
Assessing the Impact of Rice Cultivation and Off-Season Period on Dynamics of Soil Enzyme Activities and Bacterial Communities in Two Agro-Ecological Regions of Mozambique

 ,
,  , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area Description
2.2. Soil Sampling
2.3. Chemical and Enzyme Activities in Bulk Soil
2.4. Bacterial Community Analyses of Rice-Field Soils
2.4.1. DNA Extraction and Partial 16S rRNA Gene Sequencing
2.4.2. Bioinformatic and Diversity Analyses
2.5. Statistical Analyses
2.6. Deposition of Sequences
3. Results
3.1. Chemical Characteristics of Bulk Soil in Relation to Rice Cultivation and Fallow Periods
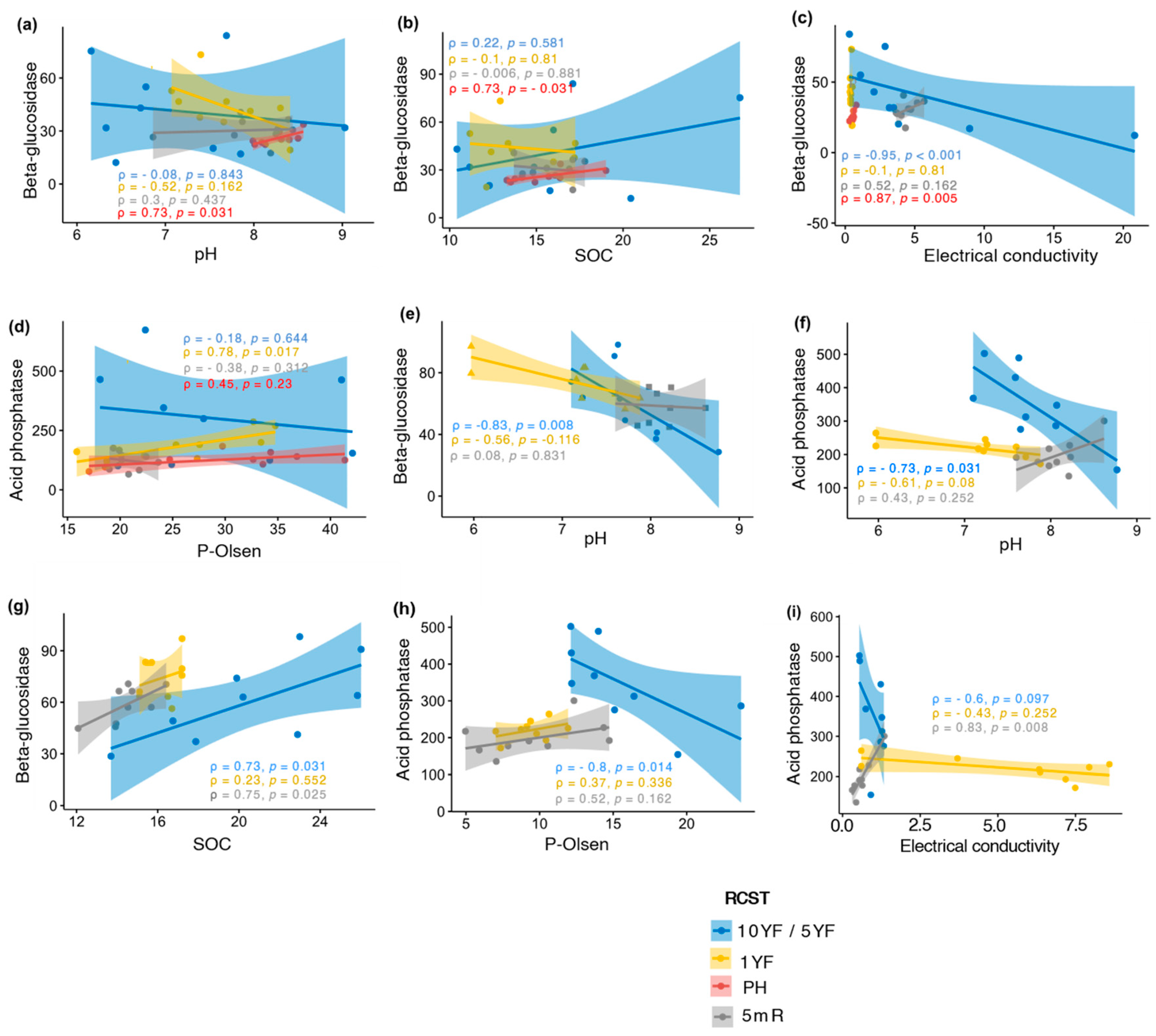
3.2. Enzyme Activities and Relationship with Sol Properties
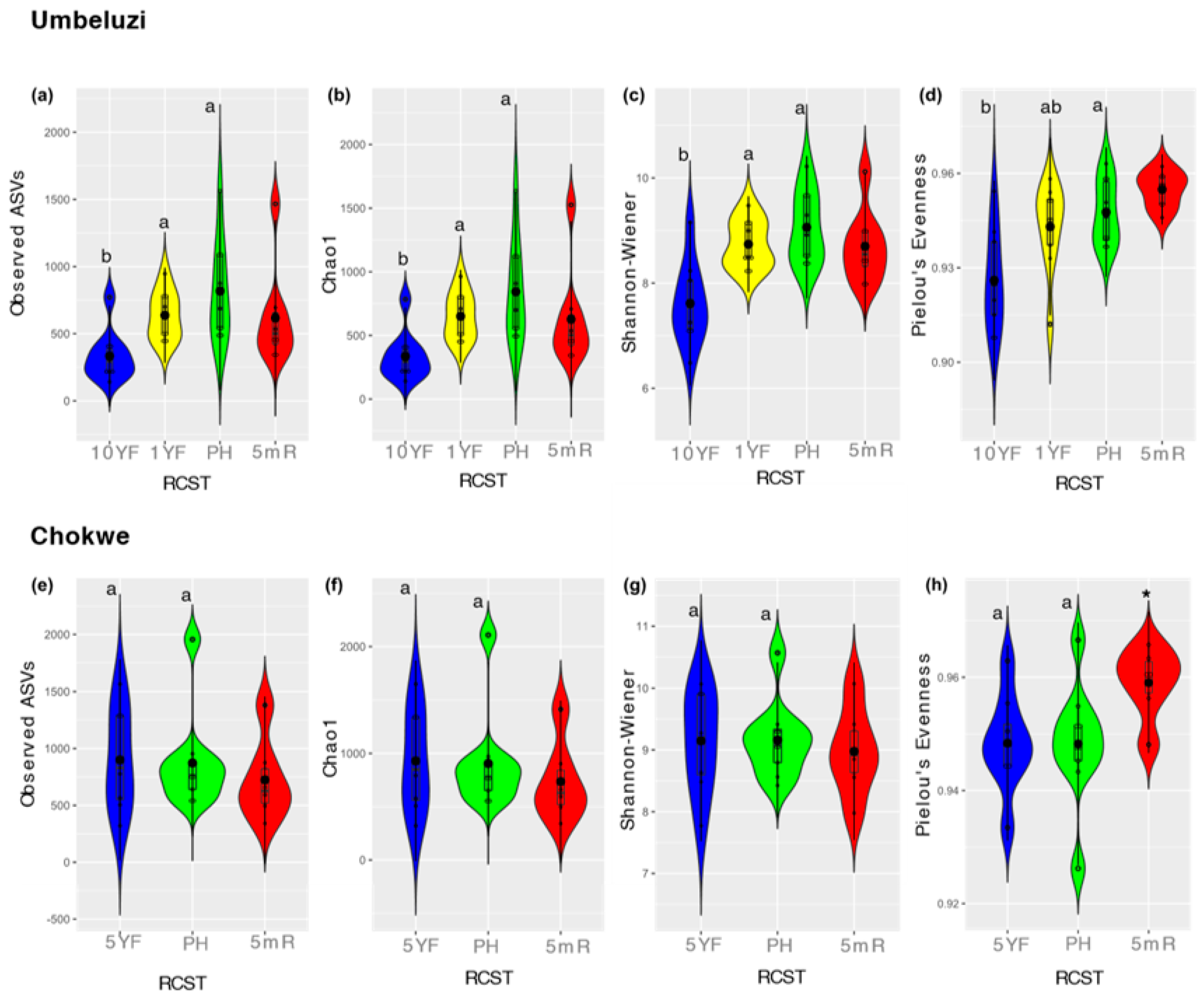
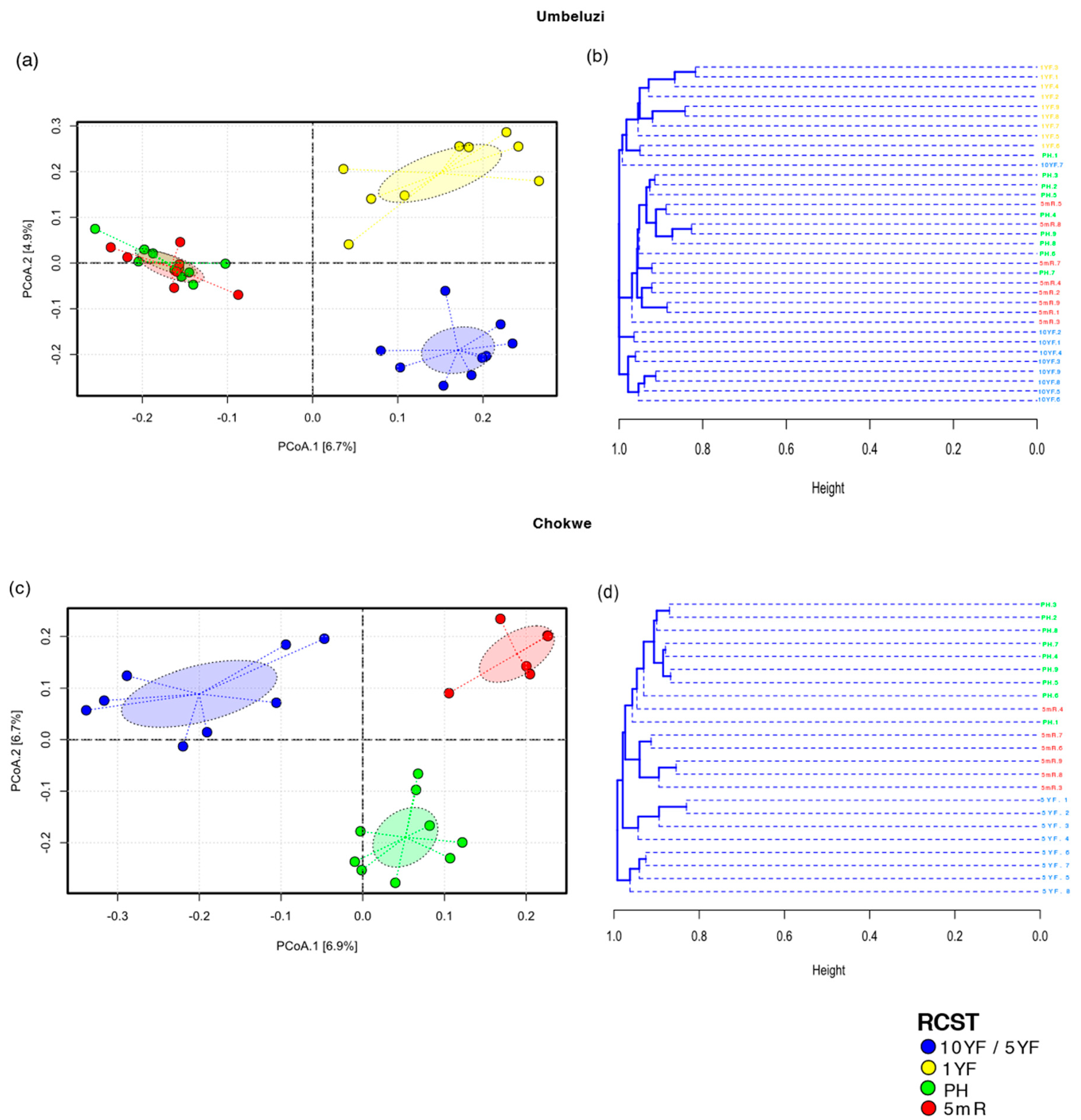
3.3. Bacterial Diversity and Community Structure in Fallow and Rice-Field Soils
3.3.1. Bacterial Diversity and Community Differentiation of ASVs
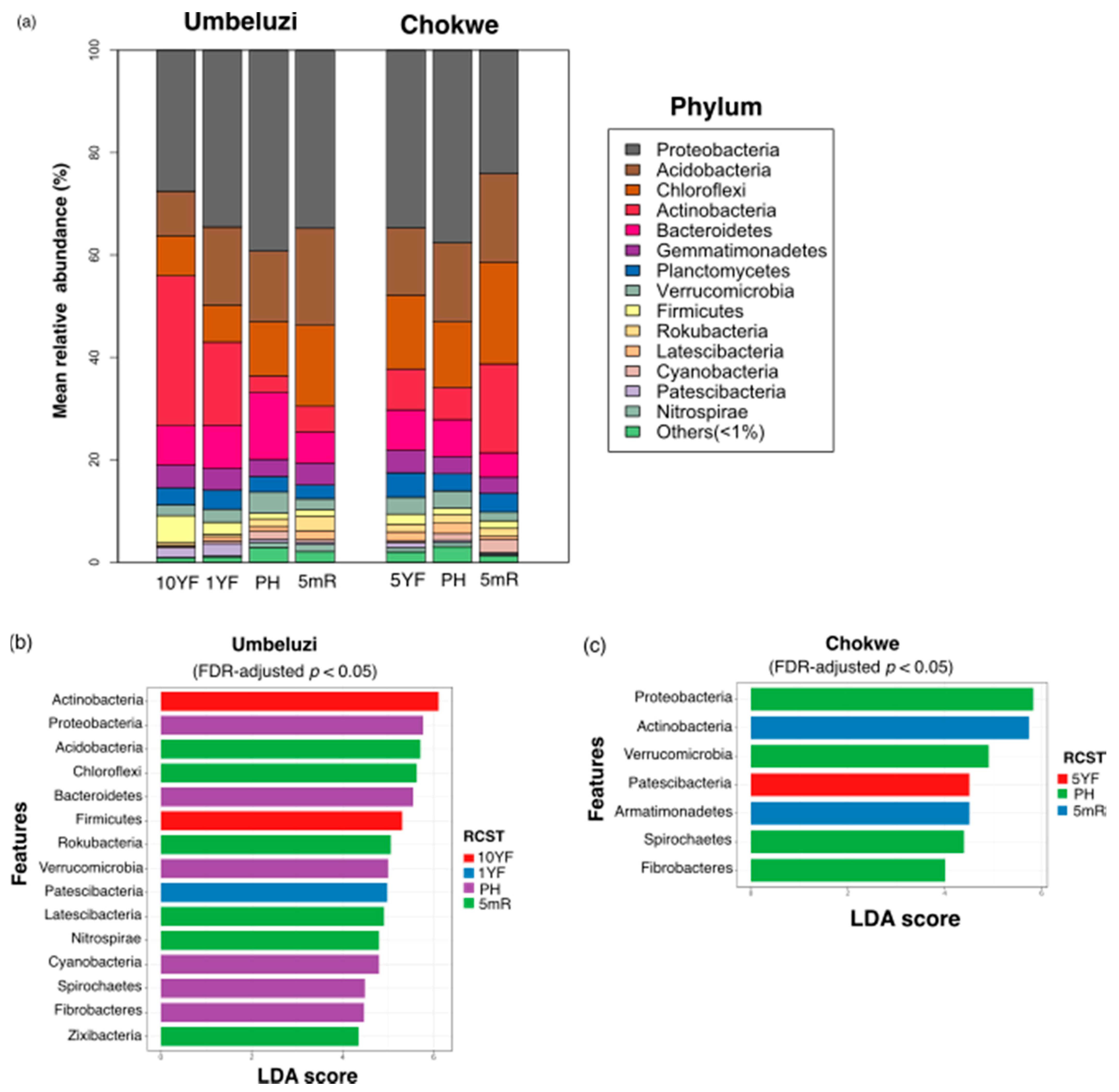
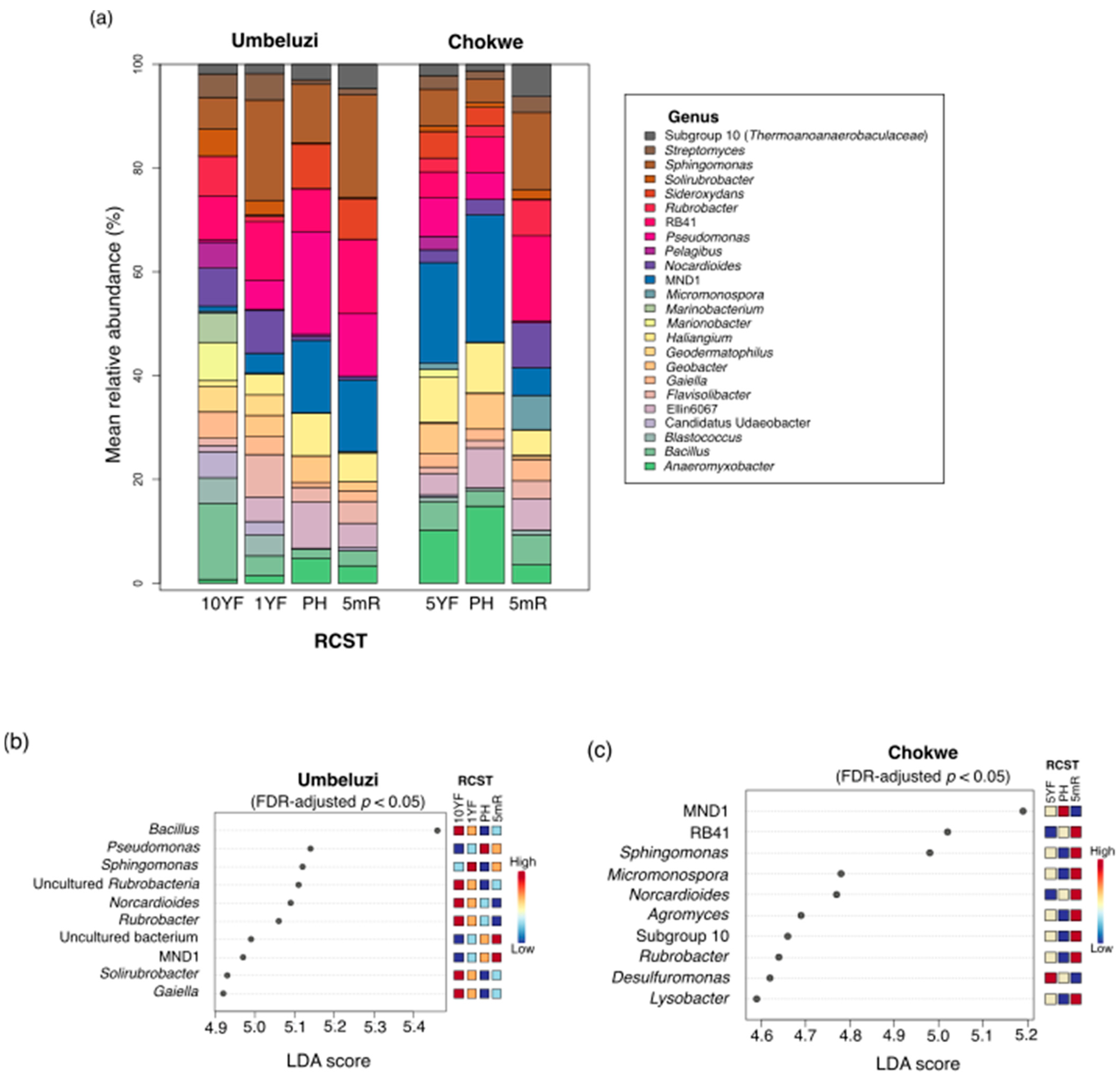
3.3.2. Taxonomic Diversity and Differentially Abundant Phylotypes
