
Chemical Components and Biological Activities of Essential Oils of Mentha × piperita L. from Field-Grown and Field-Acclimated after In Vitro Propagation Plants

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Micropropagation
2.3. Isolation of the Essential Oil and Determination of Essential Oil Composition
2.4. Determination of the Antioxidant Activities of the Essential Oils
2.5. Statistical Analysis
3. Results and Discussion
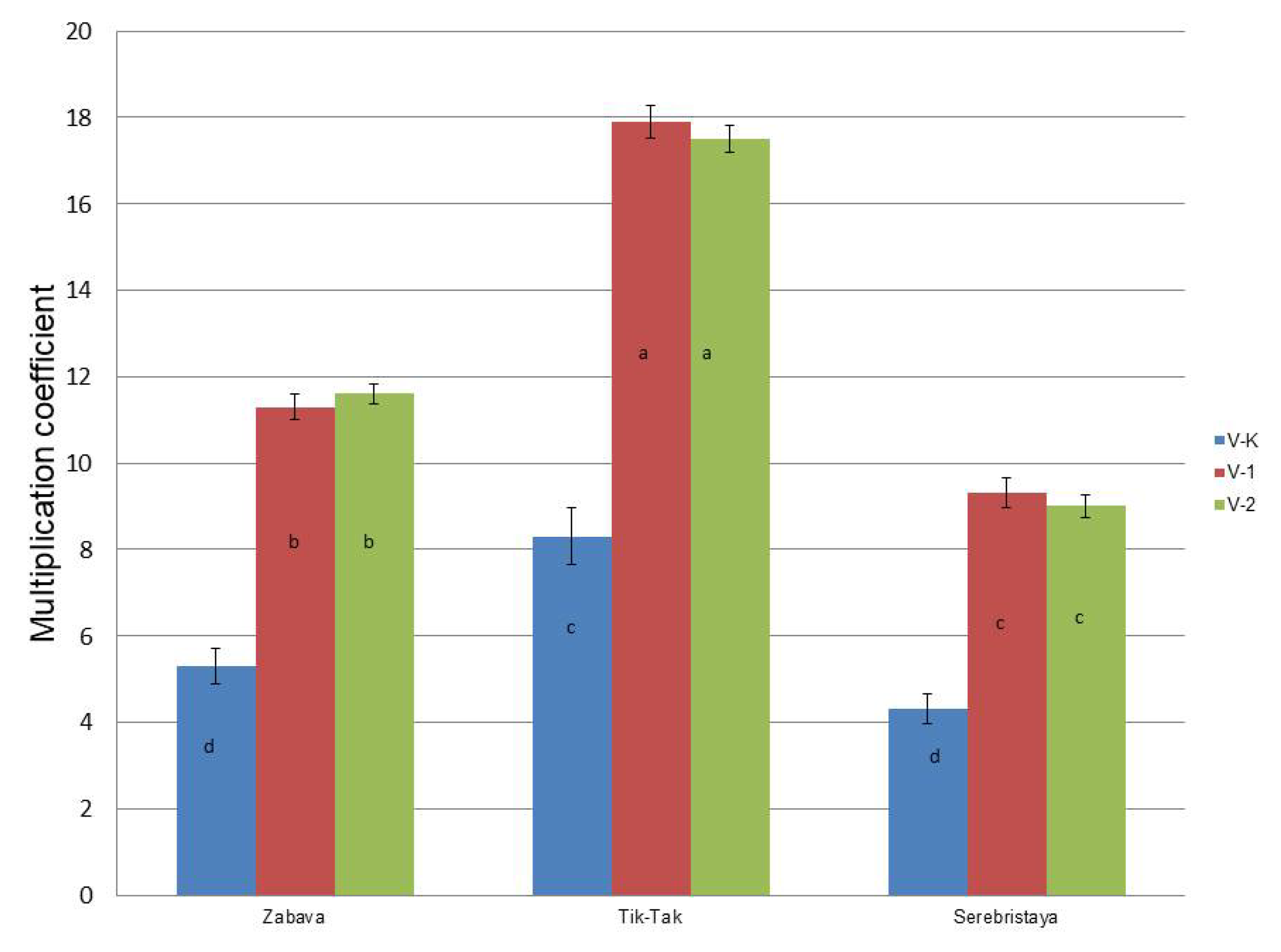
3.1. In Vitro Cultures
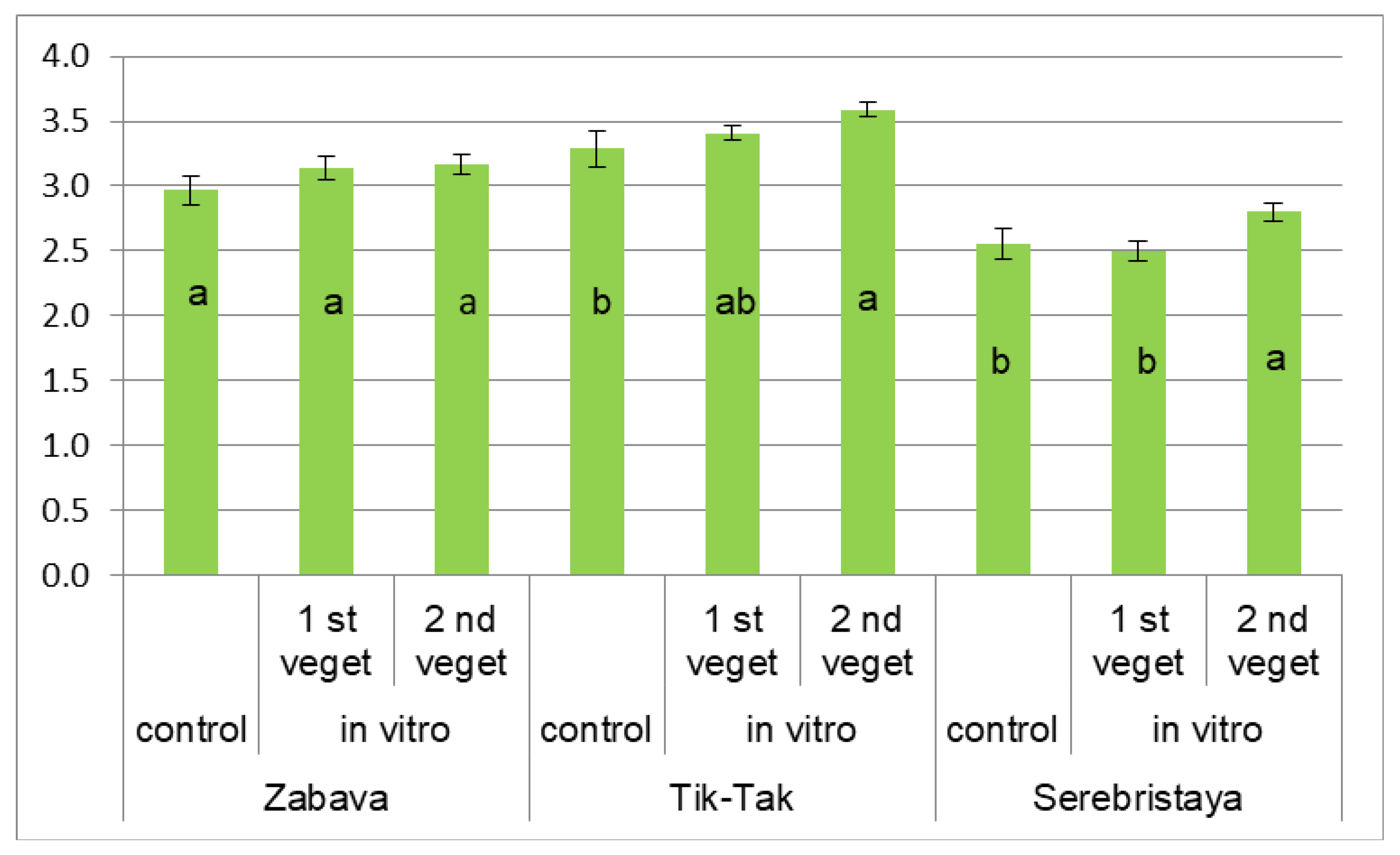
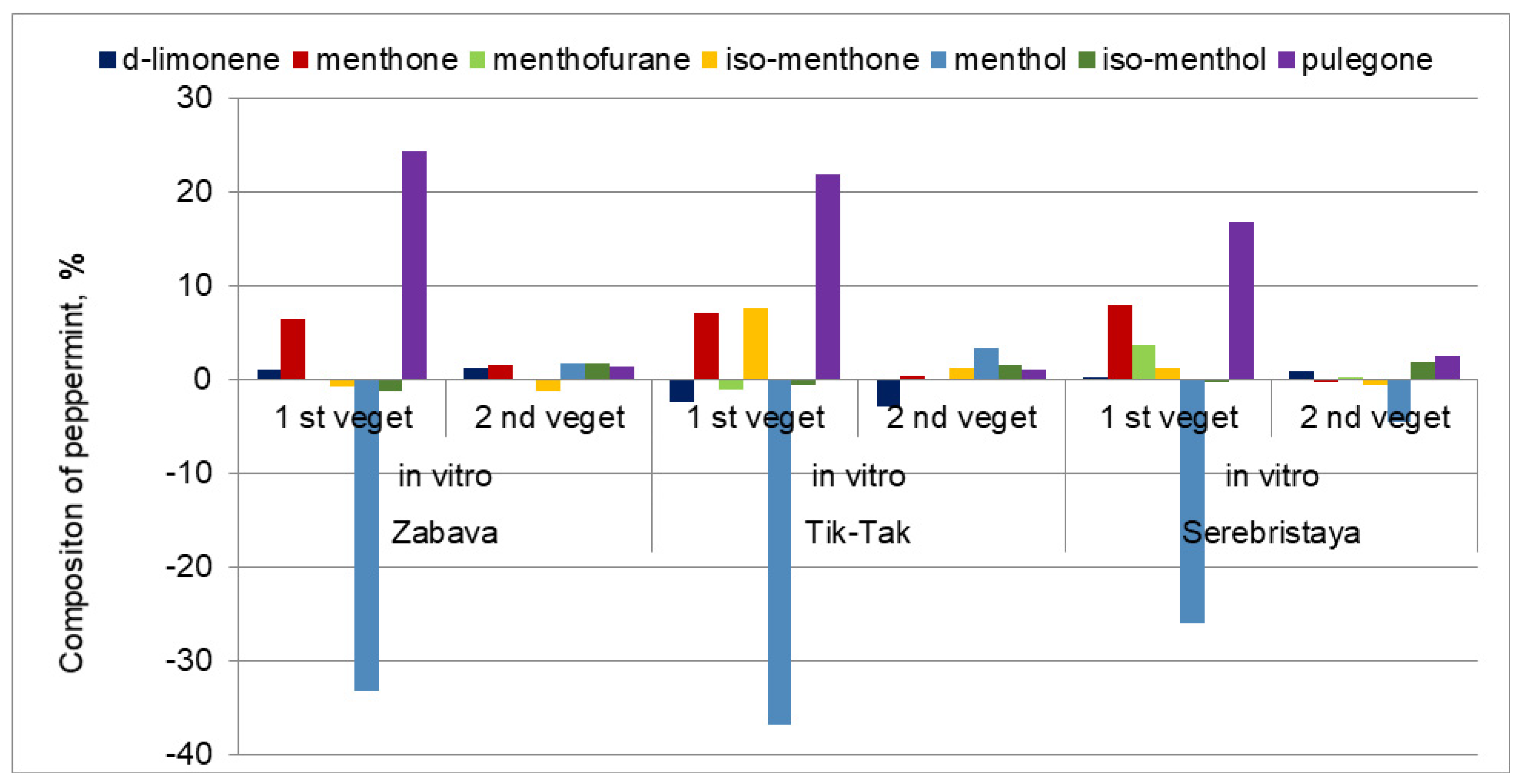
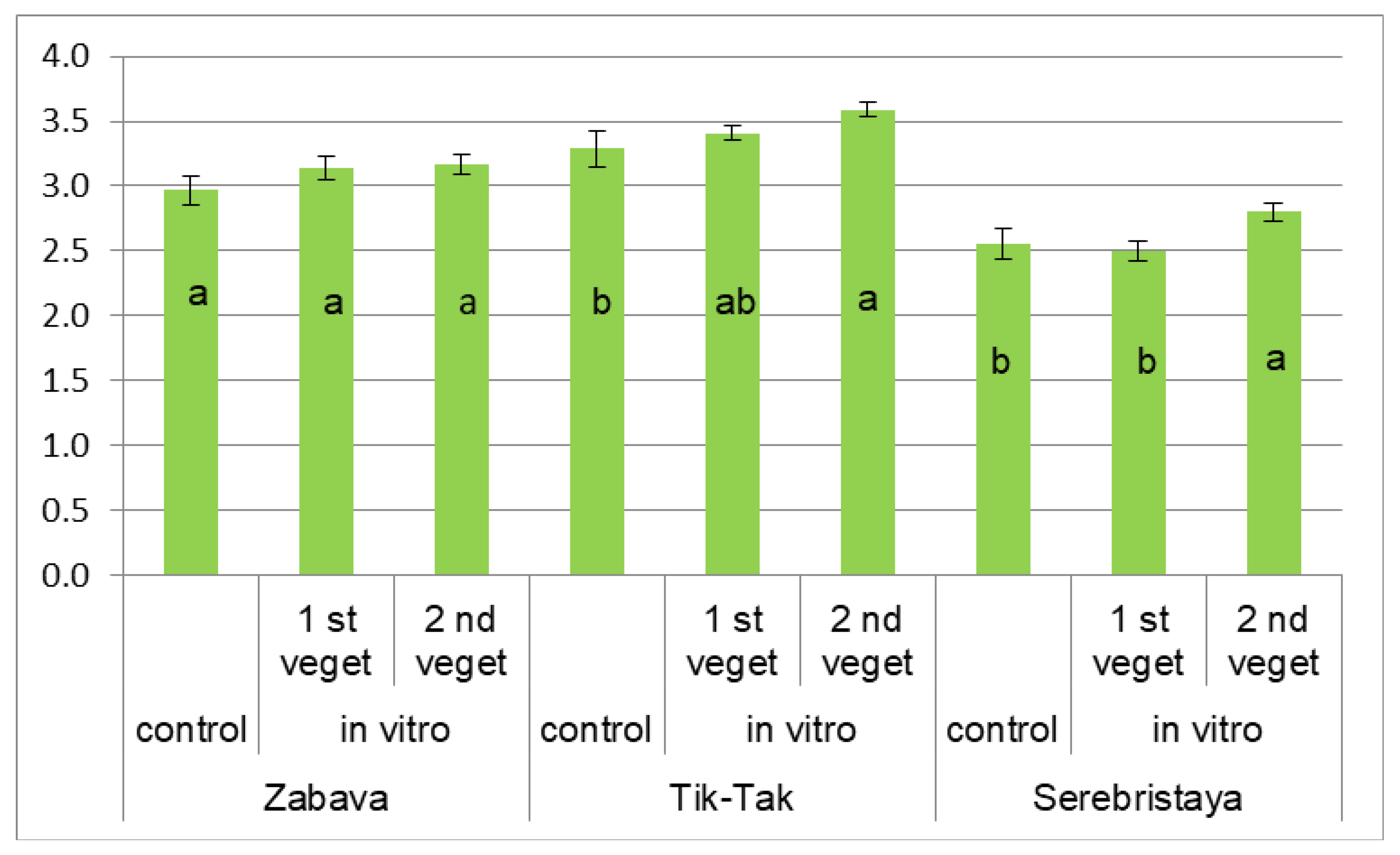
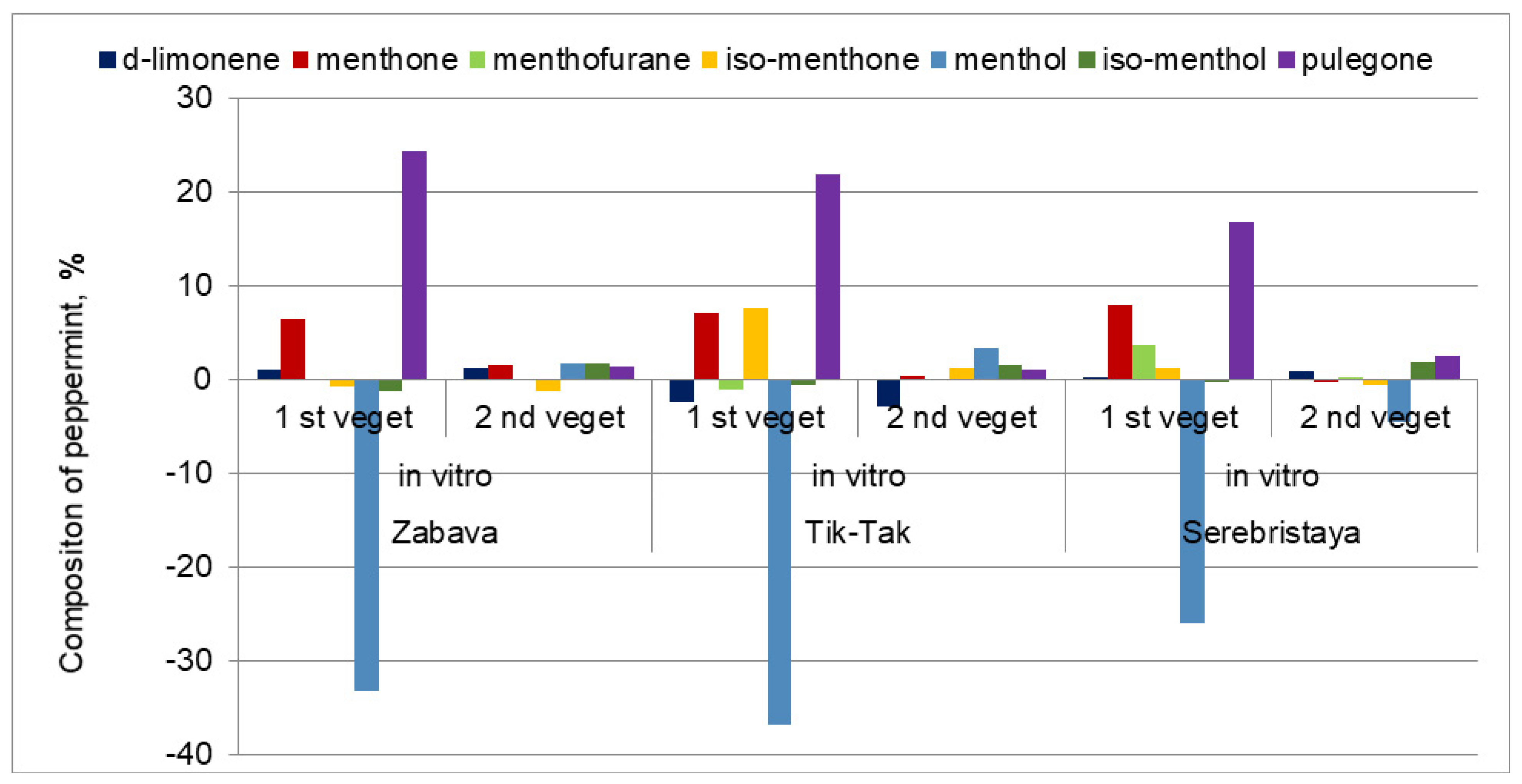
3.2. Peppermint’s Essential Oil Composition
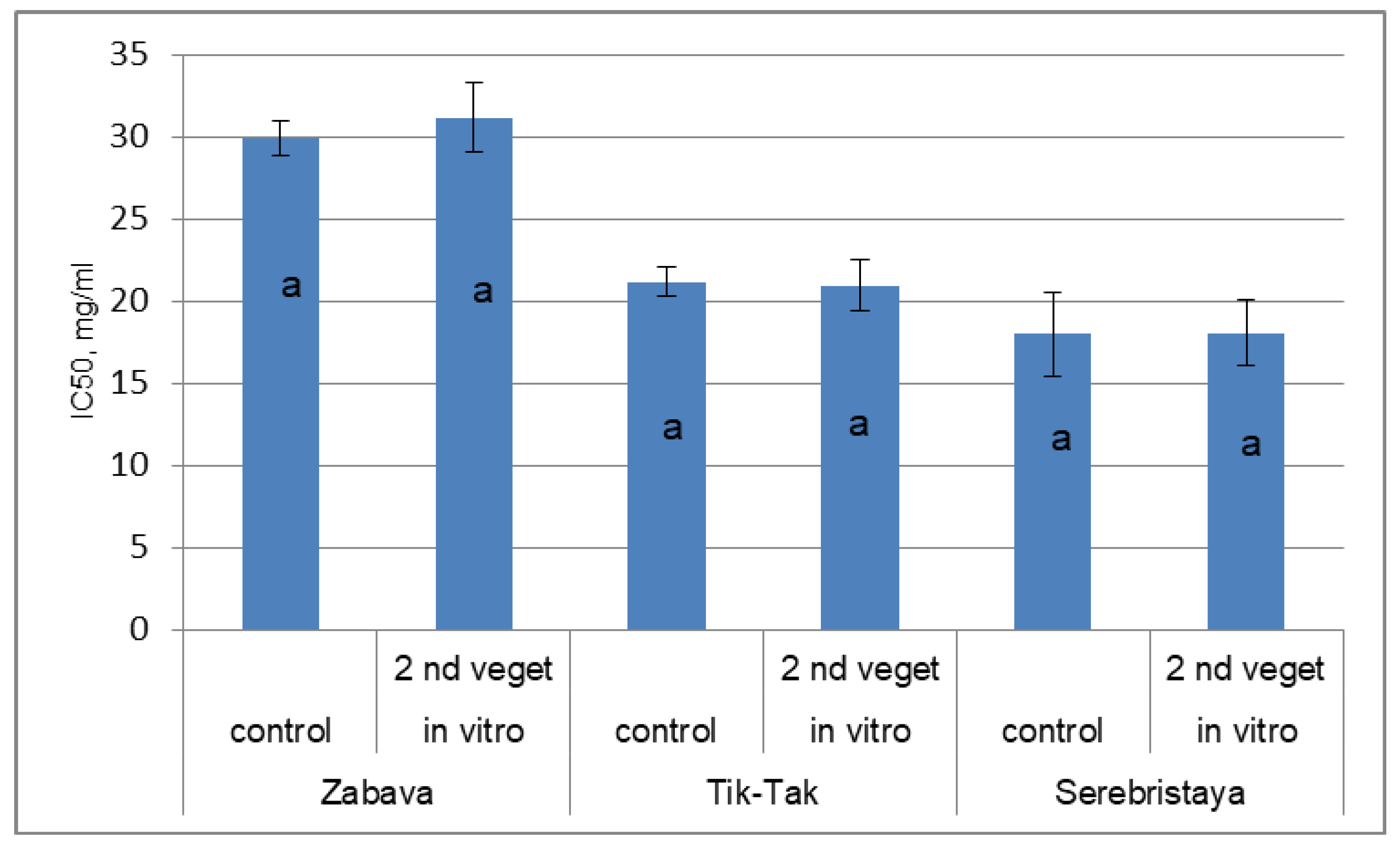
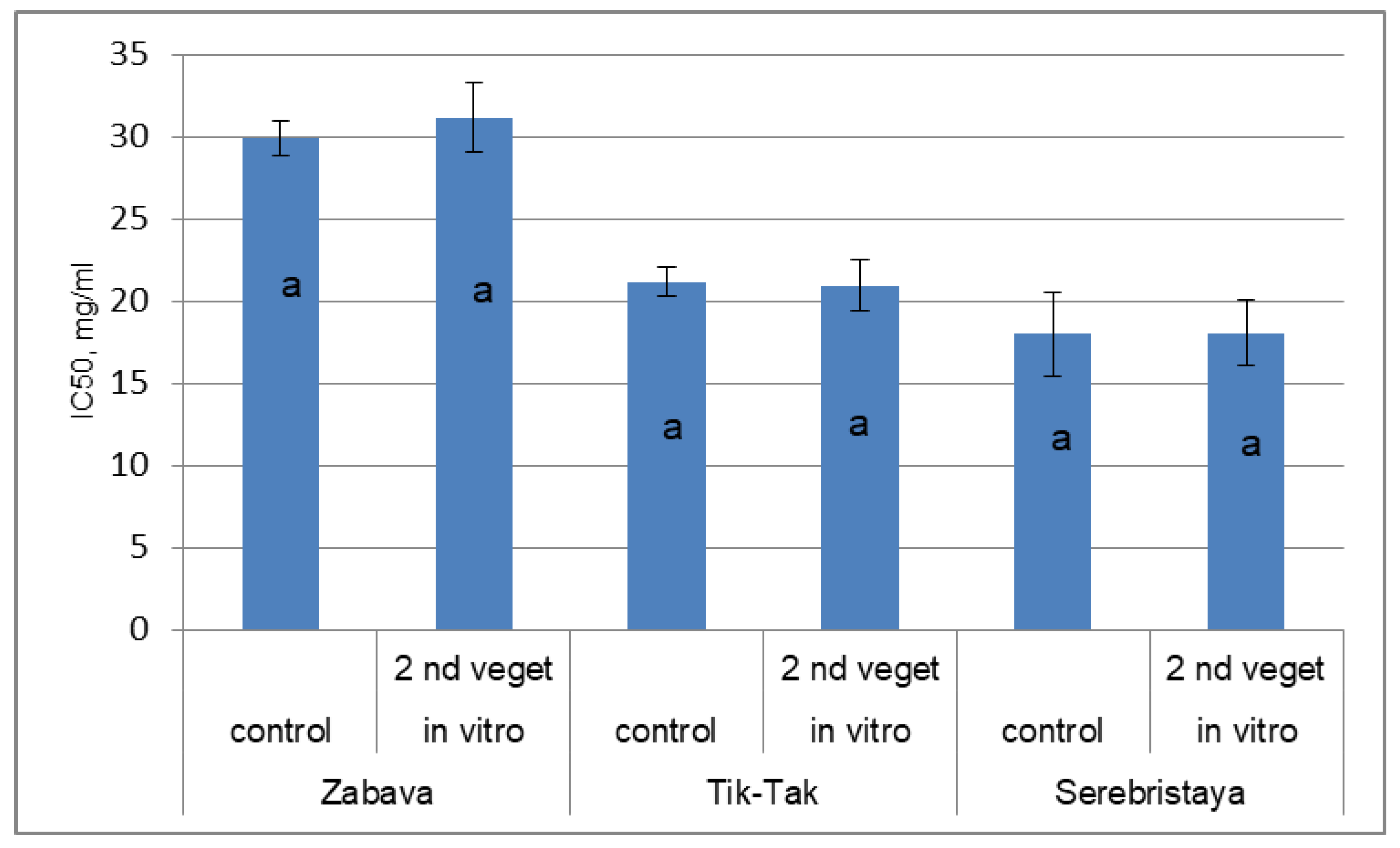
3.3. Antioxidant Activity of Peppermint Essential Oil
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Foster, S. Peppermint: Mentha× piperita. In Botantical Series; American Botanical Council: Austin, TX, USA, 1996; pp. 3–8. [Google Scholar]
