
Influence of Physicochemical Characteristics of Bean Crop Soil in Trichoderma spp. Development

 ,
,  , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Trichoderma Isolates Used in this Study
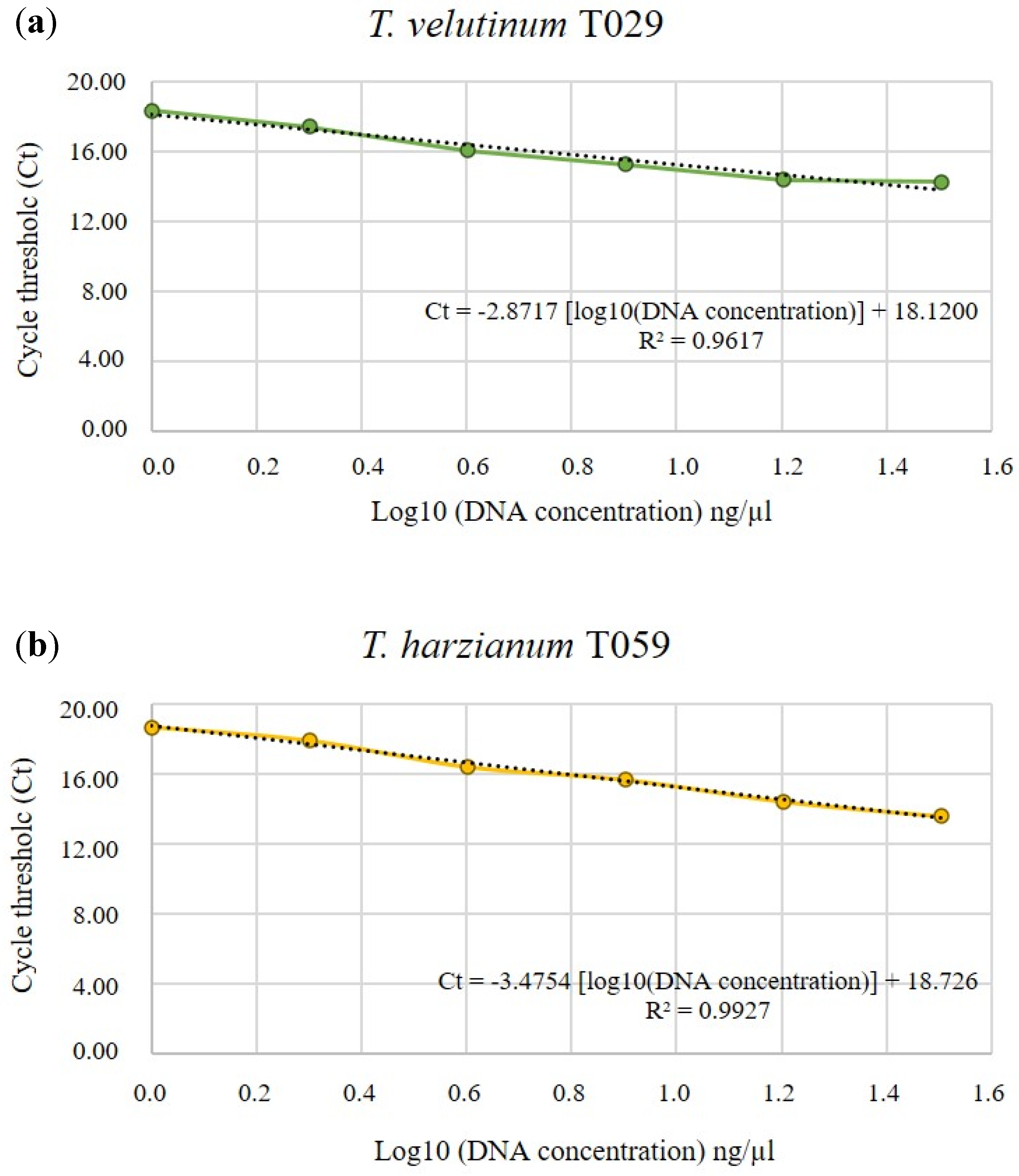
2.2. Standard Curve of Each Isolated
2.3. Soil Sampling
2.4. Physicochemical Analysis of Soils
2.5. Soil Inoculation
2.6. DNA Extraction of Soil Samples and Real Time-PCR (qPCR) Analysis
2.7. Statistical Analysis
3. Results
3.1. Standard curve of Trichoderma spp.
3.2. Soil Sampling and Physicochemical Analysis
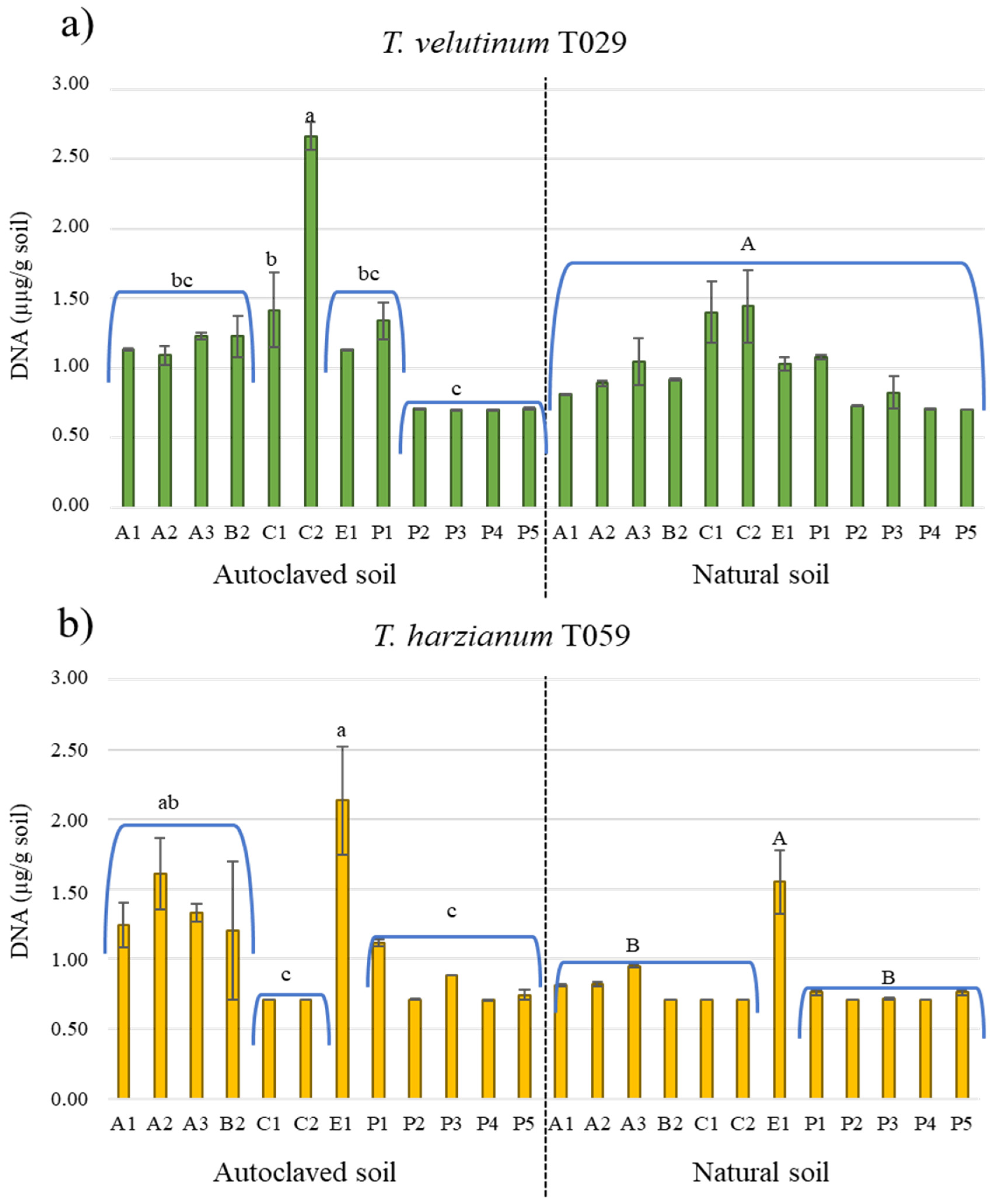
3.3. Development of Trichoderma spp. in Crop Soils
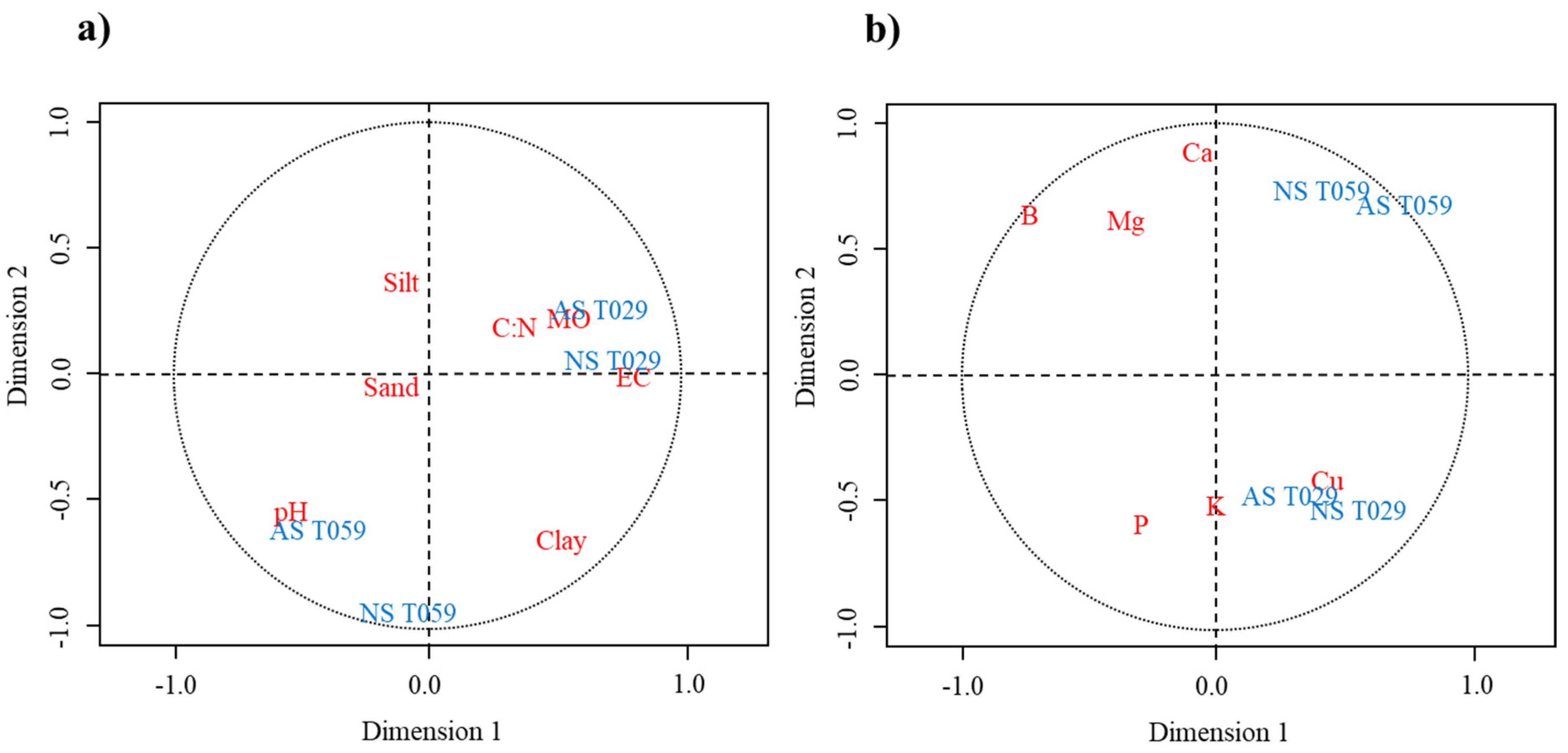
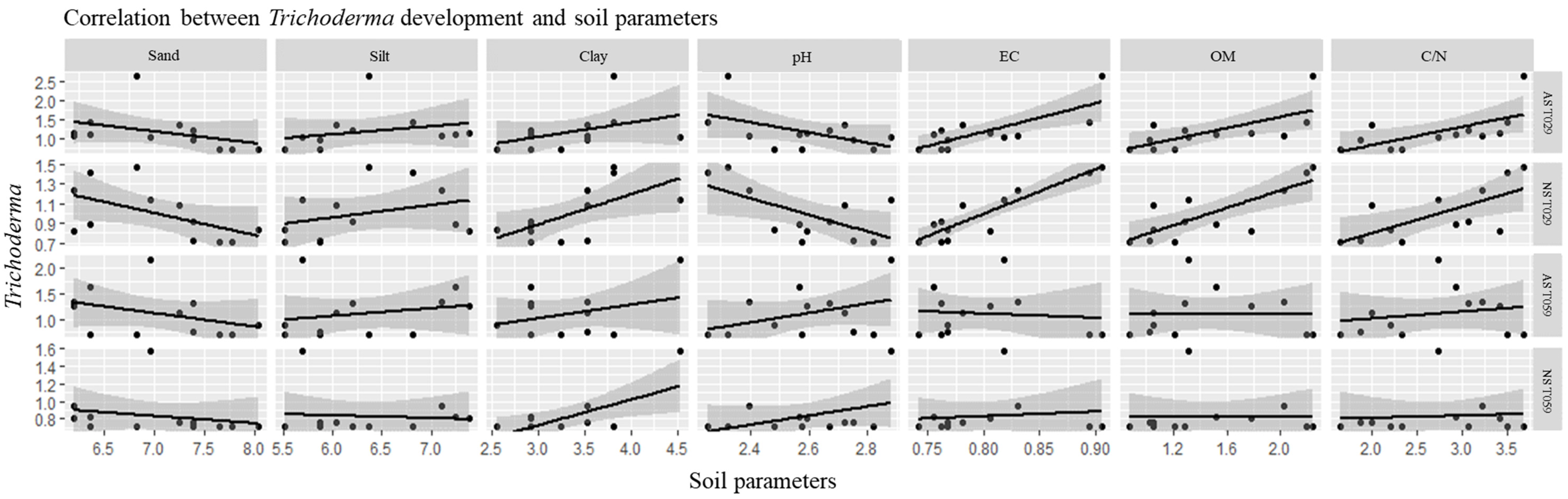
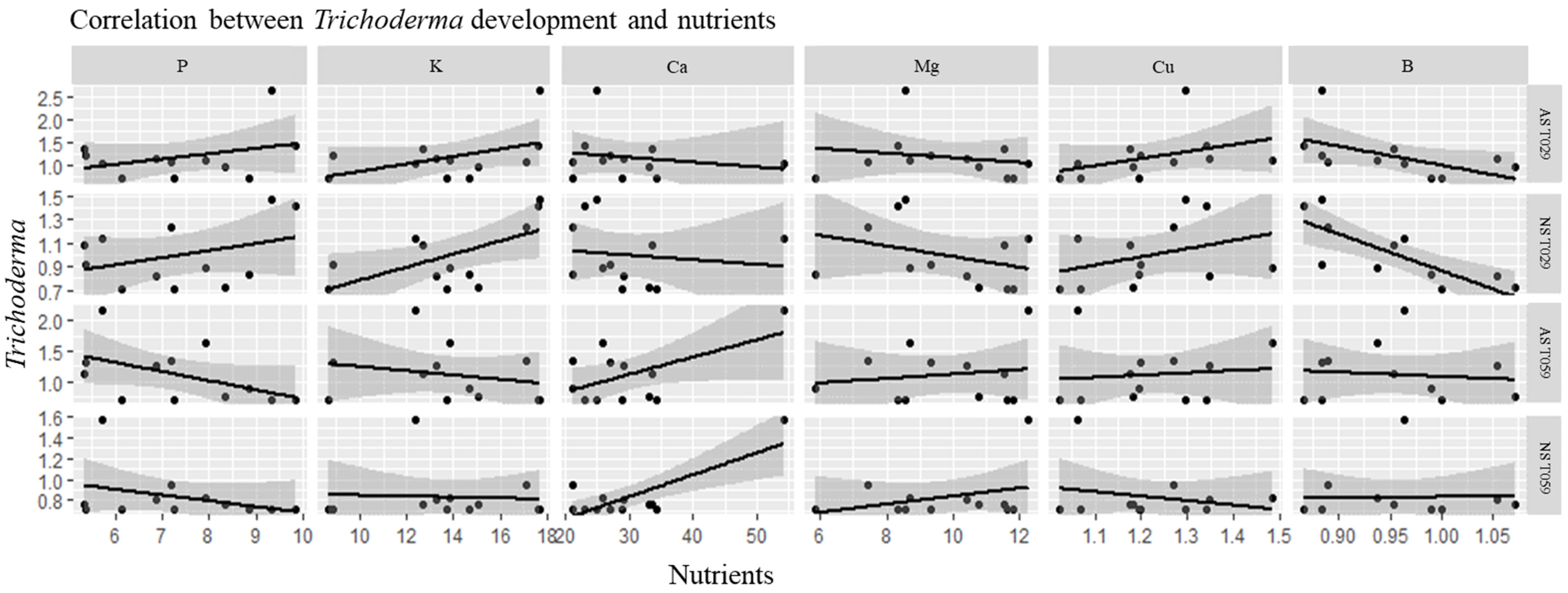
3.4. Correlations of Trichoderma Isolates and Physicochemical Characteristics of Soil Samples and CCA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- FAO FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 1 October 2020).
- Ministerio de Agricultura, Pesca y Alimentación. Anuario de Estadística. Available online: https://www.mapa.gob.es/es/estadistica/temas/publicaciones/anuario-de-estadistica/2019/default.aspx?parte=3&capitulo=07&grupo=2&seccion=2 (accessed on 22 April 2020).
- Casquero, P.A.; Reinoso, B.; Valenciano, J.B. Preliminary evaluation of common bean landraces from León (Spain). In Proceedings of the Annual Report of the Bean Improvement Cooperative, Sacramento, CA, USA, 24 October 2004; Volume 47, pp. 93–94. [Google Scholar]
- Beebe, S.E.; Corrales, M.P. Breeding for disease resistance. In Common Beans: Research for Crop Improvement; Schoonhoven, A., van Voysest, O., Eds.; CAB International: Wallingford, UK, 1991; ISBN 0-85198-679-X. [Google Scholar]
- Gupta, S.; Mathew, K.; Shyam, K.; Sharma, A. Fungicidal management of root rot (Rhizoctonia solani) of French bean. Plant Dis. Res. 1999, 14, 20–24. [Google Scholar]
- Bordoloi, D.; Paul, S.; Das, R.; Roy, A. Effect of pre-sowing seed treatment on emergence and seedling mortality of French bean. Ann. Agric. Res. 1998, 19, 348–349. [Google Scholar]
