: Effects of species, maturity at harvest, and relationships between productivity and components of feed quality. Grass Forage Sci. 2018, 73, 78–93. [Google Scholar] [CrossRef]
- Golodets, C.; Kigel, J.; Sternberg, M. Recovery of plant species composition and ecosystem function after cessation of grazing in a Mediterranean grassland. Plant Soil 2010, 329, 365–378. [Google Scholar] [CrossRef]
- Noy-Meir, I.; Gutman, M.; Kaplan, Y. Responses of Mediterranean grassland plants to grazing and protection. J. Ecol. 1989, 77, 290–310. [Google Scholar] [CrossRef]
- Perevolotsky, A.; Seligman, N. Role of grazing in Mediterranean rangeland ecosystems. Bioscience 1998, 48, 1007–1017. [Google Scholar] [CrossRef]
- Cherif, E.; Doblas-Miranda, E.; Lionelloet, P.; Borrego, C.; Giorgi, F.; Iglesias, A.; Jebari, S.; Mahmoudi, E.; Moriondo, M.; Pringault, O.; et al. Climate and Environmental Change in the Mediterranean Basin—Current Situation and Risks for the Future, First Mediterranean Assessment Report (MAR1); Chapter II Drivers of Change; UNEP/MAP: Marseille, France, 2021. [Google Scholar]
- Tielbörger, K.; Bilton, M.C.; Metz, J.; Kigel, J.; Holzapfel, C.; Lebrija-Trejos, E.; Konsens, I.; Parag, H.A.; Sternberg, M. Middle-Eastern plant communities tolerate 9 years of drought in a multi-site climate manipulation experiment. Nat. Commun. 2014, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dovrat, G.; Sheffer, E.; Landau, S.Y.; Deutch, T.; Gorelik, H.; Henkin, Z. Can Grazing Moderate Climatic Effects on Herbage Nutritional Quality? Agronomy 2021, 11, 700. https://doi.org/10.3390/agronomy11040700
Dovrat G, Sheffer E, Landau SY, Deutch T, Gorelik H, Henkin Z. Can Grazing Moderate Climatic Effects on Herbage Nutritional Quality? Agronomy. 2021; 11(4):700. https://doi.org/10.3390/agronomy11040700
Chicago/Turabian StyleDovrat, Guy, Efrat Sheffer, Serge Yan Landau, Tova Deutch, Haim Gorelik, and Zalmen Henkin. 2021. "Can Grazing Moderate Climatic Effects on Herbage Nutritional Quality?" Agronomy 11, no. 4: 700. https://doi.org/10.3390/agronomy11040700
APA StyleDovrat, G., Sheffer, E., Landau, S. Y., Deutch, T., Gorelik, H., & Henkin, Z. (2021). Can Grazing Moderate Climatic Effects on Herbage Nutritional Quality? Agronomy, 11(4), 700. https://doi.org/10.3390/agronomy11040700