- Trevisan, S.C.C.; Menezes, A.P.P.; Barbalho, S.M.; Guiguer, E.L. Properties of Mentha piperita: A brief review. World J. Pharm. Med. Res. 2017, 3, 309–313. [Google Scholar]
- Silva, H. A descriptive overview of the medical uses given to Mentha aromatic herbs throughout history. Biology 2020, 9, 484. [Google Scholar] [PubMed]
- Herro, E.; Jacob, S.E. Mentha piperita (Peppermint). Dermatitis 2010, 21, 327–329. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, A. Therapeutic uses of peppermint—A review. J. Pharm. Sci. Res. 2015, 7, 474–476. [Google Scholar]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential oils as antimicrobial agents—Myth or real alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef] [Green Version]
- Mckay, D.L.; Blumberg, J.B. A review of the bioactivity and potential health benefits of peppermint tea (Mentha piperita L.). Phythother. Res. 2006, 20, 619–633. [Google Scholar] [CrossRef]
- Salehi, B.; Stojanovic-Radic, Z.; Matejic, J.; Sharopov, F.; Antola, H.; Kręgiel, D.; Sen, S.; Sharifi-Rad, M.; Acharya, K.; Sharifi-Rad, R.; et al. Plants of genus Mentha: From farm to food factory. Plants 2018, 7, 70. [Google Scholar] [CrossRef] [Green Version]
- Amato, A.; Liotta, R.; Mule, F. Effects of menthol on circular smooth muscle of human colon: Analysis of the mechanism of action. Eur. J. Pharmacol. 2014, 740, 295–301. [Google Scholar] [CrossRef]
- Nair, B. Final report on the safety assessment of Mentha piperita (peppermint) oil, Mentha piperita (peppermint) leaf extract, Mentha piperita (peppermint) leaf, and Mentha piperita (peppermint) leaf water. Int. J. Toxicol. 2001, 20, 61–73. [Google Scholar]
- Saric-Kundalic, B.; Fialova, S.; Dobes, C.; Olzant, S.; Tekelova, D.; Grancai, D.; Reznicek, G.; Saukel, J. Multivariate numerical taxonomy of Mentha species, hybrids, varieties and cultivars. Sci. Pharm. 2009, 77, 851–876. [Google Scholar] [CrossRef]