- Valenciano, J.B.; Casquero, P.A.; Boto, J.A. Influence of sowing techniques and pesticide application on the emergence and the establishment of bean plants (Phaseolus vulgaris L.). Agronomie 2004, 24, 113–118. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Knight, R. Global patterns in bacterial diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 11436–11440. [Google Scholar] [CrossRef]
- Zhong, W.; Gu, T.; Wang, W.; Zhang, B.; Lin, X.; Huang, Q.; Shen, W. The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant Soil 2010, 326, 511–522. [Google Scholar] [CrossRef]
- Kuramae, E.E.; Yergeau, E.; Wong, L.C.; Pijl, A.S.; Veen, J.A.; Kowalchuk, G.A. Soil characteristics more strongly influence soil bacterial communities than land-use type. FEMS Microbiol. Ecol. 2012, 79, 12–24. [Google Scholar] [CrossRef]
- Valenciano, J.B.; Casquero, P.A.; Boto, J.A.; Guerra, M. Effect of sowing techniques and seed pesticide application on dry bean yield and harvest components. Field Crops Res. 2006, 96, 2–12. [Google Scholar] [CrossRef]
- Valenciano, J.B.; Casquero, P.A.; Boto, J.A. Evaluation of the occurrence of bean plants (Phaseolus vulgaris L.) affected by bean seed fly, Delia platura (Meigen), grown under different sowing techniques and with different forms of pesticide application. Field Crops Res. 2004, 85, 103–109. [Google Scholar] [CrossRef]
- Mayo, S.; Cominelli, E.; Sparvoli, F.; González-López, O.; Rodríguez-González, A.; Gutiérrez, S.; Casquero, P.A. Development of a qPCR strategy to select bean genes involved in plant defense response and regulated by the Trichoderma velutinum—Rhizoctonia solani interaction. Front. Plant Sci. 2016, 7, 1109. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Papavizas, G.C. Trichoderma and Gliocladium: Biology, ecology, and potential for biocontrol. Annu. Rev. Phytopathol. 1985, 23, 23–54. [Google Scholar] [CrossRef]
- Mayo, S.; Gutierrez, S.; Malmierca, M.G.; Lorenzana, A.; Campelo, M.P.; Hermosa, R.; Casquero, P.A. Influence of Rhizoctonia solani and Trichoderma spp. in growth of bean (Phaseolus vulgaris L.) and in the induction of plant defense-related genes. Front. Plant Sci. 2015, 6, 685. [Google Scholar] [CrossRef] [PubMed]
- Cardoza, R.E.; McCormick, S.P.; Malmierca, M.G.; Olivera, E.R.; Alexander, N.J.; Monte, E.; Gutiérrez, S. Effects of trichothecene production on the plant defense response and fungal physiology: Overexpression of the Trichoderma arundinaceum tri4 gene in T. harzianum. Appl. Environ. Microbiol. 2015, 81, 6355–6366. [Google Scholar] [CrossRef] [PubMed]
