
Effects of Cultivar, Nitrogen Rate and Harvest Time on the Content of Carbohydrates and Protein in the Biomass of Perennial Ryegrass


Abstract
1. Introduction
2. Materials and Methods
2.1. Field Experiment
- P—total monthly precipitation,
- ∑t—mean monthly temperature (sum of mean daily temperatures) > 0.
2.2. Plant Materials
2.3. Statistical Analysis
3. Results and Discussion
3.1. Weather Conditions
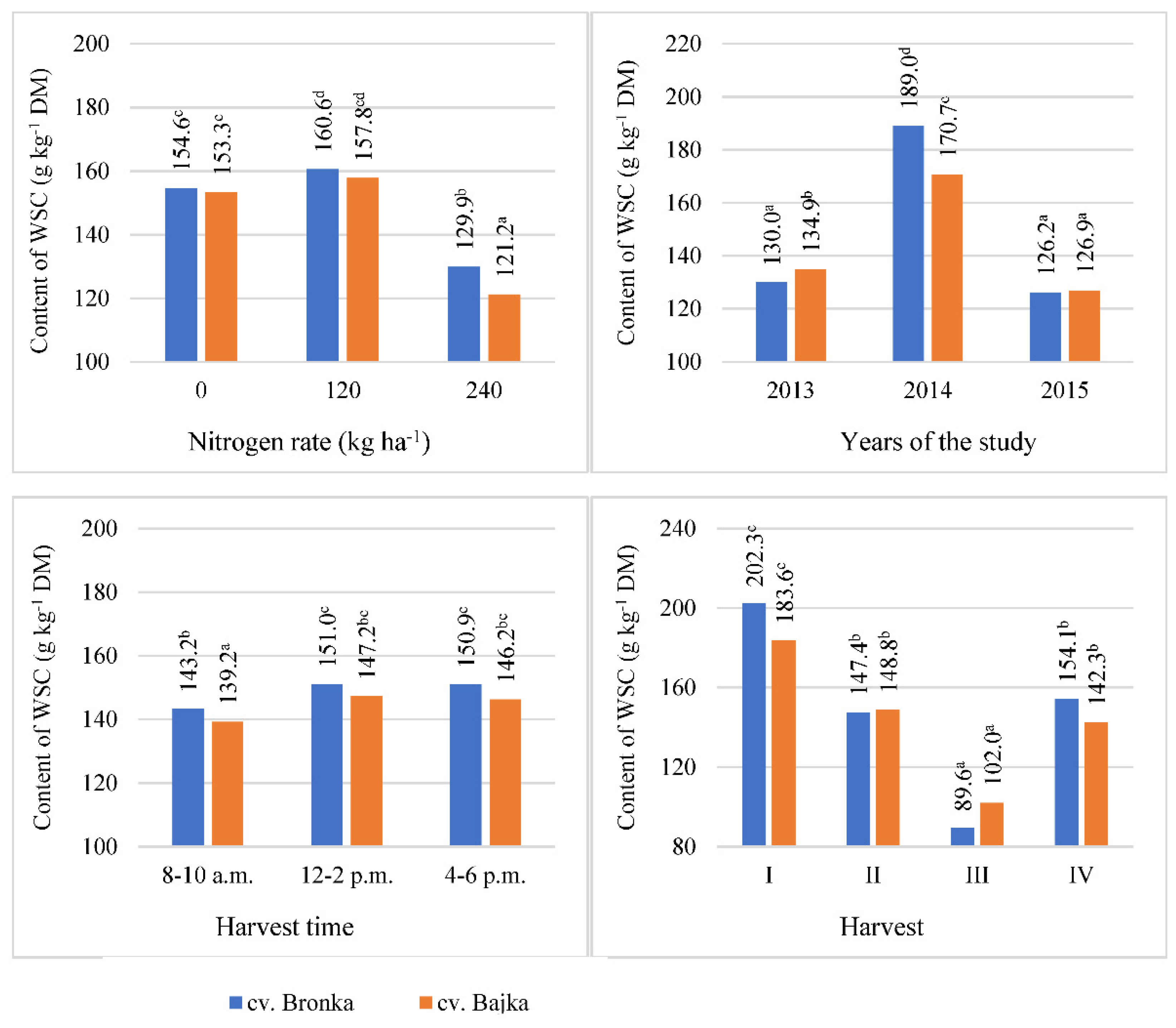
3.2. Water-Soluble Carbohydrates (WSC)
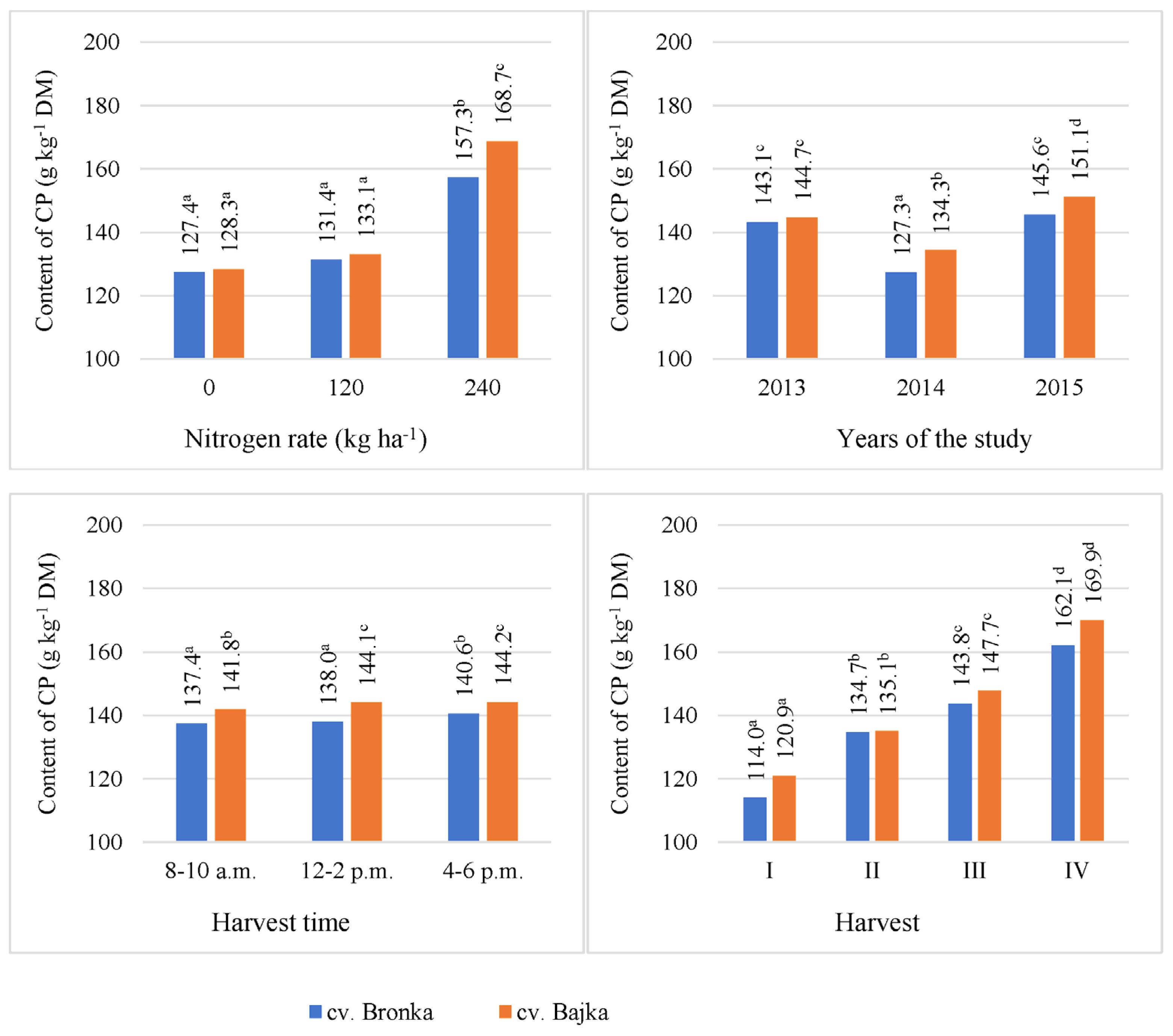
3.3. Crude Protein (CP)
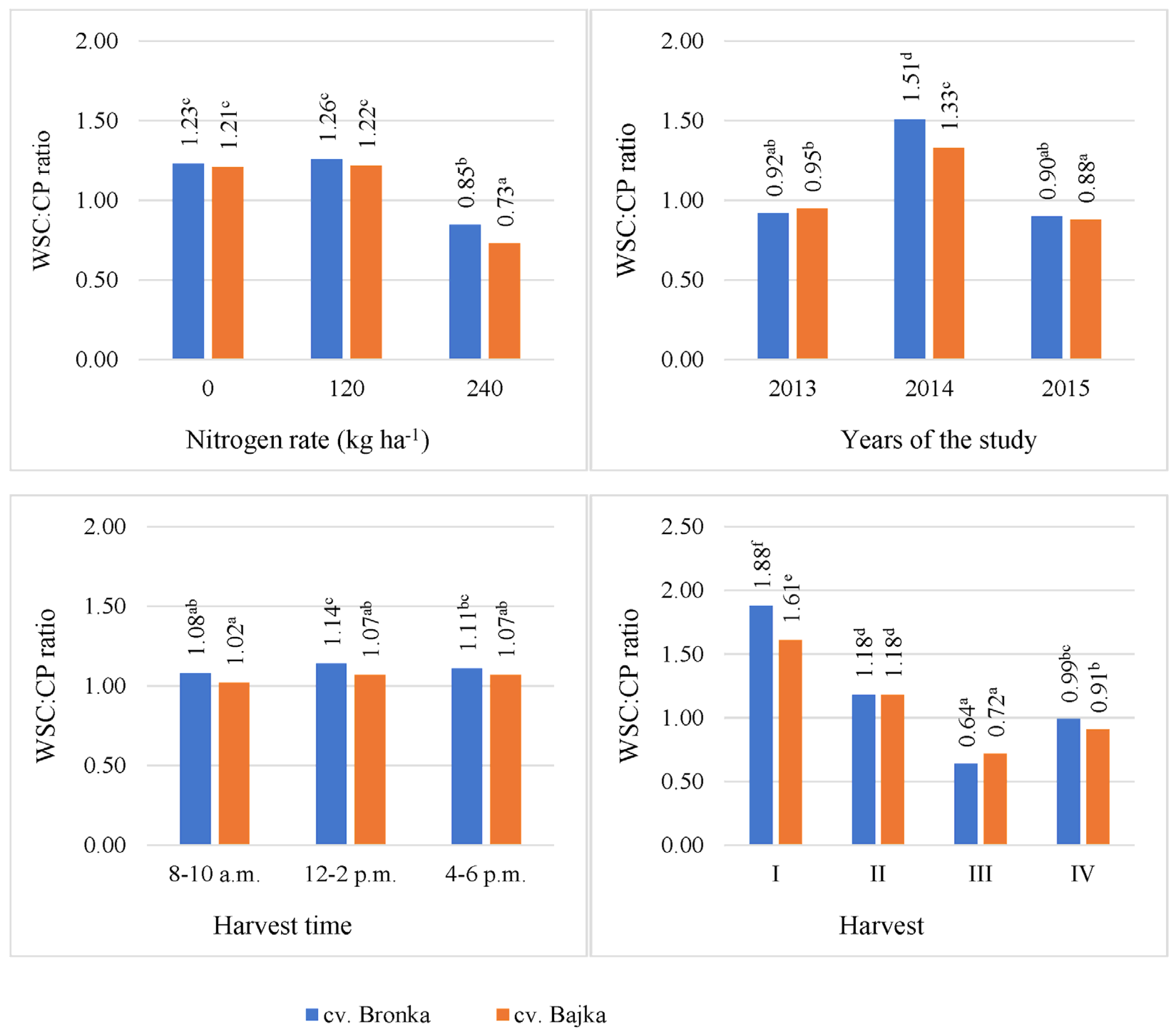
3.4. Carbohydrate-To-Protein Ratio
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brzóska, F.; Śliwiński, B. Quality of roughages in ruminants nutrition and methods for its evaluation. Part I. Characteristics of roughages and measures of their quality. Wiad. Zoot. 2011, 2, 11–23. (In Polish) [Google Scholar]
- Balocchi, O.A.; López, I.F. Herbage production, nutritive value and grazing preference of diploid and tetraploid perennial ryegrass cultivars (Lolium perenne L.). Chil. J. Agric. Res. 2009, 69, 331–339. [Google Scholar] [CrossRef]
- Sandrin, C.Z.; Domingos, M.; Figueiredo-Ribeiro, R.D.C.L. Partitioning of water soluble carbohydrates in vegetative tissues of Lolium multiflorum Lam. ssp. italicum cv. Lema. Braz. J. Plant Physiol. 2006, 18, 299–305. [Google Scholar] [CrossRef]
- Turner, L.R.; Holloway-Philips, M.M.; Rawnsley, R.P.; Donaghy, D.J.; Pembleton, K.G. The morphological and physiological responses of perennial ryegrass (Lolium perenne L.), cocksfoot (Dactylis glomerata L.) and tall fescue (Festuca arundinacea Schreb.; syn. Schedonorus phoenix Scop.) to variable water availability. Grass Forage Sci. 2012, 67, 507–518. [Google Scholar] [CrossRef]