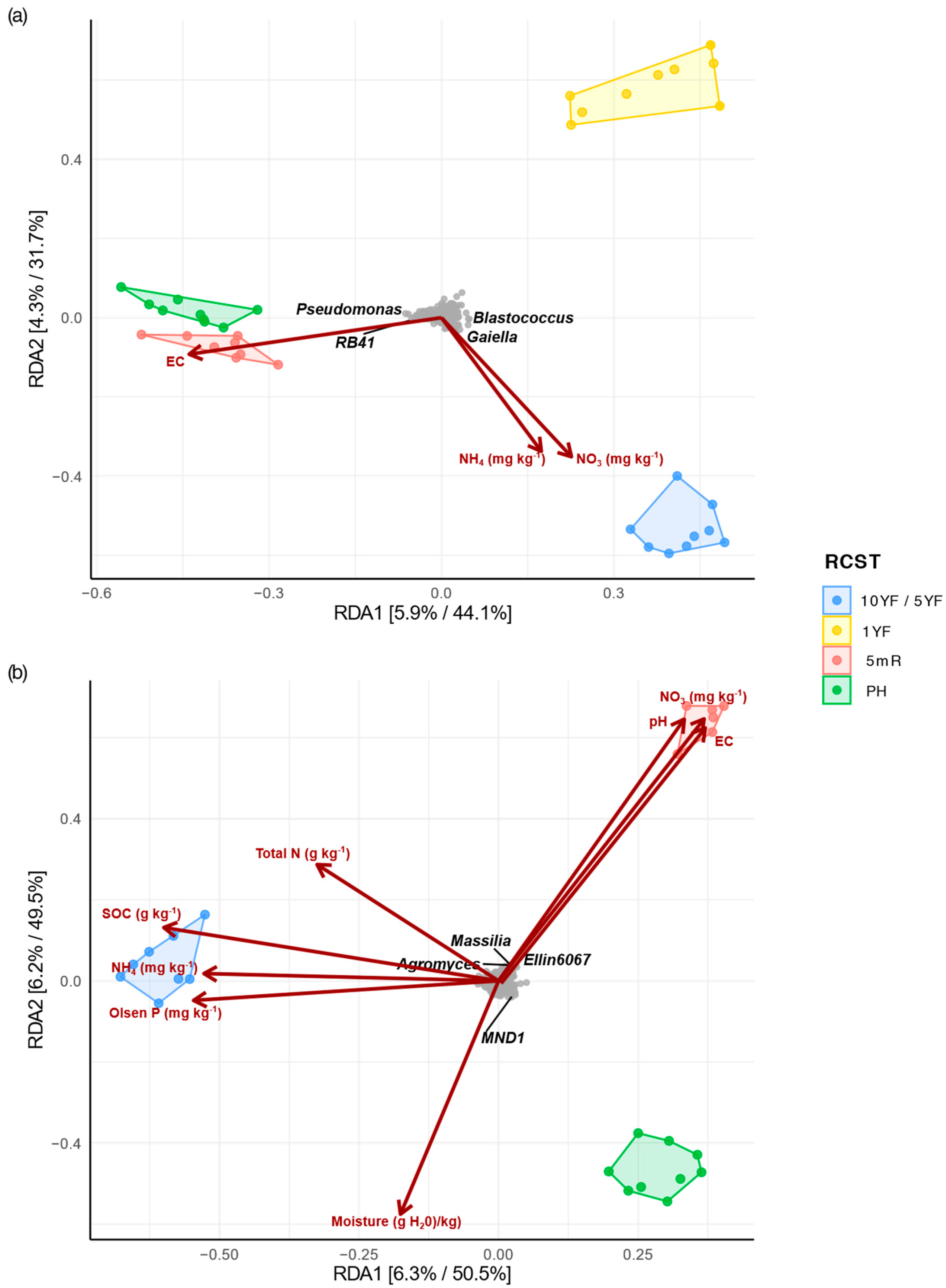
3.4. Relationship between Soil Properties and Bacterial Communities of RCST
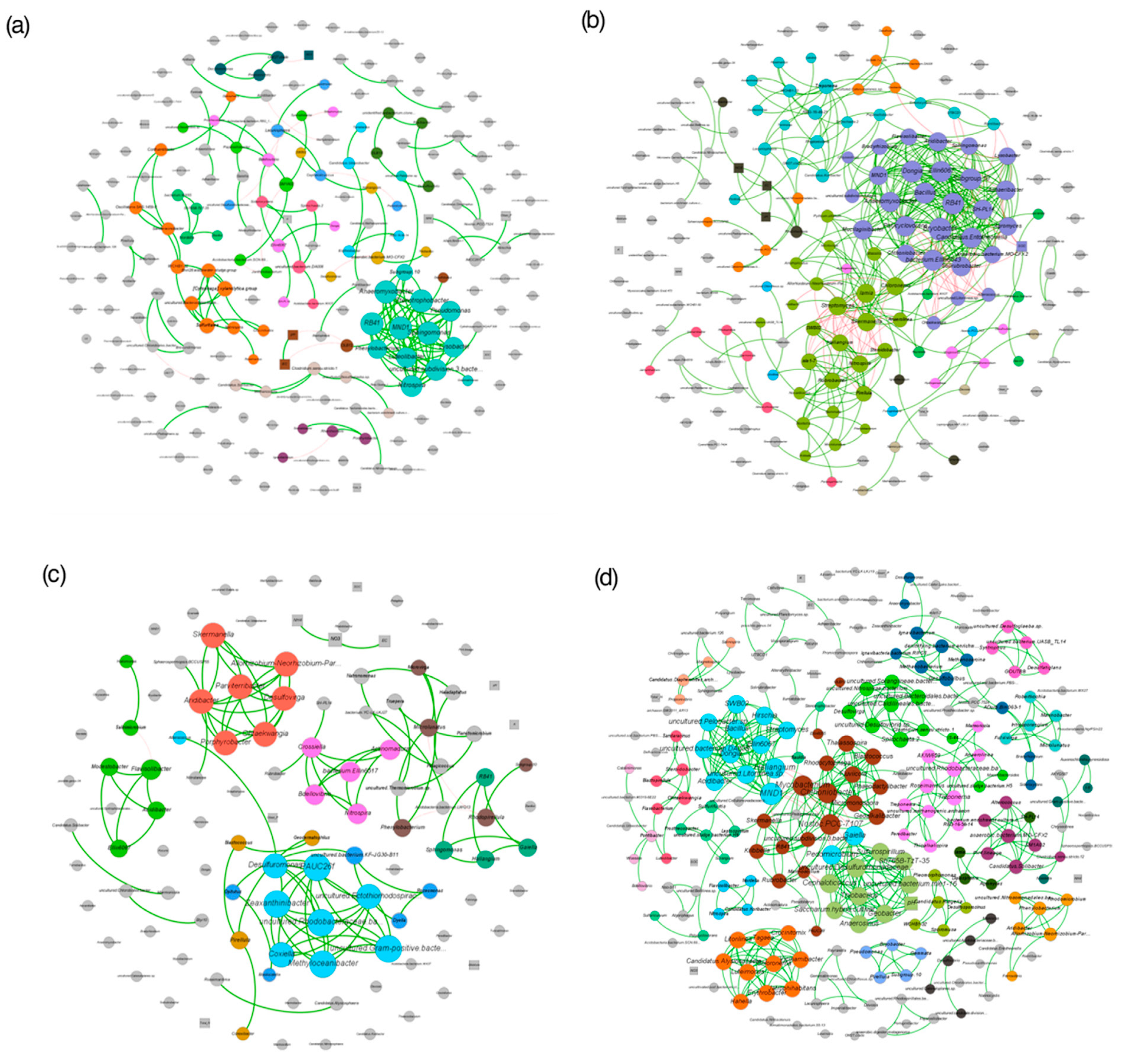
3.5. Co-Occurrence Networks of Microbes and Environmental Variables Interaction in Fallow and Rice-Cultivated Soils
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shabandeh, M. Worldwide production of grain in 2018/19, by type. Available online: https://www.statista.com/statistics/263977/world-grain-production-by-type/ (accessed on 25 October 2020).
