
Potential for the Postharvest Biological Control of Phthorimaea operculella (Lepidoptera, Gelechiidae) by Blattisocius tarsalis (Mesostigmata, Blattisociidae)

 , ,
, ,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Materials and Experimental Conditions
2.2. Evaluation of the Predatory Behaviour at Different Temperatures
2.3. Efficacy Bioassays in Microcosm
3. Results
3.1. Evaluation of the Predatory Activity at Different Temperatures
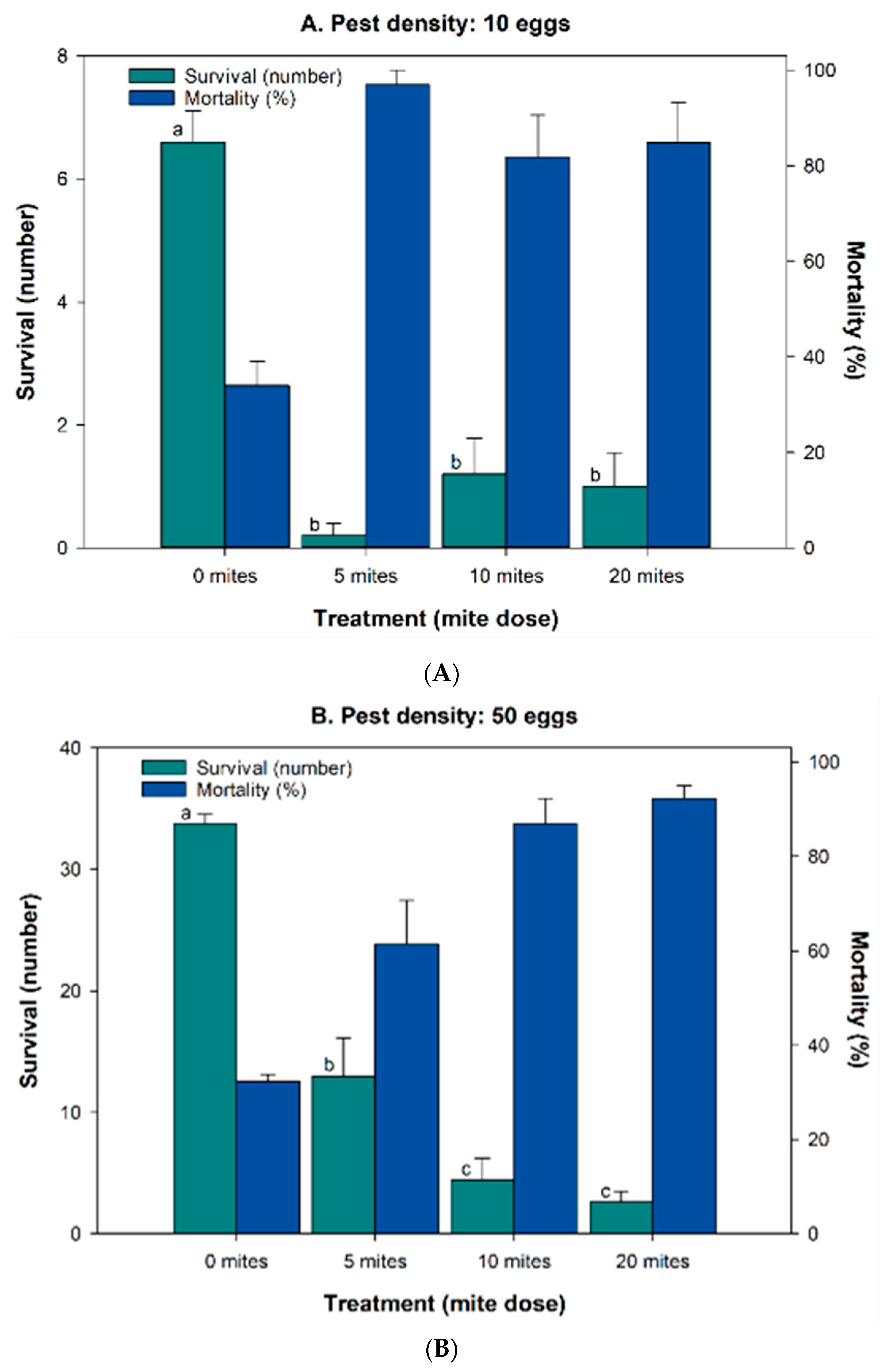
3.2. Efficacy Bioassays in Microcosm
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. FAO Statistics Division 2019. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 15 November 2019).