- Gomes, E.V.; Ulhoa, C.J.; Cardoza, R.E.; Silva, R.N.; Gutiérrez, S. Involvement of Trichoderma harzianum Epl-1 protein in the regulation of Botrytis virulence- and tomato defense-related genes. Front. Plant Sci. 2017, 8, 880. [Google Scholar] [CrossRef] [PubMed]
- Hermosa, R.; Rubio, M.B.; Cardoza, R.E.; Nicolás, C.; Monte, E.; Gutiérrez, S. The contribution of Trichoderma to balancing the costs of plant growth and defense. Int. Microbiol. 2013, 16, 69–80. [Google Scholar]
- Hermosa, R.; Cardoza, R.E.; Rubio, M.B.; Gutiérrez, S.; Monte, E. Secondary metabolism and antimicrobial metabolites of Trichoderma. In Biotechnology and Biology of Trichoderma; Gupta, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M.G., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2014; pp. 125–137. ISBN 9780444595768. [Google Scholar]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; López-Bucio, J.S.; López-Bucio, J. Enhanced plant immunity using Trichoderma. In Biotechnology and Biology of Trichoderma; Gupta, V.G., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M., Eds.; Elsevier: Oxford, UK, 2014; pp. 495–504. ISBN 9780444595768. [Google Scholar]
- Mayo-Prieto, S.; Porteous-Álvarez, A.J.; Rodríguez-González, Á.; Gutiérrez, S.; Casquero, P.A. Evaluation of substrates and additives to Trichoderma harzianum development by qPCR. Agron. J. 2020, 112, 3188–3194. [Google Scholar] [CrossRef]
- Beaulieu, R.; López-Mondéjar, R.; Tittarelli, F.; Ros, M.; Pascual, J.A. qRT-PCR quantification of the biological control agent Trichoderma harzianum in peat and compost-based growing media. Bioresour. Technol. 2011, 102, 2793–2798. [Google Scholar] [CrossRef]
- Legrand, F.; Picot, A.; Cobo-Díaz, J.F.; Cor, O.; Barbier, G.; Le Floch, G. Development of qPCR assays to monitor the ability of Gliocladium catenulatum J1446 to reduce the cereal pathogen Fusarium graminearum inoculum in soils. Eur. J. Plant Pathol. 2018, 152, 285–295. [Google Scholar] [CrossRef]
- Woodhall, J.W.; Adams, I.P.; Peters, J.C.; Harper, G.; Boonham, N. A new quantitative real-time PCR assay for Rhizoctonia solani AG3-PT and the detection of AGs of Rhizoctonia solani associated with potato in soil and tuber samples in Great Britain. Eur. J. Plant Pathol. 2013, 136, 273–280. [Google Scholar] [CrossRef]