- Calpe, C. Rice International Commodity Profile; Food and Agriculture Organiization: Rome, Italy; United Nations: New York, NY, USA, 2006; pp. 1–23. [Google Scholar]
- Gulati, A.; Narayanan, S. Rice trade libertization and poverty. Econ. Political Wkly. 2003, 38, 45–51. [Google Scholar]
- Research and Markets. Global Rice Market & Volume by Consumption, Production, Imports, Exports, Company Analysis & Forecast. 2021. Available online: https://www.researchandmarkets.com/reports/5237719/global-rice-market-and-volume-by-consumption?utm_source=BW&utm_medium=PressRelease&utm_code=jm9wt6&utm_campaign=1492948+-+Global+Rice+Market+Report+2021%3a+Market+%26+Volume+Analysis+by+Consumption%2c+Production%2c+Imports%2c+Exports%2c+Company+Analysis+%26+Forecast&utm_exec=chdo54prd (accessed on 10 February 2021).
- Van Nguyen, N.; Ferrero, A. Meeting the challenges of global rice production. Paddy Water Environ. 2006, 4, 1–9. [Google Scholar] [CrossRef]
- Fahad, S.; Adnan, M.; Noor, M.; Arif, M.; Alam, M.; Khan, I.A.; Ullah, H.; Wahid, F.; Mian, I.A.; Jamal, Y.; et al. Major constraints for global rice production. In Advances in Rice Research for Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Nahar, K., Biswas, J.K., Eds.; Elsevier Science: London, UK, 2018; pp. 1–22. ISBN 9780128143322. [Google Scholar]
- Prasad, R.; Shivay, Y.S.; Kumar, D. Current status, challenges, and opportunities in rice production. In Rice Production Worldwide; Chauhan, B., Jabran, K., Mahajan, G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–32. ISBN 9783319475165. [Google Scholar]
- deGraft-Johnson, M.; Suzuki, A.; Sakurai, T.; Otsuka, K. On the transferability of the Asian rice green revolution to rainfed areas in sub-Saharan Africa: An assessment of technology intervention in Northern Ghana. Agric. Econ. 2014, 45, 555–570. [Google Scholar] [CrossRef]
- Nin-Pratt, A.; McBride, L. Agricultural intensification in Ghana: Evaluating the optimist’s case for a Green Revolution. Food Policy 2014, 48, 153–167. [Google Scholar] [CrossRef]
- Amanor, K.S.; Chichava, S. South-South Cooperation, Agribusiness, and African Agricultural Development: Brazil and China in Ghana and Mozambique. World Dev. 2016, 81, 13–23. [Google Scholar] [CrossRef]
- Van Oort, P.A.J.; Saito, K.; Tanaka, A.; Amovin-Assagba, E.; Van Bussel, L.G.J.; van Wart, J.; de Groot, H.; van Ittersum, M.K.; Cassman, K.G.; Wopereis, M.C.S. Assessment of rice self-sufficiency in 2025 in eight African countries. Glob. Food Sec. 2014, 5, 39–49. [Google Scholar] [CrossRef]
- Haefele, S.M.; Nelson, A.; Hijmans, R.J. Soil quality and constraints in global rice production. Geoderma 2014, 235–236, 250–259. [Google Scholar] [CrossRef]
- Burns, R.G.; Deforest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil Biology & Biochemistry Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Yang, L.; Li, T.; Li, F.; Lemcoff, J.H.; Cohen, S. Fertilization regulates soil enzymatic activity and fertility dynamics in a cucumber field. Sci. Hortic. 2008, 116, 21–26. [Google Scholar] [CrossRef]
- Adeleke, R.; Nwangburuka, C.; Oboirien, B. Origins, roles and fate of organic acids in soils: A review. S. Afr. J. Bot. 2017, 108, 393–406. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Hämmerle, I.; Fuchslueger, L.; Hofhansl, F.; Knoltsch, A.; Schnecker, J.; Takriti, M.; Watzka, M.; Wild, B.; et al. Adjustment of microbial nitrogen use efficiency to carbon:Nitrogen imbalances regulates soil nitrogen cycling. Nat. Commun. 2014, 5, 1–7. [Google Scholar] [CrossRef]
- Ezeokoli, O.T.; Bezuidenhout, C.C.; Maboeta, M.S.; Khasa, D.P.; Adeleke, R.A. Structural and functional differentiation of bacterial communities in post-coal mining reclamation soils of South Africa: Bioindicators of soil ecosystem restoration. Sci. Rep. 2020, 10, 1759. [Google Scholar] [CrossRef]
- Ezeokoli, O.T.; Oladipo, O.G.; Bezuidenhout, C.C.; Adeleke, R.A.; Maboeta, M.S. Assessing the ecosystem support function of South African coal mining soil environments using earthworm (Eisenia andrei) bioassays. Appl. Soil Ecol. 2021, 157, 103771. [Google Scholar] [CrossRef]
- Ezeokoli, O.T.; Mashigo, S.K.; Maboeta, M.S.; Bezuidenhout, C.C.; Khasa, D.P.; Adeleke, R.A. Arbuscular mycorrhizal fungal community differentiation along a post-coal mining reclamation chronosequence in South Africa: A potential indicator of ecosystem recovery. Appl. Soil Ecol. 2020, 147, 103429. [Google Scholar] [CrossRef]
- Ashraf, M.N.; Hu, C.; Wu, L.; Duan, Y.; Zhang, W.; Aziz, T.; Cai, A.; Abrar, M.M.; Xu, M. Soil and microbial biomass stoichiometry regulate soil organic carbon and nitrogen mineralization in rice-wheat rotation subjected to long-term fertilization. J. Soils Sediments 2020, 20, 3103–3113. [Google Scholar] [CrossRef]
- Aspiras, R.B.; Allen, O.N.; Harris, R.F.; Chesters, G. The role of microorganisms in the stabilization of soil aggregates. Soil Biol. Biochem. 1971, 3, 347–353. [Google Scholar] [CrossRef]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Lareen, A.; Burton, F.; Schäfer, P. Plant root-microbe communication in shaping root microbiomes. Plant Mol. Biol. 2016, 90, 575–587. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef]
- Tan, S.; Gu, Y.; Yang, C.; Dong, Y.; Mei, X.; Shen, Q.; Xu, Y. Bacillus amyloliquefaciens T-5 may prevent Ralstonia solanacearum infection through competitive exclusion. Biol. Fertil. Soils 2015, 52, 341–351. [Google Scholar] [CrossRef]
- Van Der Ent, S.; Van Hulten, M.; Pozo, M.J.; Czechowski, T.; Udvardi, M.K.; Pieterse, C.M.J.; Ton, J. Priming of plant innate immunity by rhizobacteria and β-aminobutyric acid: Differences and similarities in regulation. New Phytol. 2009, 183, 419–431. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, D.; Yang, Y.; Pan, Y.; Zhao, D.; Zhu, J.; Zhang, L.; Yang, Z. Dissecting the effect of continuous cropping of potato on soil bacterial communities as revealed by high-throughput sequencing. PLoS ONE 2020, 15, e0233356. [Google Scholar] [CrossRef]
- Sun, J.; Zou, L.; Li, W.; Wang, Y.; Xia, Q.; Peng, M. Soil microbial and chemical properties influenced by continuous cropping of banana. Sci. Agric. 2018, 75, 420–425. [Google Scholar] [CrossRef]
- Ng, J.P.; Hollister, E.B.; González-Chávez, M.; del Carmen, A.; Hons, F.M.; Zuberer, D.A.; Aitkenhead-Peterson, J.A.; Loeppert, R.; Gentry, T.J.. Impacts of Cropping Systems and Long-Term Tillage on Soil Microbial Population Levels and Community Composition in Dryland Agricultural Setting. ISRN Ecol. 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Wang, W.; Luo, X.; Chen, Y.; Ye, X.; Wang, H.; Cao, Z.; Ran, W.; Cui, Z. Succession of composition and function of soil bacterial communities during key rice growth stages. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Luan, H.; Gao, W.; Huang, S.; Tang, J.; Li, M.; Zhang, H.; Chen, X.; Masiliūnas, D. Substitution of manure for chemical fertilizer affects soil microbial community diversity, structure and function in greenhouse vegetable production systems. PLoS ONE 2020, 15, e0214041. [Google Scholar] [CrossRef]
- Dumontet, S.; Cavoski, I.; Ricciuti, P.; Mondelli, D.; Jarrar, M.; Pasquale, V.; Crecchio, C. Metabolic and genetic patterns of soil microbial communities in response to different amendments under organic farming system. Geoderma 2017, 296, 79–85. [Google Scholar] [CrossRef]
- Ashworth, A.J.; DeBruyn, J.M.; Allen, F.L.; Radosevich, M.; Owens, P.R. Microbial community structure is affected by cropping sequences and poultry litter under long-term no-tillage. Soil Biol. Biochem. 2017, 114, 210–219. [Google Scholar] [CrossRef]
- Choudhary, M.; Sharma, P.C.; Jat, H.S.; Dash, A.; Rajashekar, B.; McDonald, A.J.; Jat, M.L. Soil bacterial diversity under conservation agriculture-based cereal systems in Indo-Gangetic Plains. 3 Biotech 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Habig, J.; Swanepoel, C. Effects of conservation agriculture and fertilization on soil microbial diversity and activity. Environments 2015, 2, 358–384. [Google Scholar] [CrossRef]
