
Identification of Rhizospheric Microorganisms That Manages Root Knot Nematode and Improve Oil Yield in Sweet Basil (Ocimum basilicum L.)



Abstract
1. Introduction
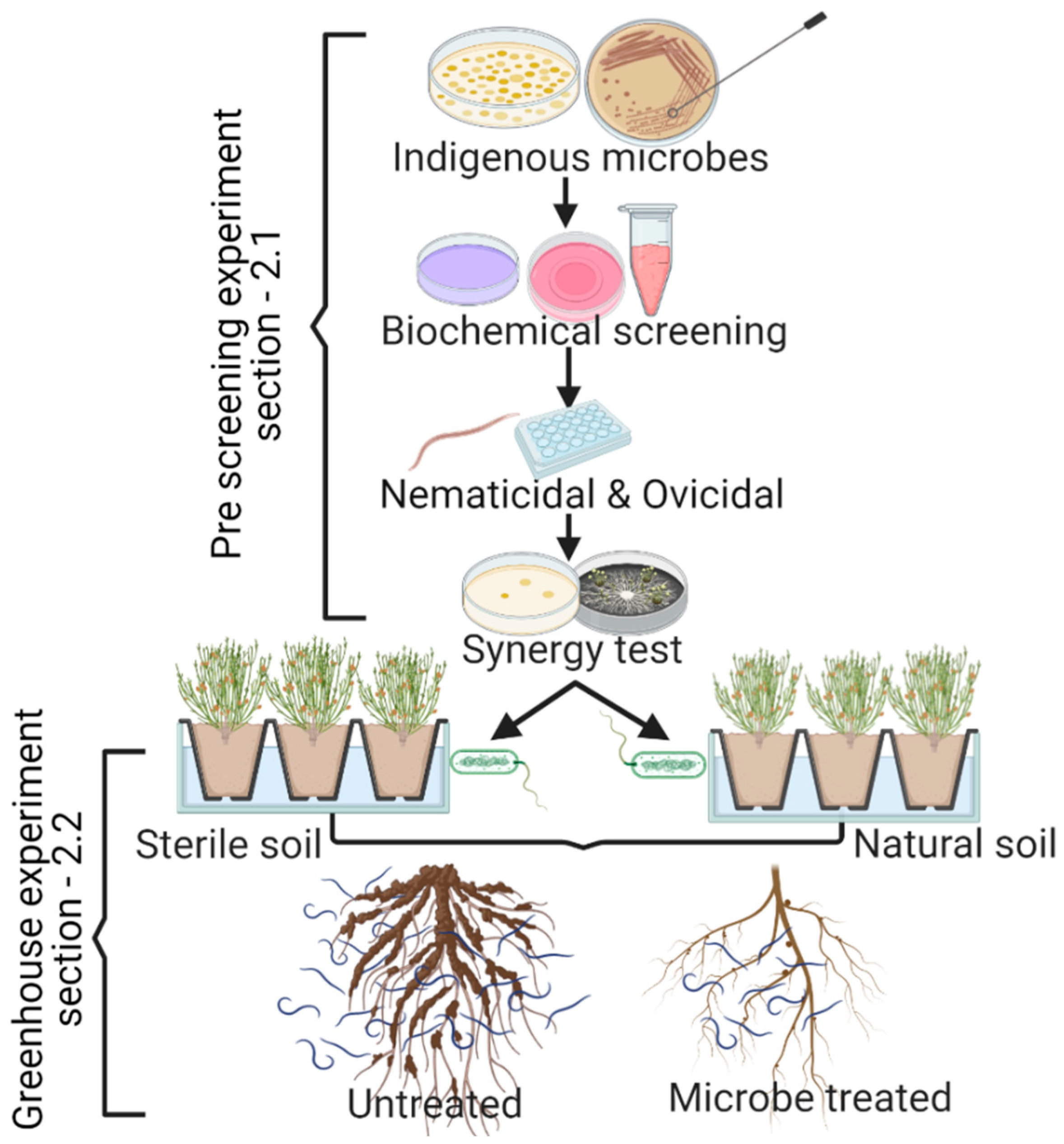
2. Material and Methods
2.1. Prescreening Experiments
2.1.1. Isolation of Bioagents
2.1.2. Biochemical Characterization, Plant Growth Promoting (“PGP”) Activity and Selection of Bioinoculants
Cellulose and Protease Activity
Estimation of Chitinases and Pectinases Activity
Gelatinase and Asparaginase Activity
PGP Activity
2.1.3. Prescreening of Bioinoculants for the Management of M. incognita in Basil Plants
2.1.4. Molecular Identification of the Selected Strains
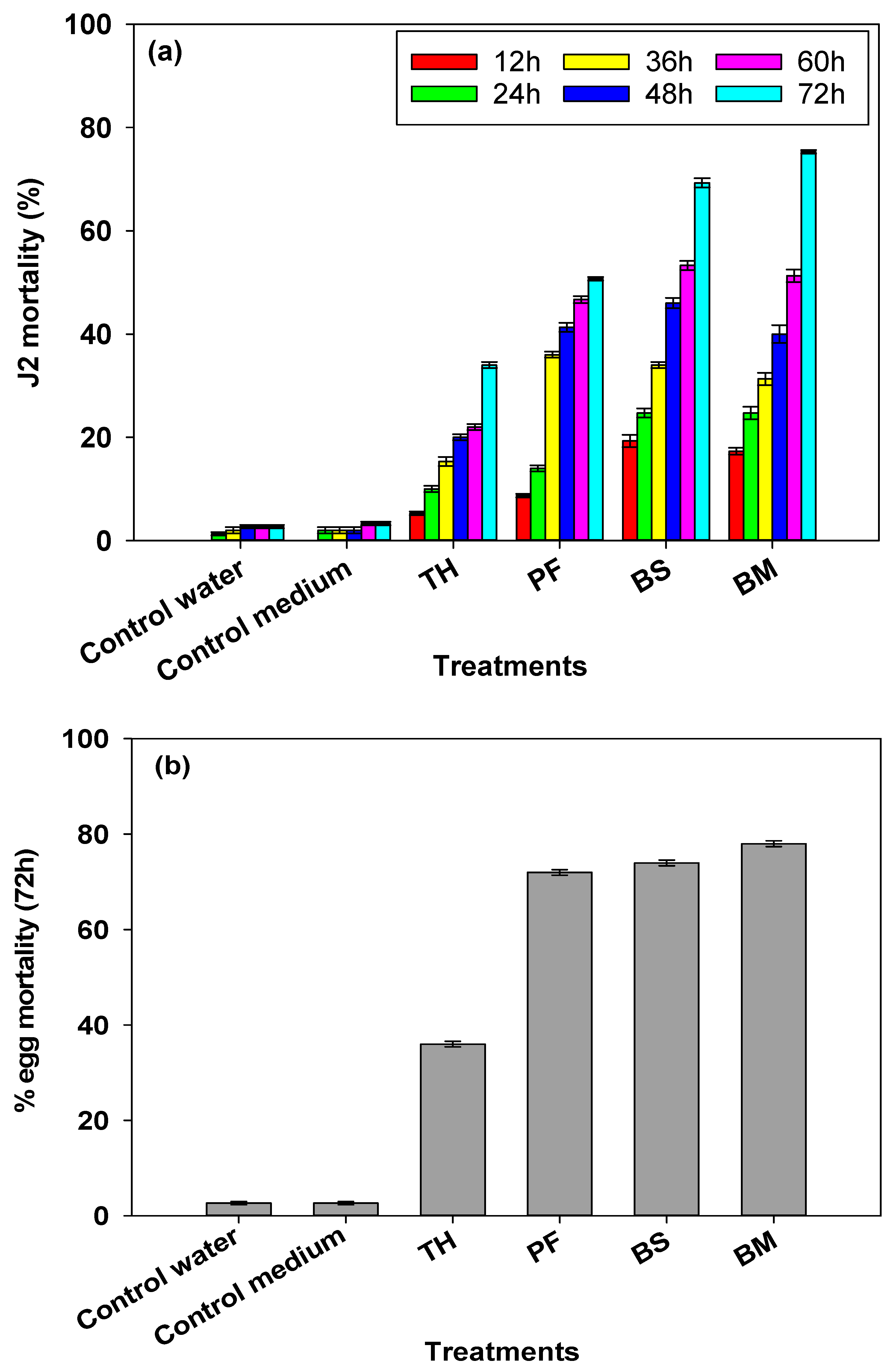
2.1.5. In Vitro Nematicidal and Ovicidal Activity of Bioinoculants
2.1.6. In Vitro Synergy Test
2.2. Greenhouse Experiments
2.2.1. Preparation of Fungal and Bacterial Strains for Rhizospheric Inoculation
2.2.2. Testing the Effects of Microbial Bioinoculants on Nematode Population and Growth Parameters of Basil Plants in a Greenhouse
2.2.3. Estimation of Nematode Management Potentials of Bioinoculants
2.2.4. Plant Yield and Essential Oil Extraction
2.2.5. Gas Chromatography Analysis of the Essential Oil
2.2.6. Nutrient Uptake Analysis
2.3. Statistical Analysis
3. Results
3.1. Prescreening Experiments
3.1.1. Biochemical Characterization and PGP Activity of Isolates and Selection of Bioinoculants
3.1.2. Prescreening of Bioinoculants for the Management of M. incognita in Basil Plants
3.1.3. Molecular Identification of Bioinoculants
3.1.4. In Vitro Nematicidal and Ovicidal Activity of Bioinoculants
3.2. Greenhouse Experiments
3.2.1. Effects of Microbial Bioinoculants on Growth Parameters of Basil Plants
3.2.2. Effects of Bioinoculants on Biomass Yield of Basil Plants
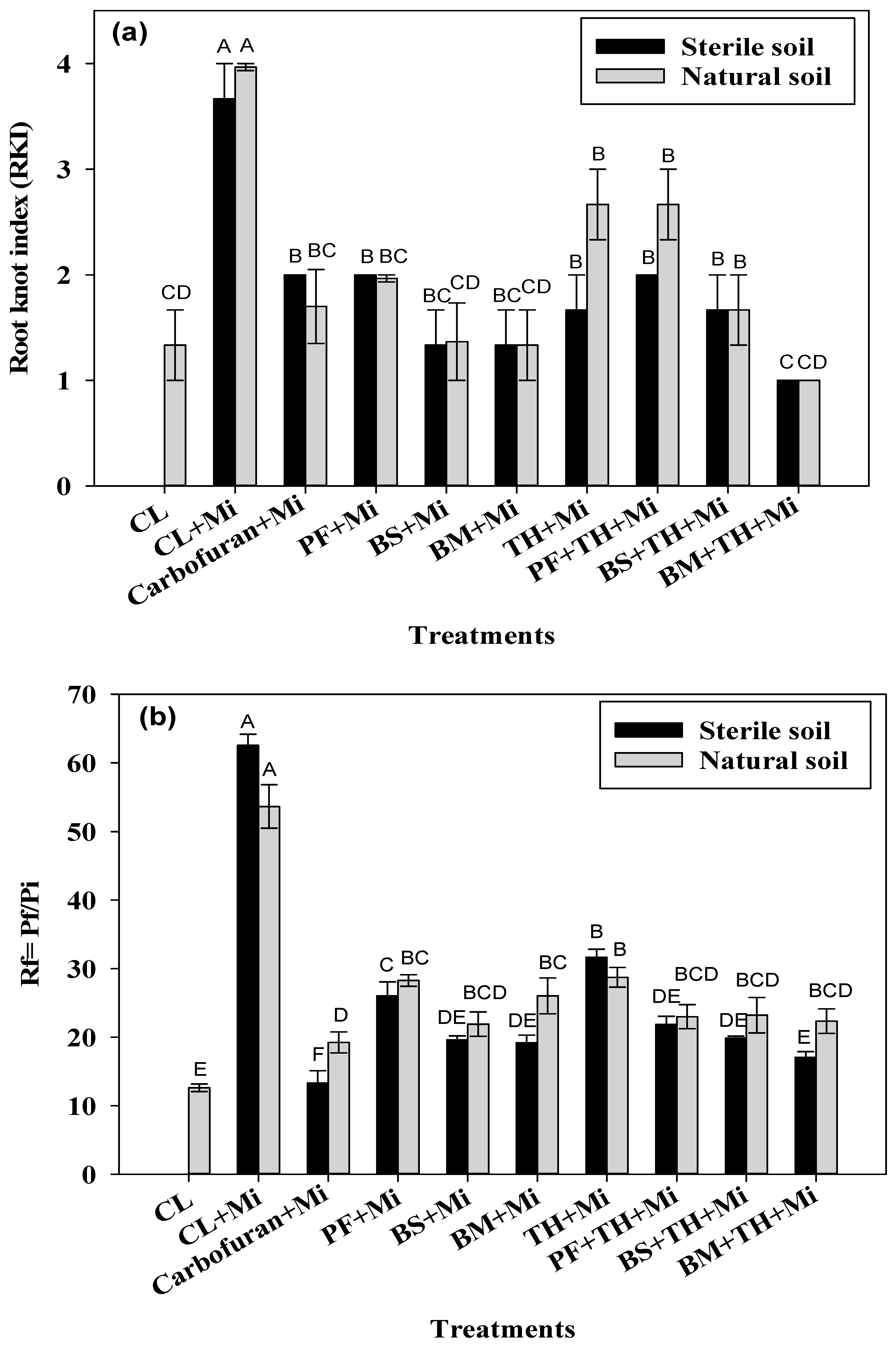
3.2.3. Estimation of Nematode Management Potentials of Bioinoculants
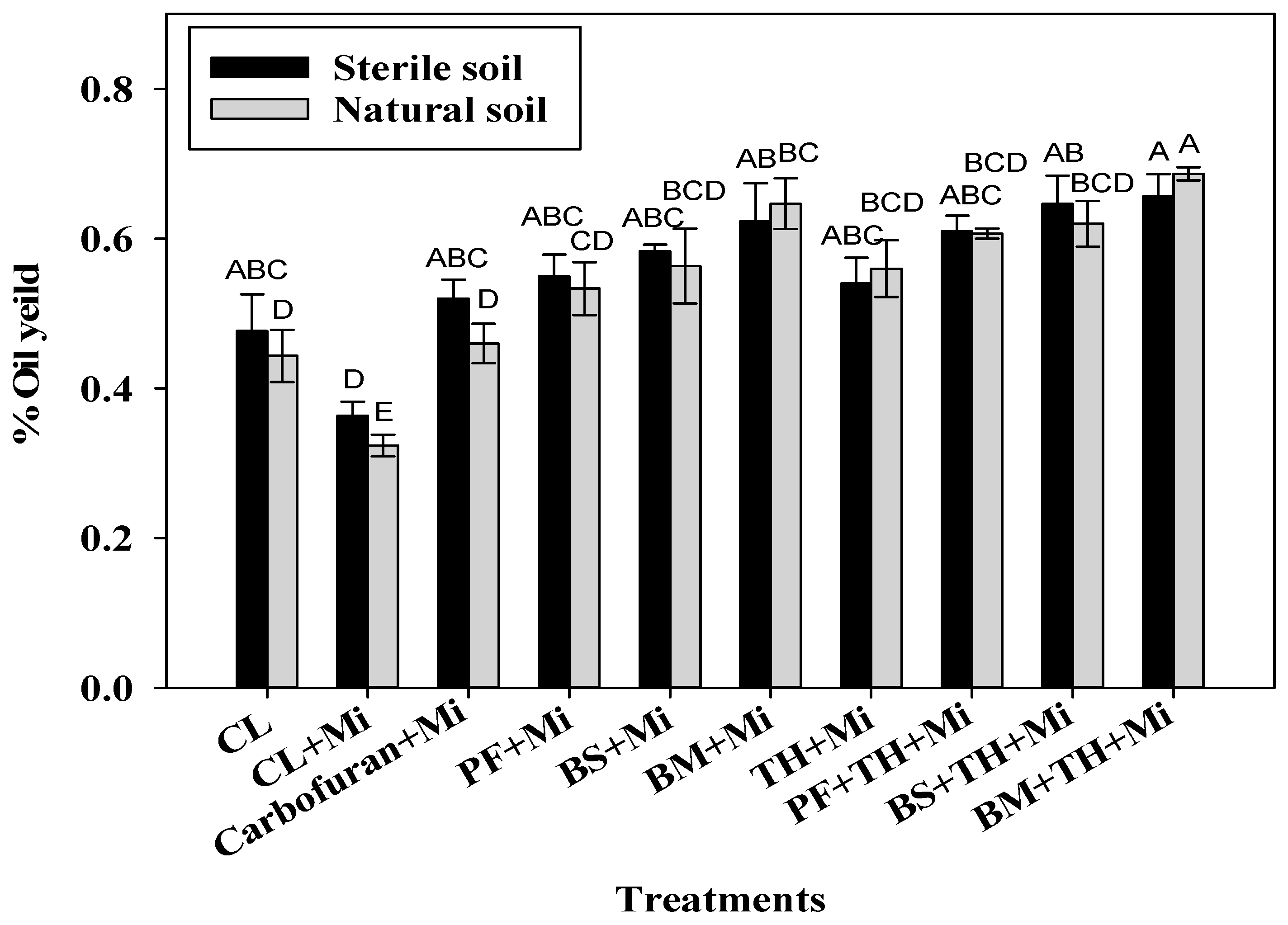
3.2.4. Effect of Bioinoculants on Essential Oil
3.2.5. Effect of bioinoculants on plant nutrient uptake.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saikia, K.S.; Tiwari, S.; Pandey, R. Rhizospheric biological weapons for growth enhancement and Meloidogyne incognita management in Withania somnifera cv. Poshita. Biol. Control. 2013, 65, 225–234. [Google Scholar] [CrossRef]