- Demeke, T.; Gräfenhan, T.; Clear, R.M.; Phan, A.; Ratnayaka, I.; Chapados, J.; Patrick, S.K.; Gaba, D.; Lévesque, C.A.; Seifert, K.A. Development of a specific TaqMan® real-time PCR assay for quantification of Fusarium graminearum clade 7 and comparison of fungal biomass determined by PCR with deoxynivalenol content in wheat and barley. Int. J. Food Microbiol. 2010, 141, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Fernández, D.; Montes-Borrego, M.; Navas-Cortés, J.A.; Jiménez-Díaz, R.M.; Landa, B.B. Identification and quantification of Fusarium oxysporum in planta and soil by means of an improved specific and quantitative PCR assay. Appl. Soil Ecol. 2010, 46, 372–382. [Google Scholar] [CrossRef]
- Li, M.; Senda, M.; Komatsu, T.; Suga, H.; Kageyama, K. Development of real-time PCR technique for the estimation of population density of Pythium intermedium in forest soils. Microbiol. Res. 2010, 165, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Mayo-Prieto, S.; Campelo, M.P.; Lorenzana, A.; Rodríguez-González, A.; Reinoso, B.; Gutiérrez, S.; Casquero, P.A. Antifungal activity and bean growth promotion of Trichoderma strains isolated from seed vs soil. Eur. J. Plant Pathol. 2020, 158, 817–828. [Google Scholar] [CrossRef]
- Cardoza, R.E.; Hermosa, M.R.; Vizcaíno, J.A.; González, F.; Llobell, A.; Monte, E.; Gutiérrez, S. Partial silencing of a hydroxy-methylglutaryl-CoA reductase-encoding gene in Trichoderma harzianum CECT 2413 results in a lower level of resistance to lovastatin and lower antifungal activity. Fungal Genet. Biol. 2007, 44, 269–283. [Google Scholar] [CrossRef]
- Restovich, S.B.; Andriulo, A.E.; Portela, S.I. Introduction of cover crops in a maize–soybean rotation of the Humid Pampas: Effect on nitrogen and water dynamics. Filed Crop. Res. 2012, 128, 62–70. [Google Scholar] [CrossRef]
- MAPA. Métodos Oficales de Análisis. Tomo III. Suelos, Aguas, Fertilizantes y Plantas; Secretaría General de Alimentación. Dirección General de Política Alimentaria, Ed.; Ministerio de Agricultura Pesca Y Alimentacion: Madrid, Spain, 1994.