- Jones, E.L.; Roberts, J.E. A note on the relationship between palatability and water-soluble carbohydrates content in perennial ryegrass. Irish J. Agric. Food Res. 1991, 30, 163–167. [Google Scholar]
- Stewart, A.; Hayes, R. Ryegrass breeding-balancing trait priorities. Irish J. Agric. Food Res. 2011, 50, 31–46. [Google Scholar]
- Gilliland, T.J.; Barrett, P.D.; Mann, R.L.; Agnew, R.E.; Fearon, A.M. Canopy morphology and nutritional quality traits as potential grazing value indicators for Lolium perenne varieties. J. Agric. Sci. 2002, 139, 257–273. [Google Scholar] [CrossRef]
- Turner, L.R.; Donaghy, D.J.; Pembleton, K.G.; Rawnsley, R.P. Longer defoliation interval ensures expression of the ‘high sugar’ trait in perennial ryegrass cultivars in cool temperate Tasmania, Australia. J. Agric. Sci. 2015, 153, 995–1005. [Google Scholar] [CrossRef]
- Jafari, A.A.; Rezaeifard, M. Effects of maturity on yield and quality traits in tall fescue (Festuca arundinace Schreb). Am. Eurasian J. Agric. Environ. Sci. 2010, 9, 98–104. [Google Scholar]
- Rezaeifard, M.; Jafari, A.A.; Assareh, M.H. Effects of phenological stages on forage yield quality traits in cocksfoot. J. Food Agric. Environ. 2010, 8, 365–369. [Google Scholar]
- Gilliland, T.; Johnson, J.; Connolly, C. A review of forage grass and clover seed use in Northern Ireland, UK, between 1980 and 2004. Grass Forage Sci. 2007, 62, 239–254. [Google Scholar] [CrossRef]