- Burgos, G.; Zum Felde, T.; Andre, C.; Kubow, S. The Potato and Its Contribution to the Human Diet and Health. In The Potato Crop. Its Agricultural, Nutritional and Social Contribution to Humankind; Campos, H., Ortiz, O., Eds.; Springer: Cham, Switzerland, 2020; pp. 37–74. [Google Scholar]
- Devaux, A.; Goffart, J.P.; Petsakos, A.; Kromann, P.; Gatto, M.; Okello, J.; Hareau, G. Global Food Security, Contributions from Sustainable Potato Agri-Food Systems. In The Potato Crop. Its Agricultural, Nutritional and Social Contribution to Humankind; Campos, H., Ortiz, O., Eds.; Springer: Cham, Switzerland, 2020; pp. 3–35. [Google Scholar]
- Gottschalk, K.; Ezekiel, R. Storage. In Handbook of Potato Production, Improvement, and Postharvest Management; Gopal, J., Kurana, S.M.P., Eds.; Food Products Press: New York, NY, USA, 2006; pp. 489–522. [Google Scholar]
- Fuglie, K.O.; Khatana, V.S.; Ilangantileke, S.G.; Scott, G.J.; Singh, J.; Kumar, D. Economics of potato storage in northern India. Q. J. Int. Agr. 2000, 39, 131–148. [Google Scholar]
- Rondon, S.I. Decoding Phthorimaea operculella (Lepidoptera: Gelechiidae) in the new age of change. J. Integr. Agr. 2020, 19, 316–324. [Google Scholar] [CrossRef]
- Chandel, R.S.; Vashisth, S.; Soni, S.; Kumar, R.; Kumar, V. The Potato Tuber Moth, Phthorimaea operculella (Zeller), in India: Biology, Ecology, and Control. Potato Res. 2020, 63, 15–39. [Google Scholar] [CrossRef]
- Arthurs, S.P.; Lacey, L.A.; Pruneda, J.N.; Rondon, S.I. Semi-field evaluation of a granulovirus and Bacillus thuringiensis ssp. kurstaki for season-long control of the potato tuber moth, Phthorimaea operculella. Entomol. Exp. Appl. 2008, 129, 276–285. [Google Scholar] [CrossRef]
- Lacey, L.A.; Headrick, H.L.; Horton, D.R.; Schreiber, A. Effect of a granulovirus on mortality and dispersal of potato tuber worm (Lepidoptera: Gelechiidae) in refrigerated storage warehouse conditions. Biocontrol. Sci. Technol. 2010, 20, 437–447. [Google Scholar] [CrossRef]
- Kroschel, J.; Mujica, N.; Okonya, J.; Alyokhin, A. Insect Pests Affecting Potatoes in Tropical, Subtropical, and Temperate Regions. In The Potato Crop. Its Agricultural, Nutritional and Social Contribution to Humankind; Campos, H., Ortiz, O., Eds.; Springer: Cham, Switzerland, 2020; pp. 251–306. [Google Scholar]
- Pinhero, R.G.; Coffin, R.; Yada, R.Y. Post-harvest storage of potatoes. In Advances in Potato Chemistry and Technology; Singh, J., Kaur, L., Eds.; Elsevier: London, UK, 2009; pp. 338–370. [Google Scholar] [CrossRef]
- Sporleder, M.; Kroschel, J.; Quispe, M.R.G.; Lagnaoui, A. A temperature-based simulation model for the potato tuberworm, Phthorimaea operculella Zeller (Lepidoptera; Gelechiidae). Environ. Entomol. 2004, 33, 477–486. [Google Scholar] [CrossRef]