- Li, Y.; Chang, S.X.; Tian, L.; Zhang, Q. Conservation agriculture practices increase soil microbial biomass carbon and nitrogen in agricultural soils: A global meta-analysis. Soil Biol. Biochem. 2018, 121, 50–58. [Google Scholar] [CrossRef]
- Lopes, L.D.; Fernandes, M.F. Changes in microbial community structure and physiological profile in a kaolinitic tropical soil under different conservation agricultural practices. Appl. Soil Ecol. 2020, 152, 103545. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef] [PubMed]
- Liesack, W.; Schnell, S.; Revsbech, N.P. Microbiology of flooded rice paddies. FEMS Microbiol. Rev. 2000, 24, 625–645. [Google Scholar] [CrossRef]
- Sun, W.; Xiao, E.; Pu, Z.; Krumins, V.; Dong, Y.; Li, B.; Hu, M. Paddy soil microbial communities driven by environment- and microbe-microbe interactions: A case study of elevation-resolved microbial communities in a rice terrace. Sci. Total Environ. 2018, 612, 884–893. [Google Scholar] [CrossRef]
- MAF (Ministry of Agriculture and Fisheries of Mozambique). ROAGRI Formulation Process. Agro-ecological zones and production systems. In Investment Program with Agricultural Extension; MAF: Maputo, Mozambique, 1996. [Google Scholar]
- INIA (Instituto Nacional de Investigação; Agronómica). Carta dos solos de Moçambique-Maputo; Ministério da Agricultura: Maputo, Moçambique, 1995. [Google Scholar]
- FAEF (Faculty of Agronomy and Forestry Engineering). Programa Competir: Região Agrícola do Chókwè Diagnóstico da Fileira Agrícola; FAEF: Maputo, Moçambique, 2001. [Google Scholar]
- Olsen, S.R.; Cole, C.V.; Sterling, R.; Watanabe, F.F.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; USDA Circular 939; US Government Printing Office: Washington, DC, USA, 1954. [Google Scholar]
- Claessen, M.E.C. Manual for Methods of Soil Analysis, 2nd ed.; Embrapa Solos: Rio de Janeiro, Brazil, 1997. [Google Scholar]
- Egnér, H.; Riehm, H.; Domingo, W.R. Untersuchungen über die chemische Bodenanalyse als Grundlage für die Beurteilung des Nährstoffzustandes der Böden. II. Chem. Extraktionsmethoden zur Phosphor- und Kaliumbestimmung K. Lantbr. Ann. 1960, 26, 199–215. [Google Scholar]
- Tabatabai, M.A. Soil Enzymes. In Methods of Soil Analysis, Part 2: Microbiological and Biochemical Properties; Weaver, R.W., Angle, J.S., Bottomley, P.S., Eds.; Book Series 5; Soil Science Society of America: Madison, WI, USA, 1994; pp. 775–833. [Google Scholar]
- Kandeler, E. Urease activity by colorimetric technique. In Methods in Soil Biology; Schinner, F., Kandeler, E., Öhlinger, R., Margesin, R., Eds.; Springer: Berlin, Germany, 1995; pp. 171–174. [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2 contributed to the design of analytical methods HHS Public Access Author manuscript. Nat Biotechnol 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O.; Glo, F.O.; Yarza, P.; et al. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H.; et al. vegan: Community Ecology Package. Available online: https://cran.r-project.org/package=vegan (accessed on 2 February 2020).
- Roberts, D.W. Labdsv: Ordination and Multivariate Analysis for Ecology. R Package Version 2.0-1. 2010. Available online: https://cran.rproject.org/web/packages/labdsv/index.html#:~:text=labdsv%3A%20Ordination%20and%20Multivariate%20Analysis,well%20as%20several%20novel%20analyses (accessed on 10 February 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; Version 4.0.2; R Core Team: Vienna, Austria, 2017. [Google Scholar]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Galili, T. dendextend: An R package for visualizing, adjusting and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Tackmann, J.; Matias Rodrigues, J.F.; von Mering, C. Rapid Inference of Direct Interactions in Large-Scale Ecological Networks from Heterogeneous Microbial Sequencing Data. Cell Syst. 2019, 9, 286–296.e8. [Google Scholar] [CrossRef]
- Aliferis, C.F.; Statnikov, A.; Tsamardinos, I.; Mani, S.; Koutsoukos, X.D. Local causal and markov blanket induction for causal discovery and feature selection for classification part I: Algorithms and empirical evaluation. J. Mach. Learn. Res. 2010, 11, 171–234. [Google Scholar]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterJournal Complex Syst. 2006, 1965, 1–9. [Google Scholar]
- Blondel, V.D.; Guillaume, J.L.; Lambiotte, R.; Lefebvre, E. Fast unfolding of communities in large networks. J. Stat. Mech. Theory Exp. 2008, 2008, P10008. [Google Scholar] [CrossRef]
- Dornbush, M.E.; von Haden, A.C. Intensified Agroecosystems and Their Effects on Soil Biodiversity and Soil Functions. In Soil Health and Intensification of Agroecosystems; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 173–193. ISBN 9780128054017. [Google Scholar]
- Kepler, R.M.; Epp Schmidt, D.J.; Yarwood, S.A.; Cavigelli, M.A.; Reddy, K.N.; Duke, S.O.; Bradley, C.A.; Williams, M.M.; Maula, J.E. Soil microbial communities in diverse agroecosystems exposed to the herbicide glyphosate. Appl. Environ. Microbiol. 2020, 86, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.; Wu, Q.; Zhu, J. Nitrogen and phosphorus losses from paddy fields and the yield of rice with different water and nitrogen management practices. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Spiertz, J.H.J. Nitrogen, sustainable agriculture and food security. A review. Agron. Sustain. Dev. 2010, 30, 43–55. [Google Scholar] [CrossRef]
- Ishii, S.; Ikeda, S.; Minamisawa, K.; Senoo, K. Nitrogen cycling in rice paddy environments: Past achievements and future challenges. Microbes Environ. 2011, 26, 282–292. [Google Scholar] [CrossRef]
- Li, Y.; Kronzucker, H.J.; Shi, W. Microprofiling of nitrogen patches in paddy soil: Analysis of spatiotemporal nutrient heterogeneity at the microscale. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Ahmed, W.; Qaswar, M.; Jing, H.; Wenjun, D.; Geng, S.; Kailou, L.; Ying, M.; Ao, T.; Mei, S.; Chao, L.; et al. Tillage practices improve rice yield and soil phosphorus fractions in two typical paddy soils. J. Soils Sediments 2020, 20, 850–861. [Google Scholar] [CrossRef]
- Rahman, S. Impact of soil fertility on rice productivity and efficiency: A case study from Bangladesh. In Soil Fertility; Lucero, D.P., Boggs, J., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2009; pp. 35–50. ISBN 9781607414667. [Google Scholar]
- Brady, N.C. Soil factors that influence rice production. In Proceedings of Symposium on Paddy Soils; Springer: Berlin/Heidelberg, Germany, 1981; pp. 1–19. [Google Scholar]
- Nguyen, T.T.; Sasaki, Y.; Kakuda, K.I.; Fujii, H. Comparison of paddy soil fertility under conventional rice straw application versus cow dung compost application in mixed crop–livestock systems in a cold temperate region of Japan. Soil Sci. Plant Nutr. 2020, 66, 106–115. [Google Scholar] [CrossRef]
- Li, Y.; Li, J.; Gao, L.; Tian, Y. Irrigation has more influence than fertilization on leaching water quality and the potential environmental risk in excessively fertilized vegetable soils. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Shakoor, M.B.; Riaz, M.; Niazi, N.K.; Ali, S.; Rizwan, M.; Arif, M.S.; Arif, M. Recent advances in arsenic accumulation in rice. In Advances in Rice Research for Abiotic Stress Tolerance; Elsevier: Amsterdam, The Netherlands, 2018; pp. 385–398. ISBN 9780128143322. [Google Scholar]
- Annisa, W.; Sosiawan, H. Dynamics of nitrogen nutrients in lowland soils with some irrigation conditions to increase rice crop productivity. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Bandung, Indonesia, 5–7 August 2019; Volume 393, p. 012058. [Google Scholar]
- Biswas, J.C.; Naher, U.A. Soil nutrient stress and rice production in Bangladesh. In Advances in Rice Research for Abiotic Stress Tolerance; Elsevier: Amsterdam, The Netherlands, 2018; pp. 431–445. ISBN 9780128143322. [Google Scholar]