- Smith, K.F.; Simpson, R.J.; Culvenor, R.A.; Humphreys, M.O.; Prud’ Homme, P.; Oram, R.N. The effects of ploidy and a phenotype conferring a high water-soluble carbohydrate concentration on carbohydrate accumulation, nutritive value and morphology of perennial ryegrass (Lolium perenne L.). J. Agric. Sci. 2001, 136, 65–74. [Google Scholar] [CrossRef]
- Dijkstra, J.; Oenema, O.; Bannink, A. Dietary strategies to reducing N excretion from cattle: Implications formethane emissions. Curr. Opin. Environ. Sustain. 2011, 3, 414–422. [Google Scholar] [CrossRef]
- Evans, J.G.; Fraser, M.D.; Owen, I.; Davies, D.A. An evaluation of two perennial ryegrass cultivars (AberDartand and Fennema) for sheep production in the uplands. J. Agric. Sci. 2011, 149, 235–248. [Google Scholar] [CrossRef]
- Cosgrove, G.P.; Burke, J.L.; Death, A.F.; Hickey, M.J.; Pacheco, D.; Fraser, K.; Lane, G.A. Ryegrass with increased water soluble carbohydrate: Evaluating the potential for grazing dairy cows in New Zealand. Proc. N. Z. Grassl. Assoc. 2007, 69, 179–185. [Google Scholar]
- Tas, B.M.; Taweel, H.Z.; Smit, H.J.; Elgersma, A.; Dijkstra, J.; Tamminga, S. Effects of perennial ryegrass cultivars on milk yield and nitrogen utilization in grazing dairy cows. J. Dairy Sci. 2006, 89, 3494–3500. [Google Scholar] [CrossRef]
- Miller, L.A.; Moorby, J.M.; Davies, D.R.; Humphreys, M.O.; Scollan, N.D.; Macrae, J.C.; Theodorou, M.K. Increased concentration of water-soluble carbohydrate in perennial ryegrass (Lolium perenne L.). Milk production from late-lactation dairy cows. Grass Forage Sci. 2001, 56, 383–394. [Google Scholar] [CrossRef]