- R Core Team R: A Language and Environment for Statistical Computing 2013. Available online: https://www.scirp.org/(S(351jmbntvnsjt1aadkposzje))/reference/ReferencesPapers.aspx?ReferenceID=1787696 (accessed on 31 March 2020).
- Ozyilmaz, U.; Benlioglu, K.; Yildiz, A.; Benlioglu, H.S. Effects of soil amendments combined with solarization on the soil microbial community in strawberry cultivation using quantitative real-time PCR. Phytoparasitica 2016, 44, 661–680. [Google Scholar] [CrossRef]
- Harries, E.; Berruezo, L.A.; Galván, M.Z.; Rajal, V.B.; Mercado Cárdenas, G.E. Soil properties related to suppression of Rhizoctonia solani on tobacco fields from northwest Argentina. Plant Pathol. 2020, 69, 77–86. [Google Scholar] [CrossRef]
- Dong, L.; Guo, Q.; Wang, P.; Zhang, X.; Su, Z.; Zhao, W.; Lu, X.; Li, S.; Ma, P. Qualitative and quantitative analyses of the colonization characteristics of Bacillus subtilis strain NCD-2 on cotton root. Curr. Microbiol. 2020. [Google Scholar] [CrossRef]
- Al-Khaliel, A.S. Effects of arbuscular mycorrhization in sterile and non-sterile soils. Trop. Life Sci. Res. 2010, 21, 55. [Google Scholar] [PubMed]
- Linderman, R.G. Vesicular-arbuscular mycorrhizae and soil microbial interactions. In Micorrhizae in Sustainable Agriculture; Bethlenfalvay, G.J., Linderman, R.G., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1992; Volume 54, pp. 45–70. [Google Scholar]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for management of soilborne siseases in crop production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef]
- Bodelier, P.L.E. Toward understanding, managing, and protecting microbial ecosystems. Front. Microbiol. 2011, 2, 80. [Google Scholar] [CrossRef] [PubMed]
- Tajik, S.; Ayoubi, S.; Lorenz, N. Soil microbial communities affected by vegetation, topography and soil properties in a forest ecosystem. Appl. Soil Ecol. 2020, 149, 103514. [Google Scholar] [CrossRef]
- Latour, X.; Corberand, T.; Laguerre, G.; Allard, F.; Lemanceau, P. The composition of fluorescent pseudomonad populations associated with roots is influenced by plant and soil type. Appl. Environ. Microbiol. 1996, 62, 2449–2456. [Google Scholar] [CrossRef]