- Tiwari, S.; Saikia, K.S.; Singh, R.; Singh, S.P.; Pandey, R. Native microbial inoculants for the management of Meloidogyne incognita in Withania somnifera cv. Poshita. Proc. Natl. Acad. Sci. India. Sect B Biol. Sci. 2014, 86, 55–63. [Google Scholar] [CrossRef]
- Gupta, R.; Pandey, R. Microbial interference ameliorates essential oil yield and diminishes root-knot infestation in sweet basil under field conditions. Biocontrol Sci. Technol. 2015, 25, 1165–1179. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Sreeramu, B.S. Cultivation of Medicinal and Aromatic Crops; University Press: Hyderabad, India, 2001. [Google Scholar]
- Tiwari, S.; Pandey, S.; Singh, P.C.; Pandey, R. Biocontrol agents in co-inoculation manages root knot nematode Meloidogyne incognita (Kofoid & White) Chitwood and enhances essential oil content in Ocimum basilicum L. Ind. Crops Prod. 2017, 97, 292–301. [Google Scholar] [CrossRef]
- Telci, I.; Bayram, E.; Yilmaz, G.; Avci, B. Variability in essential oil composition of Turkish basils (Ocimum basilicum L.). Biochem. Syst. Ecol. 2006, 34, 489–497. [Google Scholar] [CrossRef]
- Singh, R.; Soni, S.K.; Patel, R.K.; Kalra, A. Technology for improving essential oil yield of Ocimum basilicum L. (sweet basil) by application of bioinoculant colonized seeds under organic field conditions. Ind. Crop. Prod. 2013, 45, 335–342. [Google Scholar] [CrossRef]
- Phippen, W.B.; Simon, J.E. Shoot regeneration of young leaf explants from basil (Ocimum basilicum L.). In Vitro Cell. Dev. Biol. Plant 2000, 36, 250–254. [Google Scholar] [CrossRef]
- Paula, J.P.de.; Gomes-Carneiro, M.R.; Paumgartten, F.J.R. Chemical composition, toxicity and mosquito repellency of Ocimum selloi oil. J. Ethnopharmacol. 2003, 88, 253–260. [Google Scholar] [CrossRef]
- Santos, B.C.S.; Pires, A.S.; Yamamoto, C.H.; Couri, M.R.C.; Taranto, A.G.; Alves, M.S.; de Matos Araujo, A.L.D.S.; de Sousa, O.V. Methyl Chavicol and Its Synthetic Analogue as Possible Antioxidant and Antilipase Agents Based on the In Vitro and In Silico Assays. Oxidative Med. Cell. Longev. 2018, 2018, 2189348. [Google Scholar] [CrossRef] [PubMed]
- Jnanesha, A.C.; Kumar, A.; Kumar, V.M. Effect of seasonal variation on growth and oil yield in Ocimum africanum Lour. J. Pharmacogn. Phytochem. 2018, 7, 73–77. [Google Scholar]
- Sajjadi, S.E. Analysis of the essential oils of two cultivated basil (Ocimum basilicum L.) from Iran. Daru J. Pharm. Sci. 2006, 14, 128–130. [Google Scholar]