- Lee, M.R.F.; Jones, E.L.; Moorby, J.M.; Humphreys, M.O.; Theodorou, M.K.; MacRae, J.C.; Scollan, N.D. Production responses from lambs grazed on Lolium perenne selected for an elevated water-soluble carbohydrate concentration. Anim. Res. 2001, 50, 441–449. [Google Scholar] [CrossRef]
- Orr, R.J.; Rutter, S.M.; Penning, P.D.; Rook, A.J. Matching grass supply to grazing pattern for dairy cows. Grass Forage Sci. 2001, 56, 352–361. [Google Scholar] [CrossRef]
- Baert, J.; van Waes, C. Improvement of the digestibility of tall fescue (Festuca arundinacea Schreb.) inspired by perennial ryegrass (Lolium perenne L.). Grassl. Sci. Eur. 2014, 19, 172–174. [Google Scholar]
- Andrzejewska, J.; Albrecht, K.A.; Jędrzejczak, E. Plant height and feed value of alfalfa in different development stages and cuts. Fragm. Agron. 2013, 30, 14–22. (In Polish) [Google Scholar]
- Parsons, A.J.; Edwards, G.R.; Newton, P.C.D.; Chapman, D.F.; Caradus, J.R.; Rasmussen, S.; Rowarth, J.S. Past lessons and future prospects: Plant breeding for yield and persistence in cool-temperate pastures. Grass Forage Sci. 2011, 66, 153–172. [Google Scholar] [CrossRef]
- Edwards, G.R.; Parsons, A.J.; Rasmussen, S.; Bryant, R.H. High sugar ryegrasses for livestock systems in New Zealand. Proc. N. Z. Grassl. Assoc. 2007, 69, 161–171. [Google Scholar] [CrossRef]
- Wilkins, P.W.; Humphreys, M.O. Progress in breeding perennial forage grasses for temperate agriculture. J. Agric. Sci. 2003, 140, 129–150. [Google Scholar] [CrossRef]
- Valk, H.; Poelhuis, H.W.K.; Wentink, H.J. Effect of fibrous and starchy carbohydrates in concentrates as supplements in a herbage-based diet for high-yielding dairy cows. Neth. J. Agric. Sci. 1990, 38, 475–486. [Google Scholar] [CrossRef]