- Ludwiczuk, A.; Kiełtyka-Dadasiewicz, A.; Sawicki, R.; Golus, J.; Ginalska, G. Essential oils of some Mentha species and cultivars, their chemistry and bacteriostatic activity. Nat. Prod. Commun. 2016, 11, 1018. [Google Scholar] [CrossRef] [Green Version]
- Krasnyanski, S.; Ball, T.M.; Sink, K.C. Somatic hybridization in mint: Identification and characterization of Mentha piperita (+) M. spicata hybrid plants. Theor. Appl. Genet. 1998, 96, 683–687. [Google Scholar] [CrossRef]
- Kalemba, D.; Synowiec, A. Agrobiological interactions of essential oils of two menthol mints: Mentha piperita and Mentha arvensis. Molecules 2020, 25, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelepova, O.V.; Olekhnovich, L.S.; Konovalova, L.N.; Khusnetdinova, T.I.; Gulevich, A.A.; Baranova, E.N. Assessment of essential oil yield in three mint species in the climatic conditions of Central Russia. Agron. Res. 2021, 19, 1551–1562. [Google Scholar] [CrossRef]
- Chaput, M.-H.; San, H.; De Hys, L.; Grenier, E.; David, H.; David, A. How Plant Regeneration from Mentha× piperita L. and Mentha× citrata Ehrh. leaf protoplasts affects their monoterpene composition in field conditions. J. Plant Physiol. 1996, 149, 481–488. [Google Scholar] [CrossRef]
- Mehta, J.; Naruka, R.; Sain, M.; Dwivedi, A.; Sharma, D.; Mirza, J. An efficient protocol for clonal micropropagation of Mentha piperita L. (peppermint). Asian J. Plant. Sci. Res. 2012, 2, 518–523. [Google Scholar]
- Talankova-Sereda, T.E.; Kolomiets, J.V.; Holubenko, A.V.; Nuzhyna, N.V. The influence of clonal micropropagation on productivity and differentiation of Mentha piperita plant tissues. Regul. Mech. Biosyst. 2019, 10, 337–344. [Google Scholar] [CrossRef]