- Padalia, R.C.; Verma, R.S.; Upadhyay, R.K.; Chauhan, A.; Singh, V.R. Productivity and essential oil quality assessment of promising accessions of Ocimum basilicum L. from north India. Ind. Crops. Prod. 2017, 97, 79–86. [Google Scholar] [CrossRef]
- Pandey, R.; Kalra, A.; Gupta, M.L. Evaluation of bio-agents and pesticide on root-knot nematode development and oil yield of patchouli. Arch. Phytopathol. Plant Prot. 2007, 42, 419–423. [Google Scholar] [CrossRef]
- Melakeberhan, H.; Webster, J.M.; Brooke, R.C. Improved techniques for measuring the CO2 exchange rate of Meloidogyne nematode bean plants. Nematologica 1984, 30, 213–221. [Google Scholar] [CrossRef]
- Singh, R.; Tiwari, S.; Patel, R.P.; Soni, S.; Kalra, A. Bioinoculants and AM fungus colonized nursery improved management of complex root disease of Coleus forskohlii Briq. under field conditions. Biol. Control. 2018, 122, 11–17. [Google Scholar] [CrossRef]
- Saikia, S.K.; Tiwari, S.; Pandey, R. Rhizospheric innovations for growth enhancement and Meloidogyne incognita management in Mentha arvensis cv. Kosi. Int. J. Environ. Sci. Eng. Res. 2012, 3, 26–34. [Google Scholar]
- Adam, M.; Westphal, A.; Hallmann, J.; Heuer, H. Specific Microbial Attachment to Root Knot Nematodes in Suppressive Soil. Appl. Environ. Microbiol. 2014, 80, 2679–2686. [Google Scholar] [CrossRef]
- Goswami, D.; Dhandhukia, P.; Patel, P.; Thakker, J.N. Screening of PGPR from saline desert of Kutch: Growth promotion in Arachis hypogea by Bacillus licheniformis A2. Microbiol. Res. 2014, 169, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Guerrieri, M.C.; Fanfoni, E.; Fiorini, A.; Trevisan, M.; Puglisi, E. Isolation and Screening of Extracellular PGPR from the Rhizosphere of Tomato Plants after Long-Term Reduced Tillage and Cover Crops. Plants 2020, 9, 668. [Google Scholar] [CrossRef]
- Tran, D.M.; Sugimoto, H.; Nguyen, D.A.; Watanabe, T.; Suzuki, K. Identification and characterization of chitinolytic bacteria isolated from a freshwater lake. Biosci. Biotechnol. Biochem. 2018, 82, 343–355. [Google Scholar] [CrossRef]
- Bric, J.M.; Bostock, R.M.; Silverstone, S.E. Rapid in situ assay for indole acetic acid production by bacteria immobilized on nitrocellulose membrane. Appl. Environ. Microbiol. 1991, 57, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Pikovaskya, R.I. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Hussey, R.S.; Barker, K.R. A comparison of methods of collecting inocula of Meloidogyne spp., including a new technique. Plant Dis. Rep. 1973, 57, 1025–1028. [Google Scholar]