- Ilagan, L.A.; Tablizo, R.P.; Barba, R.B.; Marquez, N.A. Soil Fertility Evaluation for Rice Production in Catanduanes Province, Philippines. Int. J. Sci. Technol. Res. 2014, 3, 81–87. [Google Scholar]
- Zhang, Q.C.; Wang, G.H. Studies on nutrient uptake of rice and characteristics of soil microorganisms in a long-term fertilization experiments for irrigated rice. J. Zhejiang Univ. Sci. 2005, 6 B, 147–154. [Google Scholar] [CrossRef]
- Melman, D.A.; Kelly, C.; Schneekloth, J.; Calderón, F.; Fonte, S.J. Tillage and residue management drive rapid changes in soil macrofauna communities and soil properties in a semiarid cropping system of Eastern Colorado. Appl. Soil Ecol. 2019, 143, 98–106. [Google Scholar] [CrossRef]
- Williams, H.; Colombi, T.; Keller, T. The influence of soil management on soil health: An on-farm study in southern Sweden. Geoderma 2020, 360, 114010. [Google Scholar] [CrossRef]
- Hubanks, H.L.; Deenik, J.L.; Crow, S.E. Getting the Dirt on Soil Health and Management. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Li, W.; Liu, M.; Wu, M.; Jiang, C.; Kuzyakov, Y. Bacterial community succession in paddy soil depending on rice fertilization. Appl. Soil Ecol. 2019, 144, 92–97. [Google Scholar] [CrossRef]
- Bolan, N.S.; Hedley, M.J.; White, R.E. Processes of soil acidification during nitrogen cycling with emphasis on legume based pastures. Plant Soil 1991, 134, 53–63. [Google Scholar] [CrossRef]
- Yu, T.R. Characteristics of soil acidity of paddy soils in relation to rice growth. In Plant-Soil Interactions at Low pH; Springer: Dordrecht, The Netherlands, 1991; pp. 107–112. [Google Scholar]
- Wang, Y.; Tang, C.; Wu, J.; Liu, X.; Xu, J. Impact of organic matter addition on pH change of paddy soils. J. Soils Sediments 2013, 13, 12–23. [Google Scholar] [CrossRef]
- Peng, S.Z.; Yang, S.H.; Xu, J.Z.; Luo, Y.F.; Hou, H.J. Nitrogen and phosphorus leaching losses from paddy fields with different water and nitrogen managements. Paddy Water Environ. 2011, 9, 333–342. [Google Scholar] [CrossRef]
- Patrick, W.H.; Mikkelsen, D.S.; Wells, B.R. Plant nutrient behavior in flooded soil. In Fertilizer Technology and Use; Soil Science Society of America: Madison, WI, USA, 2015; pp. 197–228. [Google Scholar]
- Nishikawa, T.; Li, K.; Inamura, T.; Nishikawa, T.; Li, K.; Nishikawa, T. Nitrogen uptake by the rice plant and changes in the soil chemical properties in the paddy rice field during yearly application of anaerobically-digested manure for seven years. Plant Prod. Sci. 2014, 17, 237–244. [Google Scholar] [CrossRef]
- Ahmad, N.; Abid, M.; Hussain, K.; Akram, M.; Yousaf, M. Evaluation of Nutrient Status in the Rice Growing Areas of the Punjab. Asian J. Plant Sci. 2003, 2, 449–453. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef]
- Wei, X.; Zhu, Z.; Wei, L.; Wu, J.; Ge, T. Rhizosphere Biogeochemical cycles of key elements in the paddy-rice rhizosphere: Microbial mechanisms and coupling processes. Rhizosphere 2019, 10, 100145. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Lewu, F.B.; Mulidzi, R.; Ncube, B. The biological activities of β-glucosidase, phosphatase and urease as soil quality indicators: A review. J. Soil Sci. Plant Nutr. 2017, 17, 794–807. [Google Scholar] [CrossRef]
- Stenberg, B. Monitoring soil quality of arable Land: Microbiological indicators. Acta Agric. Scand. Sect. B-Soil Plant Sci. 1999, 49, 1–24. [Google Scholar] [CrossRef]
- Kunito, T.; Shiroma, T.; Moro, H.; Sumi, H. Annual variation in soil enzyme activity in a paddy field: Soil temperature and nutrient availability are important for controlling enzyme activities. Appl. Environ. Soil Sci. 2018, 2018. [Google Scholar] [CrossRef]
- Dinesh, R.; Ghoshal Chaudhuri, S.; Sheeja, T.E. Soil biochemical and microbial indices in wet tropical forests: Effects of deforestation and cultivation. J. Plant Nutr. Soil Sci. 2004, 167, 24–32. [Google Scholar] [CrossRef]
- Raiesi, F.; Beheshti, A. Soil specific enzyme activity shows more clearly soil responses to paddy rice cultivation than absolute enzyme activity in primary forests of northwest Iran. Appl. Soil Ecol. 2014, 75, 63–70. [Google Scholar] [CrossRef]
- An, S.; Zheng, F.; Zhang, F.; Van Pelt, S.; Hamer, U.; Makeschin, F. Soil quality degradation processes along a deforestation chronosequence in the Ziwuling area, China. Catena 2008, 75, 248–256. [Google Scholar] [CrossRef]
- Gu, C.; Zhang, S.; Han, P.; Hu, X.; Xie, L.; Li, Y.; Brooks, M.; Liao, X.; Qin, L. Soil enzyme activity in soils subjected to flooding and the effect on nitrogen and phosphorus uptake by oilseed rape. Front. Plant Sci. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Dick, R.P.; Breakwell, D.P.; Turco, R.F. Soil enzyme activities and biodiversity measurements as integrative microbiological indicators. In Methods of Assessing Soil Quality; Doran, J.W., Jones, A.J., Eds.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 247–271. [Google Scholar]
- Das, S.K.; Varma, A. Role of Enzymes in Maintaining Soil Health. In Soil Enzymology; Shukla, G., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 22, pp. 119–148. [Google Scholar]
- Tabatabai, M.A.; Dick, W.A. Enzymes in soil. In Enzymes in the Environment: Activity, Ecology and Applications; Burns, R.G., Dick, R.P., Eds.; Marcel Dekker: New York, NY, USA, 2002; pp. 567–596. [Google Scholar]
- Dick, R.P. Soil Enzyme Activities as Indicators of Soil Quality. In Defining Soil Quality for a Sustainable Environment; John Wiley & Sons: Hoboken, NJ, USA, 1994; pp. 107–124. [Google Scholar]
- Dick, R.P.; Kandeler, E. Enzymes in Soils. In Encyclopedia of Soils in the Environment; Elsevier Inc.: Amsterdam, The Netherlands, 2004; Volume 4, pp. 448–456. ISBN 9780080547954. [Google Scholar]
- Ezeokoli, O.T.; Mashigo, S.K.; Paterson, D.G.; Bezuidenhout, C.C.; Adeleke, R.A. Microbial community structure and relationship with physicochemical properties of soil stockpiles in selected South African opencast coal mines. Soil Sci. Plant Nutr. 2019, 65, 332–341. [Google Scholar] [CrossRef]
- Shi, A.; Chakrawal, A.; Manzoni, S.; Fischer, B.M.C.; Nunan, N.; Herrmann, A.M. Substrate spatial heterogeneity reduces soil microbial activity. Soil Biol. Biochem. 2021, 152, 108068. [Google Scholar] [CrossRef]
- Ye, H.; Lu, C.; Lin, Q. Investigation of the spatial heterogeneity of soil microbial biomass carbon and nitrogen under long-term fertilizations in fluvo-aquic soil. PLoS ONE 2019, 14, e0209635. [Google Scholar] [CrossRef]
- Pittol, M.; Scully, E.; Miller, D.; Durso, L.; Mariana Fiuza, L.; Valiati, V.H. Bacterial community of the rice floodwater using cultivation-independent approaches. Int. J. Microbiol. 2018, 2018. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, Q.; Li, Z.; Cheng, W.; Sun, J.; Guo, Z.; Li, Y.; Zhou, J.; Meng, D.; Li, H.; et al. Environmental factors shaping the diversity of bacterial communities that promote rice production. BMC Microbiol. 2018, 18, 51. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Takemura, M.; Miki, T.; Nonaka, M.; Harada, N. Differences in soil bacterial community compositions in paddy fields under organic and conventional farming conditions. Microbes Environ. 2019, 34, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Tian, L.; Chang, C.; Li, X.; Tian, C. Cultivated rice rhizomicrobiome is more sensitive to environmental shifts than that of wild rice in natural environments. Appl. Soil Ecol. 2019, 140, 68–77. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, J.; Wang, C.; Wang, Q. The 19-years inorganic fertilization increased bacterial diversity and altered bacterial community composition and potential functions in a paddy soil. Appl. Soil Ecol. 2019, 144, 60–67. [Google Scholar] [CrossRef]