- Loaiza, P.; Balocchi, O.; López, I.F. Changes in water-soluble carbohydrates relative to crude protein in perennial ryegrass in response to defoliation frequency. Grassland Sci. 2017, 63, 159–168. [Google Scholar] [CrossRef]
- Da Silva, M.S.; Tremblay, G.F.; Bélanger, G.; Lajeunesse, J.; Papadopoulos, Y.A.; Fillmore, S.A.; Jobim, C.C. Forage energy to protein ratio of several legume–grass complex mixtures. Anim. Feed Sci. Technol. 2014, 188, 17–27. [Google Scholar] [CrossRef]
- Huhtanen, P.; Broderick, G. Improving utilisation of forage protein in ruminant production by crop and feed management. Grass Forage Sci. 2016, 61, 340–349. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources, 2014, Update 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015; p. 203. [Google Scholar]
- Skowera, B.; Puła, J. Pluviometric extreme conditions in spring season in Poland in the years 1971–2000. Acta Agroph. 2004, 3, 171–177. (In Polish) [Google Scholar]
- Thomas, T.A. An automated procedure for the determination of soluble carbohydrates in herbage. J. Sci. Food Agric. 1977, 28, 639–642. [Google Scholar] [CrossRef]
- Houba, V.J.G.; Van der Lee, J.J.; Novozamsky, I. Soil and Plant Analysis; Part 5B. Soil Analysis Procedure other Procedure; Wageningen Agriculture University: Wageningen, The Netherlands, 1995. [Google Scholar]
- TIBCO Software Inc. Statistica (Data Analysis Software System, Palo Alto, USA). 2017. Version 13.3. Available online: https://docs.tibco.com/products/tibco-statistica-13-3-0 (accessed on 15 February 2021).