- Edwards, U.; Rogall, T.; Blocker, H.; Emde, M.; Bottger, E.C. Isolation and direct nucleotide determination of entire genes: Characterization of a gene encoding for 16S ribosomal RNA. Nucl. Acids Res. 1989, 17, 7843–7850. [Google Scholar] [CrossRef]
- Tiwari, S.; Singh, S.; Pandey, P.; Saikia, S.K.; Negi, A.S.; Gupta, S.K.; Pandey, R.; Banerjee, S. Isolation, structure determination and anti-aging effects of 2, 3-pentanediol from endophytic fungus of Curcuma amada and docking studies. Protoplasma 2014. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Cayrol, J.C.; Djian, C.; Pijarowski, L. Study of the nematicidal properties of the culture filtrate of the nematophagous fungus Paecilomyces lilacinus. Rev. Nematol. 1989, 12, 331–336. [Google Scholar]
- Katiyar, V.; Goel, R. Improved Plant Growth from Seed Bacterization Using Siderophore Overproducing Cold Resistant Mutant of Pseudomonas fluorescens. J. Microbiol. Biotechnol. 2004, 14, 653–657. [Google Scholar]
- Lee, Y.S.; Kim, J.; Shin, S.C.; Lee, S.G.; Park, I.K. Antifungal activity of Myrtaceae essential oils and their components against three phytopathogenic fungi. Flavour Fragr. J. 2008, 23, 23–28. [Google Scholar] [CrossRef]
- Singh, R.; Divya, S.; Ashutosh, A.; Kalra, A. Technology for efficient and successful delivery of vermicompost colonized bioinoculants in Pogostemon cablin (patchouli) Benth. World. J. Microbiol. Biotechnol. 2012. [Google Scholar] [CrossRef]
- Pandey, R. Management of Meloidogyne incognita in Artemisia pallens with bio-organics. Phytoparasitica 2005, 33, 304–308. [Google Scholar] [CrossRef]
- Pandey, R. Chemical activators: A novel and sustainable management approach for Meloidogyne incognita (Kofoid and White) Chitwood in Chamomilla recutita L. Arch. Phytopathol. Plant Prot. 2005, 38, 107–111. [Google Scholar] [CrossRef]
- Krusberg, L.R.; Nielsen, L.W. Pathogenesis of root-knot nematodes to PortoRico variety of sweet potato. Phytopathology 1958, 48, 30–39. [Google Scholar]
- Goodey, T. Laboratory Methods for Work with Plant and Soil Nematodes; Southey, J.F., Ed.; Reference Book; Ministry of Agriculture, Fisheries and Food: London, UK, 1986.