- Li, D.; Sharp, J.O.; Saikaly, P.E.; Ali, S.; Alidina, M.; Alarawi, M.S.; Keller, S.; Hoppe-Jones, C.; Drewes, J.E. Dissolved organic carbon influences microbial community composition and diversity in managed aquifer recharge systems. Appl. Environ. Microbiol. 2012, 78, 6819–6828. [Google Scholar] [CrossRef]
- Van der Bom, F.; Nunes, I.; Raymond, N.S.; Hansen, V.; Bonnichsen, L.; Magid, J.; Nybroe, O.; Jensen, L.S. Long-term fertilisation form, level and duration affect the diversity, structure and functioning of soil microbial communities in the field. Soil Biol. Biochem. 2018, 122, 91–103. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Gerard, E.; van Koten, C.; Banabas, M.; O’Callaghan, M.; Nelson, P.N. Soil physicochemical properties impact more strongly on bacteria and fungi than conversion of grassland to oil palm. Pedobiologia (Jena) 2016, 59, 83–91. [Google Scholar] [CrossRef]
- Brockett, B.F.T.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in Western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Pan, X.; Zhang, S.; Zhong, Q.; Gong, G.; Wang, G.; Guo, X.; Xu, X. Effects of soil chemical properties and fractions of Pb, Cd, and Zn on bacterial and fungal communities. Sci. Total Environ. 2020, 715, 136904. [Google Scholar] [CrossRef]
- Bach, E.M.; Baer, S.G.; Meyer, C.K.; Six, J. Soil texture affects soil microbial and structural recovery during grassland restoration. Soil Biol. Biochem. 2010, 42, 2182–2191. [Google Scholar] [CrossRef]
- Gorissen, A.; Jansen, A.E.; Olsthoorn, A.F.M. Effects of a two-year application of ammonium sulphate on growth, nutrient uptake, and rhizosphere microflora of juvenile Douglas-fir. Plant Soil 1993, 157, 41–50. [Google Scholar] [CrossRef]
- Demoling, F.; Figueroa, D.; Bååth, E. Comparison of factors limiting bacterial growth in different soils. Soil Biol. Biochem. 2007, 39, 2485–2495. [Google Scholar] [CrossRef]
- Simon, A.; Sivasithamparam, K. Microbiological differences between soils suppresive and conductive of the saprophytic growth of Gaeumannomyces graminis var. tritici. Can. J. Microbiol. 1988, 34, 860–864. [Google Scholar] [CrossRef]
- Duffy, B.K.; Ownley, H.; Weller, D.M. Soil chemical and physical properties associated with suppression of take-all of wheat by Trichoderma koningii. Phytopathology 1997, 87, 1118–1124. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Productive Zone | Code | Place | Crop |
