
The Comparison of Juvenile Hormone and Transcriptional Changes between Three Different Juvenile Hormone Analogs Insecticides on Honey Bee Worker Larval’s Development

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Honey Bee Larvae Rearing
2.2. Extraction and Determination of JH Titers in Honey Bees
2.3. RNA Extraction, Library Preparation, and Sequencing
2.4. Transcriptomic Data Analysis
2.5. Functional Analyses of DEGs
2.6. Validation of Four DEGs
2.7. Statistics
3. Result
3.1. Determination of JH Titers in Six-Day-Old Worker Larvae
3.2. Quality Analysis of Transcriptome Sequencing Results
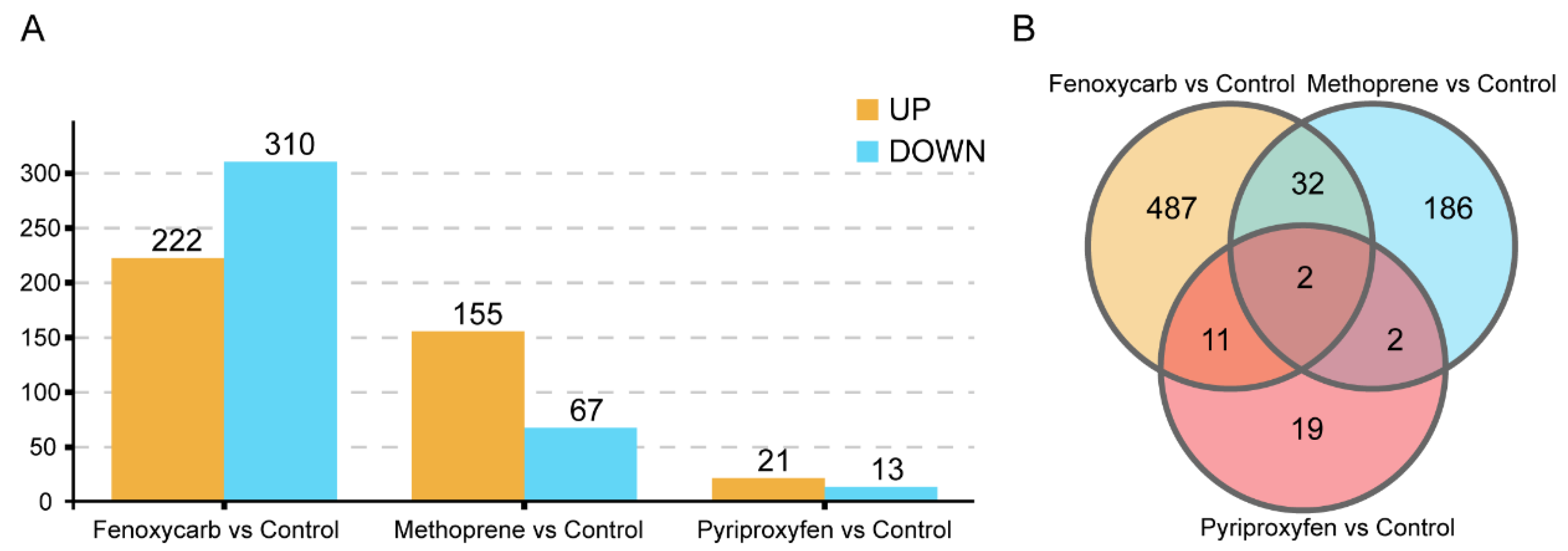
3.3. Screening of DEGs
3.4. GO Functional Annotation and KEGG Pathway
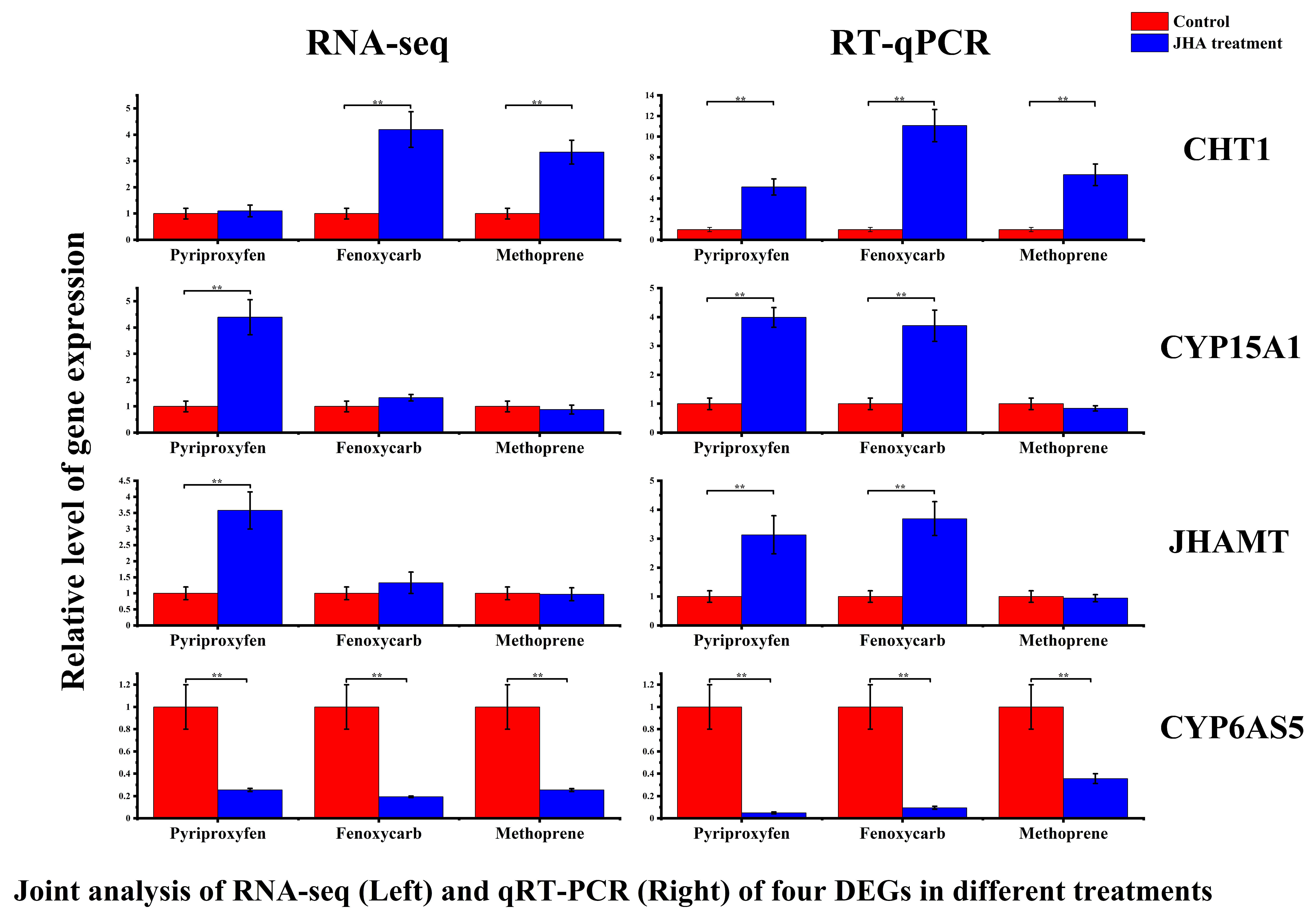
3.5. Validation of DEGs by Using RT-qPCR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| JH | Juvenile hormone |
| JHA | Juvenile hormone analog |
| CHT1 | Chitinase |
| CYP15A1 | Cytochrome CYP15A1 |
| CYP6AS5 | Cytochrome CYP6AS5 |
| JHAMT | Juvenile hormone acid methyltransferase |
| FPKM | Fragments Per Kilobase per Million |
| DMSO | Dimethyl sulfoxide |
| CK | Control check |
| Bbm | Pyriproxyfen |
| Byw | Fenoxycarb |
| Xcz | Methoprene |
References
- Ikeda, T.; Furukawa, S.; Nakamura, J.; Sasaki, M.; Sasaki, T. CpG methylation in the hexamerin 110 gene in the European honeybee, Apis mellifera. Insect Sci. 2011, 11, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sillman, J.; Uusitalo, V.; Tapanen, T.; Salonen, A.; Soukka, R.; Kahiluoto, H. Contribution of honeybees towards the net environmental benefits of food. Sci. Total Environ. 2021, 756, 143880. [Google Scholar] [CrossRef]
- Nakamura, S.; Yamamoto, S.; Sawamura, N.; Nikkeshi, A.; Kishi, S.; Kamo, T. Pollination effectiveness of European honeybee, Apis mellifera (Hymenoptera: Apidae), in an Oriental persimmon, Diospyros kaki (Ericales: Ebenaceae), orchard. Appl. Entomol. Zool. 2020, 55, 405–412. [Google Scholar] [CrossRef]
- Dearden, P.K.; Duncan, E.J.; Wilson, M.J. The honeybee Apis mellifera. Cold Spring Harb. Protoc. 2009, 2009, emo123. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X.; Zhang, Z.; Lang, H.; Zheng, H. Honey bee as a model organism to study gut microbiota and diseases. Drug Discov. Today Dis. Models. 2018, 28, 35–42. [Google Scholar] [CrossRef]
- Kohno, H.; Kubo, T. Genetics in the Honey Bee: Achievements and Prospects toward the Functional Analysis of Molecular and Neural Mechanisms Underlying Social Behaviors. Insects 2019, 10, 348. [Google Scholar] [CrossRef] [Green Version]