- Douches, D.S.; Pett, W.; Santos, F.; Coombs, J.; Grafius, E.; Li, W.; Metry, E.A.; El-Din, T.N.; Madkour, M. Field and storage testing Bt potatoes for resistance to potato tuberworm (Lepidoptera: Gelichiidae). J. Econ. Entomol. 2004, 97, 1425–1431. [Google Scholar] [CrossRef][Green Version]
- Rondon, S.I.; Gao, Y. The Journey of the Potato Tuberworm around the World. In Moths: Pests of Potato, Maize and Sugar Beet; Perveen, K., Ed.; IntechOpen: London, UK, 2018; pp. 17–52. [Google Scholar] [CrossRef]
- Keasar, T.; Steinberg, S. Evaluation of the parasitoid Copidosoma koehleri for biological control of the potato tuber moth, Phthorimaea operculella, in Israeli potato fields. Biocontrol. Sci. Tech. 2008, 18, 325–336. [Google Scholar] [CrossRef]
- Pokharkar, D.S.; Jogi, R.R. Biological suppression of potato tubermoth, Phthorimaea operculella (Zeller) with exotic parasitoids and microbial agents under field and storage conditions. J. Biol. Control. 2000, 14, 23–28. [Google Scholar] [CrossRef]
- Mandour, N.S.; Sarhan, A.A.; Atwa, D.H. The integration between Trichogramma evanescens West.(Hymenoptera: Trichogrammatidae) and selected bioinsecticides for controlling the potato tuber moth Phthorimaea operculella (Zell.)(Lepidoptera: Gelechiidae) of stored potatoes. J. Plant. Protec. Res. 2012, 52, 40–46. [Google Scholar] [CrossRef]
- Gallego, J.R.; Gamez, M.; Cabello, T. Potential of the Blattisocius mali Mite (Acari: Blattisociidae) as a biological control agent of potato tubermoth (Lep.: Gelechiidae) in stored potatoes. Potato Res. 2019, 1–11. [Google Scholar] [CrossRef]
- Gallego, J.R.; Caicedo, O.; Gamez, M.; Hernandez, J.; Cabello, T. Selection of Predatory Mites for the Biological Control of Potato Tuber Moth in Stored Potatoes. Insects 2020, 11, 196. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.Q.; Zalom, F.G.; Nicola, N.L. Laboratory studies of Blattisocius keegani (Acari: Ascidae) reared on eggs of navel orange worm: Potential for biological control. Bull. Entomol. Res. 2011, 11, 499–504. [Google Scholar] [CrossRef]
- Graham, M.M. Warehouse ecology studies of bagged maize in Kenya—II: Ecological observations of an infestation by Ephestia (Cadra) cautella (Walker)(Lepidoptera, Phycitidae). J. Stored Prod. Res. 1970, 6, 157–167. [Google Scholar] [CrossRef]
- Nielsen, P.S. The impact of temperature on activity and consumption rate of moth eggs by Blattisocius tarsalis (Acari: Ascidae). Exp. Appl. Acarol. 1999, 23, 149–157. [Google Scholar] [CrossRef]
- Nielsen, P.S. Developmental Time of Blattisocius tarsalis (Acari: Ascidae) at Different Temperatures. Exp. Appl. Acarol. 2001, 25, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Chant, D.A. The subfamily Blattisocinae Garman (=Aceosejinae Evans) (Acarina: Blattisocidae Garman)(= Aceosejidae Baker and Wharton) in North America, with descriptions of new species. Can. J. Zool. 1963, 41, 243–305. [Google Scholar] [CrossRef]