|---|---|---|---|
| Astorga | A1 | Otero de Escarpizo | Phaseolus vulgaris (Bean) |
| A2 | Otero de Escarpizo | Triticum aestivum (Wheat) | |
| A3 | Sueros de Cepeda | P. vulgaris (Bean) | |
| La Bañeza | B2 | San Juan de Torres | P. vulgaris (Bean) |
| La Cabrera | C1 | Castrocontrigo | P. vulgaris (Bean) |
| C2 | Castrocontrigo | Solanum tuberosum (Potato) | |
| Esla-Campos | E1 | Jabares de los Oteros | P. vulgaris (Bean) |
| Páramo | P1 | Bercianos del Páramo | P. vulgaris (Bean) |
| P2 | Bercianos del Páramo | Helianthus annus (Sunflower) | |
| P3 | Bustillo del Páramo | P. vulgaris (Bean) | |
| P4 | La Milla del Páramo | Beta vulgaris (Beat) | |
| P5 | La Milla del Páramo | P. vulgaris (Bean) |
| Id | Sand (%) | Silt (%) | Clay (%) | Texture | pH | CEC (cmol kg−1) | EC (dS m−1) | OM (%) | Ctotal (%) | Ntotal (%) | C/N | P (ppm) | K (ppm) | Ca (ppm) | Mg (ppm) | K/Mg | Ca/Mg | Na (ppm) | Mn (ppm) | Fe (ppm) | Cu (ppm) | Zn (ppm) | B (ppm) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | 38 | 54 | 8 | Silt loam | 6.23 | 6.79 | 0.15 | 2.67 | 1.50 | 0.14 | 11.22 | 46.53 | 175.94 | 843.64 | 108.16 | 1.63 | 7.80 | 45.98 | 22.23 | 46.37 | 1.32 | 2.24 | 0.61 |
| A2 | 40 | 52 | 8 | Silt loam | 6.09 | 4.92 | 0.07 | 1.81 | 1.07 | 0.13 | 8.11 | 62.36 | 191.58 | 661.29 | 75.35 | 2.54 | 8.78 | 13.79 | 18.98 | 55.84 | 1.70 | 2.56 | 0.38 |
| A3 | 38 | 50 | 12 | Silt loam | 5.25 | 3.69 | 0.19 | 3.58 | 2.08 | 0.21 | 9.90 | 51.18 | 293.24 | 446.87 | 54.69 | 5.36 | 8.17 | 20.69 | 32.74 | 40.11 | 1.11 | 1.14 | 0.29 |
| B2 | 54 | 38 | 8 | Sandy loam | 6.63 | 4.94 | 0.08 | 1.15 | 0.67 | 0.07 | 8.94 | 28.05 | 78.20 | 733.43 | 86.28 | 0.91 | 8.50 | 11.49 | 19.17 | 35.13 | 0.94 | 0.76 | 0.28 |
| C1 | 40 | 46 | 14 | Loam | 4.58 | 4.01 | 0.30 | 4.31 | 2.47 | 0.21 | 11.81 | 96.67 | 308.88 | 529.03 | 69.27 | 4.46 | 7.64 | 9.20 | 48.53 | 91.69 | 1.30 | 2.18 | 0.25 |
| C2 | 46 | 40 | 14 | Loam | 4.90 | 4.70 | 0.32 | 4.51 | 2.66 | 0.20 | 13.09 | 86.27 | 312.79 | 615.20 | 72.92 | 4.29 | 8.44 | 22.99 | 37.61 | 75.92 | 1.18 | 2.30 | 0.28 |
| E1 | 48 | 32 | 20 | Loam | 7.79 | 8.46 | 0.17 | 1.22 | 0.82 | 0.10 | 7.00 | 31.93 | 152.48 | 2941.73 | 149.48 | 1.02 | 19.68 | 18.39 | 18.63 | 14.20 | 0.63 | 0.66 | 0.43 |