- Blum, A.; Tuberosa, R. Dehydration survival of crop plants and its measurement. J. Exp. Bot. 2018, 69, 975–981. [Google Scholar] [CrossRef]
- Da Costa, M.; Huang, B. Osmotic adjustment associated with variation in bentgrass tolerance to drought stress. J. Am. Soc. Hortic. Sci. 2006, 131, 338–344. [Google Scholar] [CrossRef]
- Starck, Z. Plant responses to the abiotic environmental stresses—Aclimatization and adaptation. Łąk. Pol. 2005, 8, 173–184. (In Polish) [Google Scholar]
- Meisser, M.; Vitra, A.; Mosimann, E.; Deléglise, C.; Buttler, A. Linking functional plant traits and forage quality under drought conditions. Grassl. Sci. Eur. 2017, 22, 366–368. [Google Scholar]
- Nazarli, H.; Faraji, F. Response of proline, soluble sugars and antioxidant enzymes in wheat to different irrigation regimes in greenhouse condition. Cercet. Agron. Mold. 2011, 4, 27–33. [Google Scholar] [CrossRef]
- Weinberg, Z.G.; Bar-Tal, A.; Chen, Y.; Gamburg, M.; Brener, S.; Dvash, L.; Markovitz, T.; Landau, S. The effects of irrigation and nitrogen fertilization on the ensiling of safflower (Carthamus tinctorius). Anim. Feed Sci. Technol. 2007, 134, 152–161. [Google Scholar] [CrossRef]
- Hume, D.E.; Hickey, M.J.; Lyons, T.B.; Baird, D.B. Agronomic performance and water-soluble carbohydrate expression of selected ryegrasses at two locations in New Zealand. N. Z. J. Agric. Res. 2010, 53, 37–57. [Google Scholar] [CrossRef][Green Version]
- Chesney, L.; Scollan, N.; Gordon, A.; McConnell, D.A.; Lively, F.O. Investigating the effect of nitrogen application level on grass production and quality under simulated grazing. Grassl. Sci. Eur. 2020, 25, 55–57. [Google Scholar]
- Conaghan, P.; O’Kiely, P.; Halling, M.A.; O’Mara, F.P.; Nesheim, L. Yield and quality response of perennial ryegrass selected for high concentration of water-soluble carbohydrate to nitrogen application rate. Crop. Sci. 2012, 52, 28–39. [Google Scholar] [CrossRef]
- Lattanzi, F.A.; Ostler, U.; Wild, M.; Morvan-Bertrand, A.; Decau, M.L.; Lehmeier, C.A.; Meuriot, F.; Proud’homme, M.P.; Schaufele, R.; Schnyder, H. Flexes in central carbohydrate metabolism of source leaves in a fructan—Storing C3 grass: Rapid turnover and futile cycling of sucrose in continuous light under contrasted nitrogen nutrition status. J. Exp. Bot. 2012, 63, 2363–2375. [Google Scholar] [CrossRef]
- Ciepiela, G.A. Content of structural and nonstructural carbohydrates and lignin in Dactylis glomerata L. and Festulolium braunii (K. Richt.) A. Camus supplied by biostimulator Kelpak SL and nitrogen. Nauka Przyr. Technol. 2014, 8, 1–12. (In Polish) [Google Scholar]
- Ciepiela, G.A.; Jankowski, K.; Jodełka, J. Effect of various nitrogen fertilisation forms on perennial ryegrass (Lolium perenne L.) yielding and content of carbohydrates and some nitrogen forms. Acta Sci. Pol. Agric. 2003, 2, 131–139. (In Polish) [Google Scholar]
- Lee, J.M.; Donaghy, D.J.; Roche, J.R. The effect of grazing severity and fertiliser application during winter on herbage regrowth and quality of perennial ryegrass (Lolium perenne L.). Aust. J. Exp. Agric. 2007, 47, 825–832. [Google Scholar] [CrossRef]