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing: Carol Stream, IL, USA, 2007. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall of Englewood Cliffs: Englewood Cliffs, NJ, USA, 1967. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall of Englewood Cliffs: Englewood Cliffs, NJ, USA, 1973. [Google Scholar]
- Abd-El-Khair, H.; El-Nagdi, W.M.; Youssef, M.M.; Abd-Elgawad, M.M.; Dawood, M.G. Protective effect of Bacillus subtilis, B. pumilus, and Pseudomonas fluorescens isolates against root knot nematode Meloidogyne incognita on cowpea. Bull. Nat. Res. Cent. 2019, 43, 1–7. [Google Scholar] [CrossRef]
- Singh, R.; Paramaeswarn, T.N.; Rao, P.E.V.S.; Puttanna, K.; Kalra, A.; Srinivas, K.V.N.S.; Bagyaraj, D.J.; Divya, S. Effect of arbuscular mycorrhizal fungi and Pseudomonas fluorescens on root-rot/wilt, growth and yield of Coleus forskohlii. Biocontrol. Sci. Technol. 2009, 19, 835–841. [Google Scholar] [CrossRef]
- Ordookhani, K.; Sharafzadeh, S.H.; Zare, M. Influence of PGPR on growth, essential oil and nutrients uptake of Sweet basil. Adv. Environ. Biol. 2011, 5, 672–677. [Google Scholar]
- Pandey, R.; Mishra, A.K.; Tiwari, S.; Kalra, A. Nematode inhibiting organic materials and a strain of Trichoderma harzianum effectively manages Meloidogyne incognita in Withania somnifera fields. Biocontrol. Sci. Tech. 2011, 21, 1495–1499. [Google Scholar] [CrossRef]
- Kavitha, P.G.; Jonathan, E.I.; Nakkeeran, S. Effects of crude antibiotic of Bacillus subtilis on hatching of eggs and mortality of juveniles of Meloidogyne incognita. Nematol Medit. 2012, 40, 203–206. [Google Scholar]
- Gupta, R.; Tiwari, S.; Saikia, K.S.; Shukla, V.; Singh, R.; Singh, S.P.; Kumar, A.P.V.; Pandey, R. Exploitation of microbes for enhancing bacoside content and reduction of Meloidogyne incognita infestation in Bacopa monnieri L. Protoplasma 2014. [Google Scholar] [CrossRef]
- Abd-El-Khair, H.; El-Nagdi, W.M.A.; Ameen, H.H. Antagonistic effects of rhizobacteria isolates against Meloidogyne incognita infecting tomato plants under greenhouse conditions. Int. J. Pharm. Tech. Res. 2016, 9, 97–107. [Google Scholar]
- Goldman, G.H.; Hayes, C.H.; Harman, G.E. Molecular and cellular biology of biocontrol by Trichoderma spp. Trends Biotechnol. 1994, 12, 478–482. [Google Scholar] [CrossRef]
- Lorito, M.; Peterbauer, C.; Hayes, C.K.; Harman, G.E. Synergistic interaction between fungal cell-wall degrading enzymes and different antifungal compounds enhances inhibition of spore germination. Microbiology 1994, 140, 623–629. [Google Scholar] [CrossRef]
- Sharon, E.; Chet, I.; Spiegel, Y. Improved attachment and parasitism of Trichoderma on Meloidogyne javanica in vitro. Eur. J. Plant Pathol. 2009, 123, 291–299. [Google Scholar] [CrossRef]
- Wang, M.; De Deyn, G.B.; Bezemer, T.M. Separating effects of soil microorganisms and nematodes on plant community dynamics. Plant Soil. 2019, 441, 455–467. [Google Scholar] [CrossRef]
- Poveda, J.; Abril-Urias, P.; Escobar, C. Biological Control of Plant-Parasitic Nematodes by Filamentous Fungi Inducers of Resistance: Trichoderma, Mycorrhizal and Endophytic Fungi. Front. Microbiol. 2020, 11, 992. [Google Scholar] [CrossRef]
- Timper, P.; Koné, D.; Yin, J.; Ji, P.; Gardener, B.B.M. Evaluation of an antibiotic-producing strain of Pseudomonas fluorescens for suppression of plant-parasitic nematodes. J. Nematol. 2009, 41, 234–240. [Google Scholar] [PubMed]
- Lu, H.; Wang, X.; Zhang, K.; Xu, Y.; Zhou, L.; Li, G. Identification and nematicidal activity of bacteria isolated from cow dung. Ann. Microbiol. 2014, 64, 407–411. [Google Scholar] [CrossRef]
- Liu, G.; Lin, X.; Xu, S.; Liu, G.; Liu, F.; Mu, W. Screening, identification and application of soil bacteria with nematicidal activity against root-knot nematode (Meloidogyne incognita) on tomato. Pest. Manag. Sci. 2020, 76, 2217–2224. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Sharma, S.; Sharma, S.; Kumar, V. Synergistic effect of bio-inoculants on yield, nodulation and nutrient uptake of chickpea (Cicer arietinum L.) under rainfed conditions. J. Plant Nutr. 2019, 42, 374–383. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]
- Lemberkovics, E.; Petri, G.; Nguyen, H.; Mathe, L. Relationships between essential oil and flavonoid biosynthesis in basil. Acta Hort. 1995, 426, 647–655. [Google Scholar] [CrossRef]