| P1 | 52 | 36 | 12 | Sandy loam | 6.91 | 7.05 | 0.11 | 0.62 | 0.81 | 0.10 | 3.53 | 27.90 | 160.30 | 1128.20 | 132.46 | 1.21 | 8.52 | 13.79 | 32.52 | 33.97 | 0.88 | 0.67 | 0.41 |
| P2 | 58 | 34 | 8 | Sandy loam | 7.45 | 5.52 | 0.05 | 0.27 | 0.45 | 0.07 | 2.25 | 36.90 | 74.29 | 839.63 | 138.54 | 0.54 | 6.06 | 9.20 | 14.74 | 22.11 | 0.64 | 0.96 | 0.50 |
| P3 | 64 | 30 | 6 | Sandy loam | 5.66 | 3.32 | 0.09 | 0.62 | 0.54 | 0.08 | 4.38 | 77.73 | 215.04 | 442.86 | 34.03 | 6.32 | 13.02 | 2.30 | 41.29 | 64.33 | 0.93 | 1.20 | 0.48 |
| P4 | 60 | 30 | 10 | Sandy loam | 6.13 | 7.71 | 0.08 | 0.97 | 0.89 | 0.11 | 4.94 | 52.11 | 187.67 | 1180.30 | 134.89 | 1.39 | 8.75 | 13.79 | 25.73 | 50.15 | 0.55 | 0.92 | 0.50 |
| P5 | 54 | 34 | 12 | Sandy loam | 7.06 | 7.44 | 0.09 | 0.56 | 0.77 | 0.11 | 3.06 | 69.19 | 226.77 | 1102.15 | 115.45 | 1.96 | 9.55 | 13.79 | 23.98 | 31.90 | 0.90 | 1.28 | 0.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayo-Prieto, S.; Porteous-Álvarez, A.J.; Mezquita-García, S.; Rodríguez-González, Á.; Carro-Huerga, G.; del Ser-Herrero, S.; Gutiérrez, S.; Casquero, P.A. Influence of Physicochemical Characteristics of Bean Crop Soil in Trichoderma spp. Development. Agronomy 2021, 11, 274. https://doi.org/10.3390/agronomy11020274
Mayo-Prieto S, Porteous-Álvarez AJ, Mezquita-García S, Rodríguez-González Á, Carro-Huerga G, del Ser-Herrero S, Gutiérrez S, Casquero PA. Influence of Physicochemical Characteristics of Bean Crop Soil in Trichoderma spp. Development. Agronomy. 2021; 11(2):274. https://doi.org/10.3390/agronomy11020274
Chicago/Turabian StyleMayo-Prieto, Sara, Alejandra J. Porteous-Álvarez, Sergio Mezquita-García, Álvaro Rodríguez-González, Guzmán Carro-Huerga, Sara del Ser-Herrero, Santiago Gutiérrez, and Pedro A. Casquero. 2021. "Influence of Physicochemical Characteristics of Bean Crop Soil in Trichoderma spp. Development" Agronomy 11, no. 2: 274. https://doi.org/10.3390/agronomy11020274
APA StyleMayo-Prieto, S., Porteous-Álvarez, A. J., Mezquita-García, S., Rodríguez-González, Á., Carro-Huerga, G., del Ser-Herrero, S., Gutiérrez, S., & Casquero, P. A. (2021). Influence of Physicochemical Characteristics of Bean Crop Soil in Trichoderma spp. Development. Agronomy, 11(2), 274. https://doi.org/10.3390/agronomy11020274