- Richards, M.H. Socially polymorphic bees as model organisms for studying the evolution of eusociality. Insectes Sociaux. 2019, 66, 3–4. [Google Scholar] [CrossRef]
- Kelstrup, H.C.; Hartfelder, K.; Nascimento, F.S.; Riddiford, L.M. The role of juvenile hormone in dominance behavior, reproduction and cuticular pheromone signaling in the caste-flexible epiponine wasp, Synoeca surinama. Front. Zool. 2014, 11, 78. [Google Scholar] [CrossRef] [Green Version]
- Stay, B.; Tobe, S.S. The role of allatostatins in juvenile hormone synthesis in insects and crustaceans. Annu. Rev. Entomol. 2007, 52, 277–299. [Google Scholar] [CrossRef]
- Denlinger, D.L.; Yocum, G.D.; Rinehart, J.P. Hormonal Control of Diapause. Endocrinology 2012, 430–463. [Google Scholar] [CrossRef]
- Wyatt, G.R.; Davey, K.G. Cellular and molecular actions of juvenile hormone. II. Roles of juvenile hormone in adult insects. Adv. Insect Physiol. 1996, 6, 1–155. [Google Scholar] [CrossRef]
- Fei, H.; Martin, T.R.; Jaskowiak, K.M.; Hatle, J.D.; Whitman, D.W.; Borst, D.W. Starvation affects vitellogenin production but not vitellogenin mRNA levels in the lubber grasshopper, Romalea microptera. J. Insect Physiol. 2005, 51, 435–443. [Google Scholar] [CrossRef]
- Xiao, H.J.; Fu, X.W.; Liu, Y.Q.; Wu, K.M. Synchronous vitellogenin expression and sexual maturation during migration are negatively correlated with juvenile hormone levels in Mythimna separata. Sci. Rep. 2016, 6, 33309. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Brent, C.S.; Fennern, E.; Amdam, G.V. Gustatory perception and fat body energy metabolism are jointly affected by vitellogenin and juvenile hormone in honey bees. PLoS Genet. 2012, 8, e1002779. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Cheng, L.; Chapman, J.W.; Sappington, T.W.; Liu, J.; Cheng, Y.; Jiang, X. Juvenile hormone regulates the shift from migrants to residents in adult oriental armyworm, Mythimna separata. Sci. Rep. 2020, 10, 11626. [Google Scholar] [CrossRef] [PubMed]
- Araki, M.; Miyakawa, M.O.; Suzuki, T.; Miyakawa, H. Two insulin-like peptides may regulate egg production in opposite directions via juvenile hormone signaling in the queenless ant Pristomyrmex punctatus. J. Exp. Zool. Part B Mol. Dev. Evol. 2020, 334, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.G. The molecular site of action of juvenile hormone and juvenile hormone insecticides during metamorphosis: How these compounds kill insects. J. Insect. Physiol. 2004, 50, 111–121. [Google Scholar] [CrossRef]