- Bryant, R.H.; Gregorini, P.; Edwards, G.R. Effects of N fertilisation, leaf appearance and time of day on N fractionation and chemical composition of Lolium perenne cultivars in spring. Anim. Feed Sci. Technol. 2012, 173, 210–219. [Google Scholar] [CrossRef]
- Griggs, T.C.; MacAdam, J.W.; Mayland, H.F.; Burns, J.C. Nonstructural carbohydrate and digestibility patterns in temperate pastures and canopy management implications. In Proceedings of the AFGC Annual Conference, French Lick, IN, USA, 12–15 June 2011. [Google Scholar]
- Trevaskis, L.M.; Fulkerson, W.J.; Nandra, K.S. Effect of time of feeding carbohydrate supplements and pasture on production of dairy cows. Liv. Prod. Sci. 2004, 85, 275–285. [Google Scholar] [CrossRef]
- Truba, M.; Jankowski, K.; Wiśniewska-Kadżajan, B.; Malinowska, E. The effect of soil conditioners on the content of soluble carbohydrates, digestible protein and the carbohydrate–protein ratio in Lolium perenne and Dactylis glomerata. Environ. Prot. Natur. Res. 2018, 29, 1–8. [Google Scholar]
- Purwin, C.; Stanek, M.; Lipiński, K.; Wierzbowska, J.; Nogalska, A.; Fijalkowska, M. Effect of a harvest time and cultivar on the chemical composition and in vitro ruminal dry matter degradability of perennial ryegrass (Lolium perenne L.). J. Elem. 2016, 21, 811–822. [Google Scholar]
- Lessire, F.; Hornick, J.L.; Dufrasne, I. Effects of concentrate levels on milk production and traffic of cows milked by a mobile automatic milking system on pasture. Grassl. Sci. Eur. 2014, 19, 545–549. [Google Scholar]
- Delagarde, R.; Peyraud, J.L.; Delaby, L.; Faverdin, P. Vertical distribution of biomass, chemical composition and pepsin-cellulase digestibility in a perennial ryegrass sward: Interaction with month of year, regrowth age and time of day. Anim. Feed Sci. Technol. 2000, 84, 49–68. [Google Scholar] [CrossRef]
- Pirhofer-Walzl, K.; Søegaard, K.; Høgh-Jensen, H.; Eriksen, J.; Sanderson, M.A.; Rasmussen, J.; Rasmussen, J. Forage herbs improve mineral composition of grassland herbage. Grass Forage Sci. 2011, 66, 415–423. [Google Scholar] [CrossRef]
- Schlegel, P.; Wyss, U.; Arrigo, Y.; Hess, H.D. Mineral concentrations of fresh herbage from mixed grassland as influenced by botanical composition, harvest time and growth stage. Anim. Feed Sci. Tech. 2016, 219, 226–233. [Google Scholar] [CrossRef]
- Da Silva, M.S.; Tremblay, G.F.; Bélanger, G.; Lajeunesse, J.; Papadopoulos, Y.A.; Fillmore, S.A.E.; Jobim, C.C. Energy to protein ratio of grass-legume binary mixtures under frequent clipping. Agron. J. 2013, 105, 482–492. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Years of the Study | Month | ||||||
|---|---|---|---|---|---|---|---|
| April | May | June | July | August | September | November | |
| 2013 | 1.6 (o) | 1.2 (fd) | 1.2 (fd) | 2.2 (w) | 0.7 (d) | 3.0 (vw) | 0.57 (vd) |
| 2014 | 1.0 (fd) | 0.8 (d) | 1.2 (fd) | 0.6 (vd) | 1.6 (o) | 0.6 (vd) | 0.55 (vd) |
| 2015 | 1.9 (fw) | 0.8 (d) | 0.6 (vd) | 1.5 (o) | 0.2 (ed) | 1.6 (o) | 1.02 (fd) |
| Parameter | Years of the Study | Mean | ||
|---|---|---|---|---|
| 2013 | 2014 | 2015 | ||
| Cultivar | ||||