- Kumar, U.; Kumar Nayak, A.; Shahid, M.; Gupta, V.V.S.R.; Panneerselvam, P.; Mohanty, S.; Kaviraj, M.; Kumar, A.; Chatterjee, D.; Lal, B.; et al. Continuous application of inorganic and organic fertilizers over 47 years in paddy soil alters the bacterial community structure and its influence on rice production. Agric. Ecosyst. Environ. 2018, 262, 65–75. [Google Scholar] [CrossRef]
- Jiao, S.; Xu, Y.; Zhang, J.; Hao, X.; Lu, Y. Core Microbiota in Agricultural Soils and Their Potential Associations with Nutrient Cycling. mSystems 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, Y.; Zhao, L.; Zhang, W.; Liu, L. Change of soil microbial community under long-term fertilization in a reclaimed sandy agricultural ecosystem. PeerJ 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Yang, J.; Wang, X.; Xie, W.; Zheng, F.; Li, H.; Tang, C.; Zhu, H. Response of soil characteristics and bacterial communities to nitrogen fertilization gradients in a coastal salt-affected agroecosystem. Land Degrad. Dev. 2021, 32, 338–353. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Q.; Chen, Y.; Dai, Q.; Hu, J. Dynamic change in enzyme activity and bacterial community with long-term rice cultivation in mudflats. Curr. Microbiol. 2019, 76, 361–369. [Google Scholar] [CrossRef]
- Zhao, J.; Ni, T.; Li, Y.; Xiong, W.; Ran, W.; Shen, B.; Shen, Q.; Zhang, R. Responses of bacterial communities in arable soils in a rice-wheat cropping system to different fertilizer regimes and sampling times. PLoS ONE 2014, 9, e85301. [Google Scholar] [CrossRef]
- Fan, F.; Yin, C.; Tang, Y.; Li, Z.; Song, A.; Wakelin, S.A.; Zou, J.; Liang, Y. Probing potential microbial coupling of carbon and nitrogen cycling during decomposition of maize residue by 13C-DNA-SIP. Soil Biol. Biochem. 2014, 70, 12–21. [Google Scholar] [CrossRef]
- Zhu, J.; Peng, H.; Ji, X.; Li, C.; Li, S. Effects of reduced inorganic fertilization and rice straw recovery on soil enzyme activities and bacterial community in double-rice paddy soils. Eur. J. Soil Biol. 2019, 94, 103116. [Google Scholar] [CrossRef]
- Yi, X.; Yuan, J.; Zhu, Y.; Yi, X.; Zhao, Q.; Fang, K.; Cao, L. Comparison of the abundance and community structure of N-cycling bacteria in paddy rhizosphere soil under different rice cultivation patterns. Int. J. Mol. Sci. 2018, 19, 3772. [Google Scholar] [CrossRef] [PubMed]
- Krüger, K.; Chafee, M.; Ben Francis, T.; Glavina del Rio, T.; Becher, D.; Schweder, T.; Amann, R.I.; Teeling, H. In marine Bacteroidetes the bulk of glycan degradation during algae blooms is mediated by few clades using a restricted set of genes. ISME J. 2019, 13, 2800–2816. [Google Scholar] [CrossRef] [PubMed]
- Zeybek, N.; Rastall, R.A.; Buyukkileci, A.O. Utilization of xylan-type polysaccharides in co-culture fermentations of Bifidobacterium and Bacteroides species. Carbohydr. Polym. 2020, 236, 116076. [Google Scholar] [CrossRef]
- de Chaves, M.G.; Silva, G.G.; Rossetto, R.; Edwards, R.A.; Tsai, S.M.; Navarrete, A.A. Acidobacteria subgroups and their metabolic potential for carbon degradation in sugarcane soil amended with vinasse and nitrogen fertilizers. Front. Microbiol. 2019, 10, 1680. [Google Scholar] [CrossRef] [PubMed]
- Sathya, A.; Vijayabharathi, R.; Gopalakrishnan, S. Plant growth-promoting actinobacteria: A new strategy for enhancing sustainable production and protection of grain legumes. 3 Biotech 2017, 7, 102. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, F.; Huang, Y.; Zhou, M.; Gao, J.; Yan, T.; Sheng, H.; An, L. Sphingomonas sp. Cra20 increases plant growth rate and alters rhizosphere microbial community structure of Arabidopsis thaliana under drought stress. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Menon, R.R.; Kumari, S.; Kumar, P.; Verma, A.; Krishnamurthi, S.; Rameshkumar, N. Sphingomonas pokkalii sp. nov., a novel plant associated rhizobacterium isolated from a saline tolerant pokkali rice and its draft genome analysis. Syst. Appl. Microbiol. 2019, 42, 334–342. [Google Scholar] [CrossRef]
- Sørensen, S.R.; Ronen, Z.; Aamand, J. Isolation from agricultural soil and characterization of a Sphingomonas sp. able to mineralize the phenylurea herbicide isoproturon. Appl. Environ. Microbiol. 2001, 67, 5403–5409. [Google Scholar] [CrossRef]
- Gómez Expósito, R.; Postma, J.; Raaijmakers, J.M.; De Bruijn, I. Diversity and Activity of Lysobacter Species from Disease Suppressive Soils. Front. Microbiol. 2015, 6, 1243. [Google Scholar] [CrossRef]
- Albayrak, Ç.B. Bacillus species as biocontrol agents for fungal plant pathogens. In Bacilli and Agrobiotechnology: Phytostimulation and Biocontrol; Springer: Cham, Switzerland, 2019; pp. 239–265. [Google Scholar]
- El-Tarabily, K.A.; AlKhajeh, A.S.; Ayyash, M.M.; Alnuaimi, L.H.; Sham, A.; ElBaghdady, K.Z.; Tariq, S.; AbuQamar, S.F. Growth Promotion of Salicornia bigelovii by Micromonospora chalcea UAE1, an Endophytic 1-Aminocyclopropane-1-Carboxylic Acid Deaminase-Producing Actinobacterial Isolate. Front. Microbiol. 2019, 10, 1694. [Google Scholar] [CrossRef]
- De Schrijver, A.; De Mot, R. Degradation of pesticides by actinomycetes. Crit. Rev. Microbiol. 1999, 25, 85–119. [Google Scholar] [CrossRef]
- Hamdi, C.; Arous, F.; Jaouani, A. Actinobacteria: A Promising Source of Enzymes Involved in Lignocellulosic Biomass Conversion. Adv. Biotechnol. Microbiol. 2020, 13. [Google Scholar] [CrossRef]
- Shen, H.; Yan, W.; Yang, X.; He, X.; Wang, X.; Zhang, Y.; Wang, B.; Xia, Q. Co-occurrence network analyses of rhizosphere soil microbial PLFAs and metabolites over continuous cropping seasons in tobacco. Plant Soil 2020, 452, 119–135. [Google Scholar] [CrossRef]
- Tu, Q.; Yan, Q.; Deng, Y.; Michaletz, S.T.; Buzzard, V.; Weiser, M.D.; Waide, R.; Ning, D.; Wu, L.; He, Z.; et al. Biogeographic patterns of microbial co-occurrence ecological networks in six American forests. Soil Biol. Biochem. 2020, 148, 107897. [Google Scholar] [CrossRef]
- Ma, B.; Wang, Y.; Ye, S.; Liu, S.; Stirling, E.; Gilbert, J.A.; Faust, K.; Knight, R.; Jansson, J.K.; Cardona, C.; et al. Earth microbial co-occurrence network reveals interconnection pattern across microbiomes. Microbiome 2020, 8, 82. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, C.; Kou, Y.; Yao, M.; He, Z.; Li, X. Distinct mechanisms shape soil bacterial and fungal co-occurrence networks in a mountain ecosystem. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef] [PubMed]
- Obieze, C.C.; Chikere, C.B.; Selvarajan, R.; Adeleke, R.; Ntushelo, K.; Akaranta, O. Functional attributes and response of bacterial communities to nature-based fertilization during hydrocarbon remediation. Int. Biodeterior. Biodegradation 2020, 154, 105084. [Google Scholar] [CrossRef]
- Liljeros, F.; Edling, C.R.; Nunes Amaral, L.A.; Stanley, H.E.; Åberg, Y. Social networks: The web of human sexual contacts. Nature 2001, 411, 907–908. [Google Scholar] [CrossRef] [PubMed]
- Albert, R.; Jeong, H.; Barabási, A.L. Diameter of the world-wide web. Nature 1999, 401, 130–131. [Google Scholar] [CrossRef]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.A.; Kainz, K.; Carmona-Gutierrez, D.; Madeo, F. Microbial wars: Competition in ecological niches and within the microbiome. Microb. Cell 2018, 5, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Rang, Z.; Zhang, C.; Chen, W.; Tian, F.; Yin, H.; Dai, L. The succession pattern of soil microbial communities and its relationship with tobacco bacterial wilt. BMC Microbiol. 2016, 16, 1–10. [Google Scholar] [CrossRef]
- Banerjee, S.; Walder, F.; Büchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; van der Heijden, M.G.A. Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef]
- Xue, L.; Ren, H.; Brodribb, T.J.; Wang, J.; Yao, X.; Li, S. Long term effects of management practice intensification on soil microbial community structure and co-occurrence network in a non-timber plantation. For. Ecol. Manag. 2020, 459, 117805. [Google Scholar] [CrossRef]