- Saltzmann, K.A.; Saltzmann, K.D.; Neal, J.J.; Scharf, M.E.; Bennett, G.W.J. Effects of the juvenile hormone analog pyriproxyfen on German cockroach, Blattella germanica (L.), tergal gland development and production of tergal gland secretion proteins. Arch. Insect Biochem. Physiol. 2010, 63, 15–23. [Google Scholar] [CrossRef]
- Cripe, G.M.; Mckenney, C.L.; Hoglund, M.D.; Harris, P.S. Effects of fenoxycarb exposure on complete larval development of the xanthid crab, Rhithropanopeus harrisii. Environ. Pollut. 2003, 125, 295–299. [Google Scholar] [CrossRef]
- Hu, X.L.; Niu, J.J.; Meng, Q.; Chai, Y.H.; Chu, K.H.; Chan, K.M. Effects of two juvenile hormone analogue insecticides, fenoxycarb and methoprene, on Neocaridina davidi. Environ. Pollut. 2019, 253, 89–99. [Google Scholar] [CrossRef]
- Hu, X.L.; Tang, Y.Y.; Kwok, M.L.; Chan, K.M.; Chu, K.H. Impact of juvenile hormone analogue insecticides on the water flea Moina macrocopa: Growth, reproduction and transgenerational effect. Aquat. Toxicol. 2020, 220, 105402. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, A.; Hafeez, F.; Hafeez, M.; Farooq, M.; Asif Aziz, M.; Sohaib, M.; Naeem, A.; Lu, Y. Sublethal effects of a juvenile hormone analog, Pyriproxyfen on demographic parameters of non-target predator, Hippodamia convergens Guerin-Meneville (Coleoptera: Coccinellidae). Ecotoxicology 2020, 29, 1017–1028. [Google Scholar] [CrossRef]
- Horton, D.R.; Lewis, T.M.J. Effects of fenoxycarb on ovarian development, spring fecundity and longevity in winterform pear psylla. Entomologia Experimentalis Applicata. 2015, 81, 181–187. [Google Scholar] [CrossRef]
- Adnan, S.M.; Farhana, I.; Rempoulakis, P.; Taylor, P.W. Methoprene-induced matings of young Queensland fruit fly males are effective at inducing sexual inhibition in females. J. Appl. Entomol. 2020, 144, 500–508. [Google Scholar] [CrossRef]
- Boleli, I.C.; Bitondi, M.; Figueiredo, V.; Simões, Z. Mispatterning in the ommatidia of Apis mellifera pupae treated with a juvenile hormone analogue. J. Morphol. 2010, 249, 89–99. [Google Scholar] [CrossRef]
- Harano, K.I. Effects of juvenile hormone analog on physiological and behavioral maturation in honeybee drones. Apidologie 2013, 44, 586–599. [Google Scholar] [CrossRef]