| Baronka | 129.8 a | 189.0 b | 126.1 a | 148.3 b |
| Bajka | 134.9 b | 170.7 a | 126.9 a | 144.2 a |
| Nitrogen rate (kg ha−1) | ||||
| 0 | 139.5 b | 177.2 b | 147.4 c | 154.1 b |
| 120 | 143.5 b | 200.7 c | 132.7 b | 159.2 c |
| 240 | 114.1 a | 161.8 a | 100.8 a | 125.5 a |
| Harvest time | ||||
| 8.00–10.00 a.m. | 117.7 a | 177.9 a | 127.4 a | 141.2 a |
| 12.00–2.00 p.m. | 141.4 b | 181.3 a | 124.4 a | 149.1 b |
| 4.00–6.00 p.m. | 138.0 b | 180.4 a | 127.8 a | 148.5 b |
| Harvest | ||||
| I | 122.3 b | 256.1 c | 200.4 d | 192.9 c |
| II | 147.5 c | 185.2 b | 111.6 c | 148.1 b |
| III | 102.5 a | 96.3 a | 88.6 a | 95.8 a |
| IV | 157.3 d | 181.9 b | 105.3 b | 148.2 b |
| Parameter | Years of the Study | Mean | ||
|---|---|---|---|---|
| 2013 | 2014 | 2015 | ||
| Cultivar | ||||
| Baronka | 143.1 a | 127.2 a | 145.6 a | 138.7 a |
| Bajka | 144.7 a | 134.2 b | 151.1 b | 143.4 b |
| Nitrogen rate (kg ha−1) | ||||
| 0 | 137.4 a | 122.3 b | 123.8 a | 127.8 a |
| 120 | 135.5 a | 116.6 a | 144.6 b | 132.2 b |
| 240 | 158.8 b | 153.4 c | 176.7 c | 163.0 c |
| Harvest time | ||||
| 8.00–10.00 a.m. | 143.8 a | 130.0 ab | 144.9 a | 139.6 a |
| 12.00–2.00 p.m. | 144.6 a | 128.8 a | 149.9 b | 141.1 ab |
| 4.00–6.00 p.m. | 143.4 a | 133.3 b | 150.4 b | 142.4 b |
| Harvest | ||||
| I | 132.0 a | 108.6 a | 111.7 a | 117.4 a |
| II | 134.7 a | 118.9 b | 151.1 b | 134.9 b |
| III | 145.6 b | 141.8 c | 149.8 b | 145.7 c |
| IV | 163.3 c | 153.7 d | 181.0 c | 166.0 d |
| Parameter | Years of the Study | Mean | ||
|---|---|---|---|---|
| 2013 | 2014 | 2015 | ||
| Cultivar | ||||
| Baronka | 0.92 a | 1.5 b | 0.90 a | 1.11 b |
| Bajka | 0.95 a | 1.3 a | 0.88 a | 1.05 a |
| Nitrogen rate | ||||
| 0 | 1.01 b | 1.45 b | 1.18 c | 1.22 b |
| 120 | 1.07 c | 1.73 c | 0.92 b | 1.24 b |
| 240 | 0.72 a | 1.07 a | 0.57 a | 0.79 c |
| Harvest time | ||||
| 8.00–10.00 a.m. | 0.83 a | 1.40 a | 0.91 a | 1.05 a |
| 12.00–2.00 p.m. | 0.99 b | 1.45 a | 0.87 a | 1.09 b |
| 4.00–6.00 p.m. | 0.98 b | 1.40 a | 0.90 a | 1.10 b |
| Harvest | ||||
| I | 0.95 b | 2.39 d | 1.89 c | 1.74 d |
| II | 1.12 c | 1.61 c | 0.81 b | 1.18 c |
| III | 0.72 a | 0.70 a | 0.62 a | 0.68 a |
| IV | 0.99 b | 1.26 b | 0.61 a | 0.95 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olszewska, M. Effects of Cultivar, Nitrogen Rate and Harvest Time on the Content of Carbohydrates and Protein in the Biomass of Perennial Ryegrass. Agronomy 2021, 11, 468. https://doi.org/10.3390/agronomy11030468
Olszewska M. Effects of Cultivar, Nitrogen Rate and Harvest Time on the Content of Carbohydrates and Protein in the Biomass of Perennial Ryegrass. Agronomy. 2021; 11(3):468. https://doi.org/10.3390/agronomy11030468
Chicago/Turabian StyleOlszewska, Marzenna. 2021. "Effects of Cultivar, Nitrogen Rate and Harvest Time on the Content of Carbohydrates and Protein in the Biomass of Perennial Ryegrass" Agronomy 11, no. 3: 468. https://doi.org/10.3390/agronomy11030468
APA StyleOlszewska, M. (2021). Effects of Cultivar, Nitrogen Rate and Harvest Time on the Content of Carbohydrates and Protein in the Biomass of Perennial Ryegrass. Agronomy, 11(3), 468. https://doi.org/10.3390/agronomy11030468