- Banerjee, S.; Schlaeppi, K.; van der Heijden, M.G.A. Keystone taxa as drivers of microbiome structure and functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef]
- Xie, Y.; Wright, S.; Shen, Y.; Du, L. Bioactive natural products from Lysobacter. Nat. Prod. Rep. 2012, 29, 1277–1287. [Google Scholar] [CrossRef]
- Qin, Y.; Fu, Y.; Dong, C.; Jia, N.; Liu, H. Shifts of microbial communities of wheat (Triticum aestivum L.) cultivation in a closed artificial ecosystem. Appl. Microbiol. Biotechnol. 2016, 100, 4085–4095. [Google Scholar] [CrossRef]
- Puopolo, G.; Tomada, S.; Pertot, I. The impact of the omics era on the knowledge and use of Lysobacter species to control phytopathogenic micro-organisms. J. Appl. Microbiol. 2018, 124, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Pascual, J.; Huber, K.J.; Foesel, B.U.; Overmann, J. Stenotrophobacter. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2017; pp. 1–8. [Google Scholar]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A.; et al. Complete nitrification by Nitrospira bacteria. Nature 2015, 528, 504–509. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, M.; Zhao, Q.; Wang, F.; Gao, J.; Sheng, H.; An, L. Complete genome sequence of Sphingomonas sp. Cra20, a drought resistant and plant growth promoting rhizobacteria. Genomics 2020, 112, 3648–3657. [Google Scholar] [CrossRef] [PubMed]
- Brooks, C.N.; Field, E.K. Iron flocs and the three domains: Microbial interactions in freshwater iron mats. mBio 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Fru, E.C.; Piccinelli, P.; Fortin, D. Insights into the Global Microbial Community Structure Associated with Iron Oxyhydroxide Minerals Deposited in the Aerobic Biogeosphere. Geomicrobiol. J. 2012, 29, 587–610. [Google Scholar] [CrossRef]
- Roden, E.E. Microbial iron-redox cycling in subsurface environments. In Proceedings of the Biochemical Society Transactions; Portland Press: London, UK, 2012; Volume 40, pp. 1249–1256. [Google Scholar]
- Emerson, D.; Roden, E.; Twining, B.S. The microbial ferrous wheel: Iron cycling in terrestrial, freshwater, and marine environments. Front. Microbiol. 2012, 3, 383. [Google Scholar] [CrossRef]
- Weiss, J.V.; Rentz, J.A.; Plaia, T.; Neubauer, S.C.; Merrill-Floyd, M.; Lilburn, T.; Bradburne, C.; Megonigal, J.P.; Emerson, D. Characterization of neutrophilic Fe(II)-oxidizing bacteria isolated from the rhizosphere of wetland plants and description of Ferritrophicum radicicola gen. nov. sp. nov., and Sideroxydans paludicola sp. nov. Geomicrobiol. J. 2007, 24, 559–570. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location (Region) | Site Code | Description (Rice Cultivation Status or Time Point, RCST) | Sampling Date | Sampling Time (Planting Season) |
|---|---|---|---|---|
| Umbeluzi (Maputo) | 10YF | At least ten years without rice cultivation | May 2016 | Fallow field |
| PH | Cultivated fields with Makassane rice variety under an irrigated system. The fields were completely flooded. Samples were collected immediately after rice harvest (i.e., postharvest) | May 2016 | Postharvest (2015/2016) | |
| 1YF | One year without growing rice. Cultivated with Makassane rice variety one year prior | May 2016 | Fallow field | |
| 5mR | Same site as PH. Samples were collected five months postharvest but before pre-planting preparation for the next season. The field was completely flooded at the time of sampling | October 2016 | Pre-planting (2016/2017) | |
| Chokwe (Gaza) | 5YF | At least five years without rice cultivation. The fields were completely flooded at the time of sampling | May 2016 | Fallow field |
| PH | Cultivated fields with Makassane rice variety in an irrigated system. The fields were completely flooded. Samples were collected immediately after rice harvest (i.e., postharvest) | May 2016 | Postharvest (2015/2016) | |
| 5mR | Same site as PH. Samples were collected five months postharvest but before pre-planting preparation for the next season. The field was completely dry at the time of sampling | October 2016 | Pre-planting (2016/2017) |
| Properties | Umbeluzi | Chokwe | |||||
|---|---|---|---|---|---|---|---|
| 10YF | 1YF | PH | 5mR | 5YF | PH | 5mR | |
| pH (H2O) | 7.17 ± 0.93 b | 7.67 ± 0.48 b | 8.25 ± 0.21 a | 7.99 ± 0.49 | 7.76 ± 0.50 a | 8.08 ± 0.29 a | 7.11 ± 0.69 * |
| EC (mS/cm) | 5.18 ± 6.35 a | 0.41± 0.05 b | 0.55 ± 0.13 b | 4.45 ± 0.66 * | 1.00 ± 0.32 a | 0.63 ± 0.31 a | 5.42 ± 3.06 * |
| Moisture (g H2O kg−1) | 30.50 ± 9.18 a | 27.84 ± 8.33 a | 29.71 ± 12.18 a | 22.75 ± 21.12 | 53.35 ± 2.31 a | 56.67 ± 5.82 a | 39.47 ± 11.32 * |
| SOC (g kg−1) | 16.09 ± 5.08 a | 14.00 ± 2.25 a | 15.24 ± 1.85 a | 15.92 ± 1.44 | 20.68 ± 4.16 a | 14.44 ± 1.21 b | 16.23 ± 0.91 * |
| NH4+–N (mg N kg−1) | 33.28 ± 58.39 a | 2.13 ± 0.83 c | 6.96 ± 2.34 b | 4.73 ± 0.90 * | 6.36 ± 1.38 a | 4.34 ± 0.93 b | 10.22 ± 15.1 |
| NO3− –N (mg N kg−1) | 29.17 ± 51.46 a | 0.10 ± 0.31 b | 0.06 ± 0.19 b | 1.87 ± 4.62 * | 0.00 ± 0.00 | 0.00 ± 0.00 | 22.52 ± 25.12 * |
| Total N (g kg−1 dry soil) | 0.70 ± 0.34 a | 0.75 ± 0.19 a | 0.77 ± 0.15 a | 0.91 ± 0.12 | 1.34 ± 0.63 a | 0.77 ± 0.11 a | 1.14 ± 0.08 * |
| P-Olsen (mg kg−1) | 28.30 ± 8.86 a | 26.44 ± 6.59 a | 29.87 ± 8.22 a | 20.81 ± 1.65 | 15.42 ± 3.91 a | 9.67 ± 3.58 a | 9.39 ± 1.67 |
| Egner Rhiem K (mg kg−1) | 240.70 ± 45.21 a | 251.37 ± 35.77 a | 239.25 ± 33.92 a | 328.59 ± 60.59 * | 463.61 ± 104.05 a | 524.98 ± 60.14 a | 492.85 ± 249.28 |
| Location | RCST | β-Glucosidase (µg p- Nitrophenol g−1 h−1) | Acid Phosphatase (µg p- Nitrophenol g−1 h−1) | Alkaline Phosphatase (µg p- Nitrophenol g−1 h−1) | Urease (mg N–NH4+ kg−1 2 h−1) |
|---|---|---|---|---|---|
| Umbeluzi | 10YF | 41.18 ± 25.55 ab | 303.54 ± 201.14 a | 218.68 ± 104.83 a | 49.59 ± 32.97 a |
| 1YF | 43.99 ± 14.5 a | 188.31 ± 57.79 a | 170.91 ± 31.69 a | 42.67 ± 8.97 a | |
| PH | 25.83 ± 3.68 b | 127.15 ± 31.99 a | 148.68 ± 39.31 a | 25.11 ± 11.33 a | |
| 5mR | 30.50 ± 6.69 * | 122.98 ± 38.36 | 121.43 ± 45.49 | 24.34 ± 11.64 | |
| Chokwe | 5YF | 60.70 ± 23.99 a | 351.9 ± 111.1 a | 240.51 ± 69.41 a | 71.22 ± 19.47 a |
| PH | 58.51 ± 9.97 a | 198.37 ± 44.24 b | 172.99 ± 43.16 a | 82.00 ± 21.10 a | |
| 5mR | 74.26 ± 12.91 * | 220.03 ± 27.04 | 191.61 ± 27.48 | 82.30 ± 20.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ezeokoli, O.T.; Nuaila, V.N.A.; Obieze, C.C.; Muetanene, B.A.; Fraga, I.; Tesinde, M.N.; Ndayiragije, A.; Coutinho, J.; Melo, A.M.P.; Adeleke, R.A.; et al. Assessing the Impact of Rice Cultivation and Off-Season Period on Dynamics of Soil Enzyme Activities and Bacterial Communities in Two Agro-Ecological Regions of Mozambique. Agronomy 2021, 11, 694. https://doi.org/10.3390/agronomy11040694
Ezeokoli OT, Nuaila VNA, Obieze CC, Muetanene BA, Fraga I, Tesinde MN, Ndayiragije A, Coutinho J, Melo AMP, Adeleke RA, et al. Assessing the Impact of Rice Cultivation and Off-Season Period on Dynamics of Soil Enzyme Activities and Bacterial Communities in Two Agro-Ecological Regions of Mozambique. Agronomy. 2021; 11(4):694. https://doi.org/10.3390/agronomy11040694
Chicago/Turabian StyleEzeokoli, Obinna T., Valter N. A. Nuaila, Chinedu C. Obieze, Belo A. Muetanene, Irene Fraga, Maria Natalia Tesinde, Alexis Ndayiragije, João Coutinho, Ana M. P. Melo, Rasheed A. Adeleke, and et al. 2021. "Assessing the Impact of Rice Cultivation and Off-Season Period on Dynamics of Soil Enzyme Activities and Bacterial Communities in Two Agro-Ecological Regions of Mozambique" Agronomy 11, no. 4: 694. https://doi.org/10.3390/agronomy11040694
APA StyleEzeokoli, O. T., Nuaila, V. N. A., Obieze, C. C., Muetanene, B. A., Fraga, I., Tesinde, M. N., Ndayiragije, A., Coutinho, J., Melo, A. M. P., Adeleke, R. A., Ribeiro-Barros, A. I., & Fangueiro, D. (2021). Assessing the Impact of Rice Cultivation and Off-Season Period on Dynamics of Soil Enzyme Activities and Bacterial Communities in Two Agro-Ecological Regions of Mozambique. Agronomy, 11(4), 694. https://doi.org/10.3390/agronomy11040694