- Tian, B.; Yang, J.; Zhang, K.Q. Bacteria used in the biological control of plant-parasitic nematodes: Populations, mechanisms of action, and future prospects. FEMS Microbiol. Ecol. 2007, 61, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Lugtenberg, B.; Kamilova, F. Plant growth promoting rhizobacteria. Annu. Rev Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Keneni, A.; Assefa, F.; Parbu, P.C. Isolation of phosphate solubilizing bacteria from the rhizosphere of faba bean of Ethiopia and their abilities on solubilizing insoluble phosphates. J. Agric. Sci. Technol. 2010, 12, 79–89. [Google Scholar]
- Norabadia, M.T.; Sahebania, N.; Etebarianb, H.R. Biological control of root-knot nematode (Meloidogyne javanica) disease by Pseudomonas fluorescens (Chao). Arch. Phytopathol. Plant Prot. 2014, 47, 615–621. [Google Scholar] [CrossRef]
- Rika, A.I.; Nyoman, P.A.; Tati, S.S. Role of Indigenous Rhizosphere Bacteria in Suppressing Root-knot Nematode and Improve Plant Growth Tomato. Plant Pathol. J. 2017, 16, 25–32. [Google Scholar]
- Pandey, R.; Gupta, A.; Haider, R.; Kalra, A. Management of root-knot nematode in Ocimum basilicum through bio-organics. J. Mycol. Plant Pathol. 2009, 39, 86–89. [Google Scholar]
- Pandey, R.; Gupta, A.; Singh, H.N.; Kalra, A. Phytonematode management through bacteria: An underground battle for existence. In Recent Advances in Biopesticides Biotechnological Applications; Johri, J.K., Ed.; NBRI: Lucknow, India, 2009; pp. 1–26. [Google Scholar]
- Singh, R.; Patel, R.P.; Singh, D.; Soni, S.K.; Tiwari, S. Bioinoculant coated seed improved the growth and yield of Withania somnifera (L.) Dunal. Medicinal Plants. Int. J. Phytomedicines Relat. Ind. 2018, 10, 191–195. [Google Scholar] [CrossRef]
- Reimann, S.; Hauschild, R.; Hildebrandt, U.; Sikora, R.A. Interrelationships between Rhizobium etli G12 and Glomus intraradices and multitrophic effects in the biological control of the root-knot nematode Meloidogyne incognita on tomato. J. Plant Dis. Prot. 2008, 115, 108–113. [Google Scholar] [CrossRef]
- Timper, P. Conserving and enhancing biological control of nematodes. J. Nematol. 2014, 46, 75–89. [Google Scholar]
- Chen, S.; Dickson, D.W.; Kimbrough, J.W.; McSorley, R.; Mitchell, D.J. Fungi associated with females and cysts of Heterodera glycines in a Florida soybean field. J. Nematol. 1994, 26, 296–303. [Google Scholar]
- Topalović, O.; Hussain, M.; Heuer, H. Plants and Associated Soil Microbiota Cooperatively Suppress Plant-Parasitic Nematodes. Front. Microbiol. 2020, 11, 313. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Feng, H.; Schuelke, T.; De Santiago, A.; Zhang, Q.; Zhang, J.; Luo, C.; Wei, L. Rhizosphere Microbiomes from Root Knot Nematode Non-infested Plants Suppress Nematode Infection. Microb. Ecol. 2019, 78, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Colagiero, M.; Rosso, L.C.; Ciancio, A. Diversity and biocontrol potential of bacterial consortia associated to root-knot nematodes. Biol. Control. 2017, 120, 11–16. [Google Scholar] [CrossRef]
- Rhoades, H.L. Effects of fallowing, summer cover crops, and fenamiphos on nematode populations and yields in a cabbage-field corn rotation in Florida. Nematropica 1984, 14, 131–138. [Google Scholar]
- Ferraz, H.B.; Bertolucci, P.H.F.; Pereira, J.S.; Lima, J.G.C.; Andrade, L.A.F. Chronic exposure to the fungicide maneb may produce symptoms and signs of CNS manganese intoxication. Neurology 1988, 38, 550–553. [Google Scholar] [CrossRef]
- Oteino, R.D.; Lally, S.; Kiwanuka, A.; Lloyd, D.; Ryan, K.J.; Germaine, D.N. Dowling Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Naveed, M.; Ali, M.; Zahir, Z.A.; Nadeem, S.M.; Yaseen, M.; Meena, V.S.; Farooq, M.; Singh, R.; Rahman, M.; et al. Perspectives of potassium solubilizing microbes in sustainable food production system: A review. Appl. Soil Ecol. 2019, 133, 146–159. [Google Scholar] [CrossRef]
- He, Y.; Pantigoso, H.; Wu, Z.; Vivanco, J. Co-inoculation of Bacillus sp. and Pseudomonas putida at different development stages acts as a biostimulant to promote growth, yield and nutrient uptake of tomato. J. Appl. Microbiol. 2019, 127, 196–207. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Abbreviation Used |
|---|---|
| Untreated control | CL |
| untreated inoculated control + M. incognita | CL + Mi |
| Carbofuran + M. incognita | Carbofuran +Mi |
| Pseudomonas fluorescence + M. incognita | PF + Mi |
| Bacillus subtilis + M. incognita | BS + Mi |
| Bacillus megaterium + M. incognita | BM + Mi |
| Trichoderma harzianum + M. incognita | TH + Mi |
| P. fluorescence + T. harzianum + M. incognita | PF + TH + Mi |