- Fujita, K.; Shimomura, K.; Yamamoto, K.; Yamashita, T.; Suzuki, K. A chitinase structurally related to the glycoside hydrolase family 48 is indispensable for the hormonally induced diapause termination in a beetle. Biochem. Biophys. Res. Commun. 2006, 345, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Helvig, C.; Koener, J.F.; Unnithan, G.C.; Feyereisen, R. CYP15A1, the cytochrome P450 that catalyzes epoxidation of methyl farnesoate to juvenile hormone III in cockroach corpora allata. Proc. Natl. Acad. Sci. USA 2004, 101, 4024–4029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Veeran, S.; Wang, J.; Li, S.; Li, K.; Liu, S.N. Dual roles of juvenile hormone signaling during early oogenesis in Drosophila. Insect Sci. 2020, 27, 665–674. [Google Scholar] [CrossRef]
- Tavares, D.A.; Dussaubat, C.; Kretzschmar, A.; Carvalho, S.M.; Silva-Zacarin, E.C.M.; Malaspina, O.; Berail, G.; Brunet, J.L.; Belzunces, L.P. Exposure of larvae to thiamethoxam affects the survival and physiology of the honey bee at post-embryonic stages. Environ. Pollut. 2017, 229, 386–393. [Google Scholar] [CrossRef]
- Tasei, J. Impact of agrochemicals on non-Apis bees. In Honey Bees: Estimating the Environmental Impact of Chemicals; CRC Press: Boca Raton, FL, USA, 2002; ISBN 978-0-415-27518-7. [Google Scholar]
- Milchreit, K.; Ruhnke, H.; Wegener, J.; Bienefeld, K.J.E. Effects of an insect growth regulator and a solvent on honeybee (Apis mellifera L.) brood development and queen viability. Ecotoxicology 2016, 25, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Sasagawa, H.; Kuwahara, Y. Quantitative Determination Method for a Juvenile Hormone III Titer in Honeybee Haemolymph by High-Performance Liquid Chromatography. Agric. Biol. Chem. 2016, 52, 1295–1297. [Google Scholar] [CrossRef]
- Furuta, K.; Ichikawa, A.; Murata, M.; Kuwano, E.; Shiotsuki, T. Determination by LC-MS of Juvenile Hormone Titers in Hemolymph of the Silkworm, Bombyx mori. J. Agric. Chem. Soc. Jpn. 2013, 77, 988–991. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Shpigler, H.Y.; Magory Cohen, T.; Ben-Shimol, E.; Ben-Betzalel, R.; Levin, E. Juvenile hormone functions as a metabolic rate accelerator in bumble bees (Bombus terrestris). Horm. Behav. 2021, 136, 105073. [Google Scholar] [CrossRef]
- Hartfelder, K.; Engels, W. Social Insect Polymorphism: Hormonal Regulation of Plasticity in Development and Reproduction in the Honeybee. Curr. Top. Dev. Biol. 1998, 40, 45–77. [Google Scholar] [CrossRef]
- Barchuk, A.R.; Cristino, A.S.; Kucharski, R.; Costa, L.F.; Simoes, Z.L.; Maleszka, R. Molecular determinants of caste differentiation in the highly eusocial honeybee Apis mellifera. BMC Dev. Biol. 2007, 7, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Huang, Z.Y.; Liu, F.; Li, Z.; Yan, L.; Zhang, S.; Chen, S.; Zhong, B.; Su, S. Molecular cloning and characterization of juvenile hormone acid