- Haines, C.P. Laboratory studies on the role of an egg predator, Blattisocius tarsalis (Acari: Ascidae), in relation to the natural control of Ephestia cautella (Lep.: Pyralidae) in warehouses. Bull. Entomol. Res. 1981, 71, 557–574. [Google Scholar] [CrossRef]
- Darst, P.H.; King, E.W. Biology of Melichares tarsalis in association with Plodia interpunctella. Ann. Entomol. Soc. Am. 1969, 62, 747–749. [Google Scholar] [CrossRef]
- Nesbitt, H.J. A taxonomic study of the Phytoseiinae (family Laelaptidae) predaceous upon Tetranychidae of economic importance. Zool. Verh. Leiden 1951, 12, 1–64. [Google Scholar]
- Haines, C.P. A revision of the genus Blattisocius (Mesostigmata: Ascidae) with especial reference to B. tarsalis and the description of a new species. Acarologia 1978, 20, 19–38. [Google Scholar]
- Greenspan, L. Humidity fixed points of binary saturated aqueous solutions. J. Res. Natl. Bur. Stand. 1977, 81, 89–96. [Google Scholar] [CrossRef]
- Fenemore, P.G. Oviposition of potato tuber moth, Phthorimaea operculella (Lep.: Gelechiidae); fecundity in relation to mated state, age, and pupal weight. N. Z. J. Zool. 1977, 4, 18191. [Google Scholar] [CrossRef]
- Healy, M.J.R. A table of Abbott’s correction for natural mortality. Ann. Appl. Biol. 1952, 39, 211–212. [Google Scholar] [CrossRef]
- Al-Ali, A.S.; Al-Neamy, I.K.; Abbas, S.A.; Abdul-Masih, A.M.E. Observations on the biology of the potato tuber moth Phthorimaea operculella Zell. (Lepidoptera, Gelechiidae) in Iraq. Z. Angew. Entomol. 1975, 79, 345–351. [Google Scholar] [CrossRef]
- Rall, B.C.; Vucic-Pestic, O.; Ehnes, R.B.; Emmerson, M.; Brose, U. Temperature, predator–prey interaction strength and population stability. Glob. Chang. Biol. 2010, 16, 2145–2157. [Google Scholar] [CrossRef]
- Kroschel, J.; Schaub, B. Biology and ecology of potato tuber moths as major pests of potato. In Insect Pests of Potato: Global Perspectives on Biology and Management; Giordanengo, P., Vincent, C., Alyokhin, A., Eds.; Elsevier: Waltham, MA, USA, 2013; pp. 165–192. [Google Scholar]
- Richards, O.W.; Thomson, W.S. A contribution to the study of the genera Ephestia, Gn. (including Stymax, Dyar), and Plodia, Gn. (Lepidoptera, Phycitidae) with notes on parasites of the larvae. Trans. R. Ent. Soc. Lond. 1932, 80, 169–250. [Google Scholar] [CrossRef]
- Sabelis, M.W. The functional response of predatory mites to the density of two-spotted spider mites. In Dynamics of Physiologically Structured Populations; Metz, J.A.J., Diekmann, O., Eds.; Springer: Berlin/Heidelberg, Germany, 1986; Volume 68, pp. 298–321. [Google Scholar]
- Riudavets, J.; Maya, M.; Monserrat, M. Predation by Blattisocius tarsalis (Acari: Ascidae) on stored product pests. IOBC WPRS Bull. 2002, 25, 121–126. [Google Scholar]
- Dicke, M.; de Jong, M.; Alers, M.P.T.; Stelder, F.C.T.; Wunderink, R.; Post, J. Quality control of mass-reared arthropods: Nutritional effects on performance of predatory mites. J. Appl. Entomol. 1989, 108, 462–475. [Google Scholar] [CrossRef]