| B. subtilis + T. harzianum+ M. incognita | BS + TH + Mi |
| B. megaterium + T. harzianum+ M. incognita | BM + TH + Mi |
| Treatments | Plant Height (cm2) | Canopy (cm2) | Dry Herb Yield (g pot−1) | Total Dry Herb Yield (g pot−1) | Root Knot Index (RKI) | ||
|---|---|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | ||||
| UC | 62.33 ± 1.45 ef | 7.33 ± 0.88 b | 24.66 ± 1.76 ab | 12.93 ± 0.42 cd | 1.45 ± 0.05 bc | 14.39 ± 0.47 cd | 0.00 ± 0.00 d |
| UIC | 42.33 ± 0.88 g | 5.00 ± 0.58 c | 15.33 ± 2.02 c | 9.62 ± 0.45 f | 1.16 ± 0.09 d | 10.78 ± 0.54 f | 3.67 ± 0.33 a |
| CAB | 60.33 ± 0.89 f | 9.50 ± 0.29 ab | 3.67 ± 0.66 ab | 11.85 ± 0.38 e | 1.48 ± 0.09 bc | 13.34 ± 0.48 e | 1.67 ± 0.31 c |
| RPN 1 | 72.33 ± 3.84 ab | 8.50 ± 0.77 ab | 26.33 ± 1.02 ab | 12.4 ± 0.14 de | 1.50 ± 0.04 bc | 13.97 ± 0.28 de | 2.67 ± 0.33 b |
| RPN 2 | 76.00 ± 1.51 a | 9.167 ± 0.44 ab | 23.00 ± 0.57 ab | 14.05 ± 0.23 ab | 1.76 ± 0.05 a | 15.81 ± 0.05 ab | 1.67 ± 0.31 c |
| RPN 3 | 75.67 ± 1.86 a | 10.40 ± 0.49 a | 27.00 ± 0.50 a | 14.57 ± 0.047 a | 1.70 ± 0.05 ab | 16.27 ± 0.20 a | 2.00 ± 0.00 bc |
| RPN 4 | 70.00 ± 0.57 bc | 9.33 ± 0.33 ab | 24.67 ± 1.21 ab | 13.07 ± 0.07 cd | 1.56 ± 0.08 ab | 14.64 ± 0.11 cd | 2.33 ± 0.00 bc |
| RPN 5 | 70.67 ± 1.76 ab | 9.00 ± 0.57 ab | 22.67 ± 1.15 ab | 13.13 ± 0.03 cd | 1.51 ± 0.08 bc | 14.64 ± 0.41 cd | 2.33 ± 0.00 bc |
| MTN 2 | 67.67 ± 0.88 cd | 8.67 ± 0.66 ab | 24.00 ± 1.52 ab | 12.67 ± 0.32 cd | 1.43 ± 0.05 c | 14.06 ± 0.11 de | 2.67 ± 0.33 b |
| MTN 100 | 67.00 ± 1.52 de | 8.33 ± 1.20 ab | 24.00 ± 0.57 ab | 13.07 ± 0.06 cd | 1.56 ± 0.10 ab | 14.64 ± 0.16 cd | 1.33 ± 0.31 c |
| MTN 101 | 74.33 ± 1.21 ab | 9.33 ± 0.72 ab | 25.33 ± 0.88 ab | 13.47 ± 0.22 bc | 1.69 ± 0.10 ab | 15.13 ± 0.32 bc | 2.00 ± 0.00 bc |
| MTNK4 | 72.67 ± 1.20 ab | 9.33 ± 0.73 ab | 25.67 ± 0.66 ab | 13.07 ± 0.06 cd | 1.49 ± 0.08 bc | 14.57 ± 0.17 cd | 1.33 ± 0.31 c |
| MTN 121 | 74.33 ± 1.22 ab | 9.33 ± 0.77 ab | 22.00 ± 2.51 b | 13.09 ± 0.07 cd | 1.53 ± 0.05 ab | 14.62 ± 0.16 cd | 2.00 ± 0.00 bc |
| Treatments | Sterile Soil | Natural Soil | ||
|---|---|---|---|---|
| Shoot Dry wt. (g pot−1) | Root Dry wt. (g pot−1) | Shoot Dry wt. (g pot−1) | Root Dry wt. (g pot−1) | |
| CL | 20.32 ± 0.48 d | 4.70 ± 0.15 cd | 18.10 ± 0.76 de | 3.98 ± 0.56 bc |
| CL + Mi | 13.43 ± 0.67 e | 2.01 ± 0.06 f | 11.36 ± 0.73 f | 3.18 ± 0.13 c |
| Carbofuran + Mi | 22.41 ± 0.48 cd | 4.16 ± 0.42 de | 17.43 ± 0.34 e | 4.59 ± 0.23 b |
| PF + Mi | 24.02 ± 0.63 bc | 4.85 ± 0.28 cd | 20.16 ± 0.55 cd | 5.74 ± 0.11 a |
| BS + Mi | 24.94 ± 1.02 ab | 5.09 ± 0.17 bc | 21.74 ± 0.87 bc | 6.28 ± 0.33 a |
| BM + Mi | 25.40 ± 0.69 ab | 4.63 ± 0.18 cd | 24.13 ± 0.95 ab | 6.50 ± 0.30 a |
| TH + Mi | 24.22 ± 1.61 bc | 3.93 ± 0.12 e | 22.18 ± 0.93 bc | 6.05 ± 0.46 a |
| PF + TH + Mi | 25.72 ± 0.26 ab | 5.82 ± 0.49 ab | 24.66 ± 0.58 a | 5.97 ± 0.23 a |
| BS + TH + Mi | 24.33 ± 0.71 bc | 6.21 ± 0.36 a | 25.88 ± 0.74 a | 6.69 ± 0.16 a |
| BM + TH + Mi | 27.58 ± 1.15 a | 5.81 ± 0.22 ab | 26.03 ± 1.06 a | 6.64 ± 0.22 a |
| (a) | |||||||||
| Sample Name | Limonene | Linalool | Methyl Chavicol | Eugenol | Methyl Eugenol | Caryophyllene | Citral 1 | Geraniol | Citral 2 |
| CL | 0.38 ± 0.003 a | 23.81 ± 0.32 cd | 66.59 ± 0.21 c | 0.18 ± 0.005 e | 0.20 ± 0.003 f | 0.77 ± 0.005 d | 0.77 ± 0.006 e | 0.18 ± 0.003 e | 0.94 ± 0.003 d |
| CL + Mi | 0.23 ± 0.03 d | 22.70 ± 0.28 e | 50.16 ± 0.23 h | 1.34 ± 0.006 a | 0.03 ± 0.003 i | 0.78 ± 0.01 d | 1.09 ± 0.006 d | 0.53 ± 0.01 c | 1.36 ± 0.06 b |
| Carbofuran + Mi | 0.23 ± 0.003 d | 22.86 ± 0.03 e | 60.52 ± 0.03 g | 0.22 ± 0.004 d | 0.02 ± 0.002 i | 0.34 ± 0.002 h | 2.13 ± 0.015 a | 1.13 ± 0.04 a | 1.47 ± 0.004 a |
| PF + Mi | 0.27 ± 0.03 c | 23.36 ± 0.11 de | 63.68 ± 0.26 e | 0.23 ± 0.005 d | 0.31 ± 0.006 d | 0.63 ± 0.009 e | 1.54 ± 0.06 b | 0.71 ± 0.003 b | 1.17 ± 0.003 c |
| BS + Mi | 0.34 ± 0.005 b | 24.68 ± 0.29 b | 62.24 ± 0.58 f | 0.42 ± 0.008 b | 0.55 ± 0.008 b | 0.85 ± 0.002 b | 1.27 ± 0.005 c | 0.48 ± 0.003 c | 1.52 ± 0.006 a |
| BM + Mi | 0.27 ± 0.008 c | 22.57 ± 0.06 e | 65.34 ± 0.08 d | 0.18 ± 0.006 e | 0.25 ± 0.006 e | 0.46 ± 0.003 g | 2.10 ± 0.01 a | 1.08 ± 0.01 a | 1.18 ± 0.015 c |
| TH + Mi | 0.23 ± 0.01 d | 22.85 ± 0.07 e | 63.52 ± 0.31 e | 0.35 ± 0.008 c | 0.42 ± 0.006 c | 0.82 ± 0.017 c | 1.12 ± 0.11 d | 0.37 ± 0.02 d | 1.20 ± 0.018 c |
| PF + TH + Mi | 0.22 ± 0.005 d | 26.13 ± 0.08 a | 67.22 ± 0.23 c | 0.13 ± 0.007 f | 0.08 ± 0.001 h | 0.62 ± 0.005 e | 0.65 ± 0.008 ef | 0.16 ± 0.01 e | 0.68 ± 0.008 f |