methyltransferase in the honey bee, Apis mellifera, and its differential expression during caste differentiation. PLoS ONE 2013, 8, e68544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez, C.V.; Maestro, J.L. Expression of juvenile hormone acid O-methyltransferase and juvenile hormone synthesis in Blattella germanica. Insect Sci. 2018, 25, 787–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Tao, T.; Liu, M.; Zhou, Y.; Liu, Z.; Zhu, D. The potential role of juvenile hormone acid methyltransferase in methyl farnesoate (MF) biosynthesis in the swimming crab, Portunus trituberculatus. Anim. Reprod. Sci. 2016, 168, 40–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchal, E.; Zhang, J.; Badisco, L.; Verlinden, H.; Hult, E.F.; van Wielendaele, P.; Yagi, K.J.; Tobe, S.S.; Vanden Broeck, J. Final steps in juvenile hormone biosynthesis in the desert locust, Schistocerca gregaria. Insect Biochem. Mol. Biol. 2011, 41, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.P.; Yin, C.M.; Tatar, M. Mutations in insulin signaling pathway alter juvenile hormone synthesis in Drosophila melanogaster. Gen. Comp. Endocrinol. 2005, 142, 347–356. [Google Scholar] [CrossRef]
- Huang, J.; Marchal, E.; Hult, E.F.; Zels, S.; Vanden Broeck, J.; Tobe, S.S. Mode of action of allatostatins in the regulation of juvenile hormone biosynthesis in the cockroach, Diploptera punctata. Insect Biochem. Mol. Biol. 2014, 54, 61–68. [Google Scholar] [CrossRef]
- Mamidala, P.; Wijeratne, A.J.; Wijeratne, S.; Kornacker, K.; Sudhamalla, B.; Rivera-Vega, L.J.; Hoelmer, A.; Meulia, T.; Jones, S.C.; Mittapalli, O.J.B.G. RNA-Seq and molecular docking reveal multi-level pesticide resistance in the bed bug. BMC Genom. 2012, 13, 6. [Google Scholar] [CrossRef] [Green Version]
- Mao, W.; Berenbaum, S. CYP9Q-mediated detoxification of acaricides in the honey bee (Apis mellifera). Proc. Natl. Acad. Sci. USA. 2011, 108, 12657–12662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Fan, R.L.; Ji, W.N.; Zhang, W.W.; Chen, X.M.; Wang, S.; Yin, L.; Gao, F.C.; Chen, G.H.; Ji, T. Transcriptome Analysis of Newly Emerged Honeybees Exposure to Sublethal Carbendazim During Larval Stage. Front. Genet. 2018, 9, 426. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, S.; Chen, L.; Luo, J.; Zeng, D.; Li, X. Juvenile hormone and transcriptional changes in honey bee worker larvae when exposed to sublethal concentrations of thiamethoxam. Ecotoxicol. Environ. Saf. 2021, 225, 112744. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, T.D.; Unnithan, G.C. A cytochrome P450 terpenoid hydroxylase linked to the suppression of insect juvenile hormone. Proc. Natl. Acad. Sci. USA 1998, 95, 12884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaguchi, H.; Masuoka, Y.; Inoue, T.; Maekawa, K. Expressions of juvenile hormone biosynthetic genes during presoldier differentiation in the incipient colony of Zootermopsis nevadensis (Isoptera: Archotermopsidae). Appl. Entomol. Zool. 2015, 50, 497–508. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Component | Diet A (Day1) | Diet B (Day3) | Diet C (Day4, 5) |
|---|---|---|---|
| Royal jelly | 50% | 50% | 50% |
| Glucose | 6% | 7.5% | 9% |
| Fructose | 6% | 7.5% | 9% |
| Yeast Extract | 1% | 1.5% | 2% |
| H2O | 37% | 33.5% | 30% |
| Gene | Pyriproxyfen | Fenoxycarb | Methoprene |
|---|---|---|---|
| CHT1 | 1.10 | 4.20 | 3.34 |
| CYP6AS5 | 0.26 | 0.19 | 0.25 |
| CYP15A1 | 4.40 | 1.33 | 0.88 |
| JHAMT | 3.58 | 1.33 | 0.97 |
| Gene | Pyriproxyfen | Fenoxycarb | Methoprene |
|---|---|---|---|
| CHT1 | 5.13 | 11.08 | 6.31 |
| CYP6AS5 | 0.05 | 0.09 | 0.36 |
| CYP15A1 | 3.99 | 3.70 | 0.84 |
| JHAMT | 3.13 | 3.69 | 0.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, J.; Liu, S.; Hou, J.; Chen, L.; Li, H.; Liao, S.; Tan, Q.; Yang, T.; Yi, G.; Zhang, F.; et al. The Comparison of Juvenile Hormone and Transcriptional Changes between Three Different Juvenile Hormone Analogs Insecticides on Honey Bee Worker Larval’s Development. Agronomy 2021, 11, 2497. https://doi.org/10.3390/agronomy11122497
Luo J, Liu S, Hou J, Chen L, Li H, Liao S, Tan Q, Yang T, Yi G, Zhang F, et al. The Comparison of Juvenile Hormone and Transcriptional Changes between Three Different Juvenile Hormone Analogs Insecticides on Honey Bee Worker Larval’s Development. Agronomy. 2021; 11(12):2497. https://doi.org/10.3390/agronomy11122497
Chicago/Turabian StyleLuo, Jie, Sheng Liu, Jiangan Hou, Lichao Chen, Honghong Li, Shucheng Liao, Qimei Tan, Taiyi Yang, Guoqiang Yi, Fu Zhang, and et al. 2021. "The Comparison of Juvenile Hormone and Transcriptional Changes between Three Different Juvenile Hormone Analogs Insecticides on Honey Bee Worker Larval’s Development" Agronomy 11, no. 12: 2497. https://doi.org/10.3390/agronomy11122497
APA StyleLuo, J., Liu, S., Hou, J., Chen, L., Li, H., Liao, S., Tan, Q., Yang, T., Yi, G., Zhang, F., & Li, X. (2021). The Comparison of Juvenile Hormone and Transcriptional Changes between Three Different Juvenile Hormone Analogs Insecticides on Honey Bee Worker Larval’s Development. Agronomy, 11(12), 2497. https://doi.org/10.3390/agronomy11122497