- Flanders, S.; Badgley, M. Prey-predator interactions in self-balanced laboratory populations. Hilgardia 1963, 35, 145–183. [Google Scholar] [CrossRef][Green Version]
- Schausberger, P. Cannibalism among phytoseiid mites: A review. Exp. Appl. Acarol. 2003, 29, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Barker, P.S. Bionomics of Blattisocius keegani (Fox)(Acarina: Ascidae), a predator on eggs of pests of stored grains. Can. J. Zool. 1967, 45, 1093–1099. [Google Scholar] [CrossRef]
- Golizadeh, A.; Zalucki, M.P. Estimating temperature-dependent developmental rates of potato tuberworm, Phthorimaea operculella (Lepidoptera: Gelechiidae). Insect Sci. 2012, 19, 609–620. [Google Scholar] [CrossRef]
- Moraes, J.G.; Venancio, R.; Santos, V.L.V.; Paschoal, D. Potential of Ascidae, Blattisociidae and Melicharidae (Acari: Mesostigmata) as Biological Control Agents of Pest Organisms. In Prospects for Biological Control of Plant Feeding Mites and Other Harmful Organisms; Progress in Biological Control; Carrillo, D., Moraes, G.J., Peña, J.E., Eds.; Springer: Cham, Switzerland, 2015; Volume 19, pp. 33–75. [Google Scholar] [CrossRef]


{kind=link}
| Temperature (°C) | Survivors (Number) 1 | Mortality (%) | Efficacy Percentage (%) | ||
|---|---|---|---|---|---|
| Treatment | Control | Treatment | Control | ||
| 10 | 2.28 ± 0.22 a | 4.48 ± 0.11 b | 55.86 ± 4.14 | 9.66 ± 1.89 | 49.66 ± 5.06 |
| 20 | 0.90 ± 0.20 a | 4.33 ± 0.14 b | 82.00 ± 3.99 | 14.00 ± 2.74 | 78.17 ± 4.77 |
| 30 | 1.03 ± 0.19 a | 4.20 ± 0.14 b | 79.38 ± 3.76 | 16.67 ± 2.63 | 75.57 ± 4.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gavara, J.; Piedra-Buena, A.; Hernandez-Suarez, E.; Gamez, M.; Cabello, T.; Gallego, J.R. Potential for the Postharvest Biological Control of Phthorimaea operculella (Lepidoptera, Gelechiidae) by Blattisocius tarsalis (Mesostigmata, Blattisociidae). Agronomy 2021, 11, 288. https://doi.org/10.3390/agronomy11020288
Gavara J, Piedra-Buena A, Hernandez-Suarez E, Gamez M, Cabello T, Gallego JR. Potential for the Postharvest Biological Control of Phthorimaea operculella (Lepidoptera, Gelechiidae) by Blattisocius tarsalis (Mesostigmata, Blattisociidae). Agronomy. 2021; 11(2):288. https://doi.org/10.3390/agronomy11020288
Chicago/Turabian StyleGavara, Jorge, Ana Piedra-Buena, Estrella Hernandez-Suarez, Manuel Gamez, Tomas Cabello, and Juan R. Gallego. 2021. "Potential for the Postharvest Biological Control of Phthorimaea operculella (Lepidoptera, Gelechiidae) by Blattisocius tarsalis (Mesostigmata, Blattisociidae)" Agronomy 11, no. 2: 288. https://doi.org/10.3390/agronomy11020288
APA StyleGavara, J., Piedra-Buena, A., Hernandez-Suarez, E., Gamez, M., Cabello, T., & Gallego, J. R. (2021). Potential for the Postharvest Biological Control of Phthorimaea operculella (Lepidoptera, Gelechiidae) by Blattisocius tarsalis (Mesostigmata, Blattisociidae). Agronomy, 11(2), 288. https://doi.org/10.3390/agronomy11020288