| BS + TH + Mi | 0.24 ± 0.004 d | 24.47 ± 0.06 bc | 69.11 ± 0.06 b | 0.12 ± 0.004 f | 0.14 ± 0.01 g | 0.57 ± 0.008 f | 0.63 ± 0.01 f | 0.06 ± 0.003 f | 0.78 ± 0.05 e |
| BM + TH + Mi | 0.22 ± 0.01 d | 14.72 ± 0.01 f | 78.14 ± 0.02 a | 0.12 ± 0.01 f | 0.62 ± 0.005 a | 0.94 ± 0.01 a | 0.26 ± 0.02 g | 0.13 ± 0.03 ef | 0.35 ± 0.003 g |
| (b) | |||||||||
| Sample name | Limonene | Linalool | MethylChavicol | Eugenol | MethylEugenol | Caryophyllene | Citral1 | Geraniol | Citral2 |
| CL | 0.2 ± 0.008 a | 25.48 ± 0.35 b | 66.42 ± 0.12 d | 0.17 ± 0.006 c | 0.23 ± 0.01 bc | 0.54 ± 0.02 h | 0.85 ± 0.02 e | 0.37 ± 0.006 d | 0.66 ± 0.01 e |
| CL + Mi | 0.24 ± 0.003 cde | 22.53 ± 0.45 d | 46.86 ± 0.22 g | 0.65 ± 0.005 a | 1.15 ± 0.005 a | 0.96 ± 0.01 c | 1.42 ± 0.01 c | 1.12 ± 0.07 a | 1.79 ± 0.07 a |
| Carbofuran + Mi | 0.26 ± 0.006 abc | 24.84 ± 0.24 b | 64.14 ± 0.41 f | 0.12 ± 0.04 de | 0.06 ± 0.006 de | 0.83 ± 0.006 d | 1.32 ± 0.001 d | 0.66 ± 0.02 c | 0.85 ± 0.02 c |
| PF + Mi | 0.26 ± 0.002 ab | 23.35 ± 0.12 c | 63.70 ± 0.31 f | 0.24 ± 0.003 b | 0.31 ± 0.006 b | 0.61 ± 0.01 g | 1.55 ± 0.004 b | 0.71 ± 0.003 c | 1.15 ± 0.01 b |
| BS + Mi | 0.24 ± 0.008 de | 23.31 ± 0.14 c | 69.30 ± 0.23 b | 0.14 ± 0.008 d | 0.08 ± 0.005 cde | 0.75 ± 0.002 e | 0.52 ± 0.01 h | 0.22 ± 0.01 e | 0.52 ± 0.02 f |
| BM + Mi | 0.23 ± 0.004 de | 25.23 ± 0.15 b | 65.31 ± 0.18 e | 0.13 ± 0.007 de | 0.03 ± 0.004 e | 1.46 ± 0.008 a | 0.77 ± 0.007 f | 0.17 ± 0.003 ef | 0.86 ± 0.006 c |
| TH + Mi | 0.23 ± 0.003 e | 25.15 ± 0.07 b | 66.24 ± 0.02 d | 0.12 ± 0.004 de | 0.20 ± 0.001 bcd | 1.32 ± 0.02 b | 0.70 ± 0.03 g | 0.14 ± 0.007 f | 0.84 ± 0.01 c |
| PF + TH + Mi | 0.25 ± 0.004 bcd | 27.03 ± 0.01 a | 66.44 ± 0.35 d | 0.07 ± 0.002 f | 0.05 ± 0.0001 e | 0.84 ± 0.005 d | 0.70 ± 0.003 g | 0.11 ± 0.008 f | 0.87 ± 0.01 c |
| BS + TH + Mi | 0.23 ± 0.004 e | 23.36 ± 0.21 c | 67.88 ± 0.36 c | 0.11 ± 0.003 e | 0.06 ± 0.0004 de | 0.56 ± 0.002 h | 2.38 ± 0.02 a | 0.86 ± 0.03 b | 0.83 ± 0.02 c |
| BM + TH + Mi | 0.25 ± 0.008 bcd | 22.26 ± 0.12 d | 71.16 ± 0.04 a | 0.08 ± 0.005 f | 0.04 ± 0.0006 e | 0.67 ± 0.006 f | 0.77 ± 0.008 f | 0.23 ± 0.01 e | 0.74 ± 0.005 d |
| Treatments | Sterilized Soil | Natural Soil | ||||
|---|---|---|---|---|---|---|
| N% | P% | K% | N% | P% | K% | |
| CL | 0.71 ± 0.02 de | 0.37 ± 0.005 bc | 0.89 ± 0.01 c | 0.91 ± 0.01 d | 0.64 ± 0.02 a | 1.28 ± 0.02 b |
| CL + Mi | 0.64 ± 0.01 de | 0.32 ± 0.004 c | 0.90 ± 0.04 c | 0.97 ± 0.008 d | 0.36 ± 0.01 f | 0.83 ± 0.13 d |
| Carbofuran + Mi | 0.86 ± 0.02 d | 0.37 ± 0.007 bc | 0.93 ± 0.02 c | 1.22 ± 0.01 bc | 0.44 ± 0.006 bc | 1.15 ± 0.07 bc |
| PF + Mi | 0.54 ± 0.03 e | 0.32 ± 0.005 c | 0.94 ± 0.01 c | 1.19 ± 0.01 c | 0.45 ± 0.04 b | 1.18 ± 0.05 bc |
| BS + Mi | 1.57 ± 0.19 b | 0.50 ± 0.007 a | 0.92 ± 0.02 c | 1.32 ± 0.03 b | 0.42 ± 0.006 bcd | 1.53 ± 0.04 a |
| BM + Mi | 0.73 ± 0.02 de | 0.42 ± 0.015 ab | 1.12 ± 0.06 b | 1.24 ± 0.05 bc | 0.41 ± 0.004 cde | 0.98 ± 0.04 cd |
| TH + Mi | 1.45 ± 0.04 b | 0.33 ± 0.02 bc | 0.95 ± 0.03 c | 1.29 ± 0.04 b | 0.39 ± 0.001 def | 1.24 ± 0.02 b |
| PF + TH + Mi | 1.90 ± 0.01 a | 0.37 ± 0.07 bc | 1.29 ± 0.03 a | 1.31 ± 0.05 b | 0.38 ± 0.006 ef | 1.26 ± 0.04 b |
| BS + TH + Mi | 1.52 ± 0.12 b | 0.41 ± 0.01 bc | 1.32 ± 0.06 a | 1.69 ± 0.04 a | 0.43 ± 0.001 bc | 1.36 ± 0.01 ab |
| BM + TH + Mi | 1.20 ± 0.04 c | 0.42 ± 0.02 ab | 1.31 ± 0.07 a | 1.21 ± 0.009 bc | 0.43 ± 0.001 bc | 1.31 ± 0.02 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiwari, S.; Pandey, R.; Gross, A. Identification of Rhizospheric Microorganisms That Manages Root Knot Nematode and Improve Oil Yield in Sweet Basil (Ocimum basilicum L.). Agronomy 2021, 11, 570. https://doi.org/10.3390/agronomy11030570
Tiwari S, Pandey R, Gross A. Identification of Rhizospheric Microorganisms That Manages Root Knot Nematode and Improve Oil Yield in Sweet Basil (Ocimum basilicum L.). Agronomy. 2021; 11(3):570. https://doi.org/10.3390/agronomy11030570
Chicago/Turabian StyleTiwari, Sudeep, Rakesh Pandey, and Avner Gross. 2021. "Identification of Rhizospheric Microorganisms That Manages Root Knot Nematode and Improve Oil Yield in Sweet Basil (Ocimum basilicum L.)" Agronomy 11, no. 3: 570. https://doi.org/10.3390/agronomy11030570
APA StyleTiwari, S., Pandey, R., & Gross, A. (2021). Identification of Rhizospheric Microorganisms That Manages Root Knot Nematode and Improve Oil Yield in Sweet Basil (Ocimum basilicum L.). Agronomy, 11(3), 570. https://doi.org/10.3390/agronomy11030570