- Shkopynska, T.Y.; Kolomiets, Y.V.; Hryhoriuk, I.P.; Kucenko, N.I. Economic valuable traits of promising breeding samples and ‘Chornolysta’ variety of Mentha piperita L. after on vitro sanitation and micropropagation. Plant Var. Stud. Protect. 2019, 15, 424–433. [Google Scholar]
- Bertoli, A.; Leonardi, M.; Krzyzanowska, J.; Oleszek, W.; Pistelli, L. In vitro production of M. × piperita not containing pulegone and menthofuran. Acta Biochim. Pol. 2012, 59, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Fejér, J.; Gruľová, D.; De Feo, V.; Ürgeová, E.; Obert, B.; Preťová, A. Mentha× piperita L. nodal segments cultures and their essential oil production. Ind. Crops Products 2018, 112, 550–555. [Google Scholar] [CrossRef]
- Łyczko, J.; Piotrowski, K.; Kolasa, K.; Galek, R.; Szumny, A. Mentha piperita L. micropropagation and the potential influence of plant growth regulators on volatile organic compound composition. Molecules 2020, 25, 2652. [Google Scholar] [CrossRef] [PubMed]
- Zayova, E.; Kirova, E.; Geneva, M. Optimized cultural conditions for rapid in vitro propagation and conservation of Mentha piperita L. Comptes Rendus Acad. Bulg. Sci. 2021, 74, 945–954. [Google Scholar]
- Khan, S.; Shende, S.M.; Bonde, D.R. In vitro micropropagation of mint (Mentha). World J. Pharm. Pharmaceut. Sci. 2021, 10, 1688–1692. [Google Scholar]
- Prins, C.L.; Vieira, I.J.C.; Freitas, S.P. Growth regulators and essential oil production. Brazil. Soc. Plant Physiol. 2010, 22, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Khanam, D.; Mohammad, F. Effect of structurally different plant growth regulators (PGRs) on the concentration, yield, and constituents of peppermint essential oil. J. Herbs. Spices Med. Plants 2018, 23, 26–35. [Google Scholar] [CrossRef]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Ekiert, H.; Klimek-Szczykutowicz, M.; El-Ansary, D.O.; Mahmoud, E.A. Polyphenol profile and antimicrobial and cytotoxic activities of natural Mentha× piperita and Mentha longifolia populations in Nothern Saudi Arabia. Processes 2020, 8, 479. [Google Scholar] [CrossRef] [Green Version]
- Malankina, E.L.; Kozlovskaya, L.N.; Tkacheva, E.N. Epidermal structures of leaves in some Mentha × piperita L. varieties in connection with their productivity. Veget. Crops Russ. 2019, 6, 67–71. (In Russian) [Google Scholar] [CrossRef] [Green Version]
- Bochkarev, N.I.; Zelencov, S.V.; Shuvaeva, T.P.; Borodkina, A.P. Taxonomy, morphology and breeding of menthol mints (review). Nauchno-Teh. Byulleten Vserossiiskogo Nauchno-Issledovatelskogo Inst. Maslichnih Kult. 2015, 2, 106–124. (In Russian) [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio-assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Zagorskaya, M.S.; Egorova, N.A. Optimizaciya sostava pitatel’noj sredy dlya klonal’nogo mikrorazmnozheniya sortov myaty Azhurnaya and Bergamotnaya. Biol. Chem. 2018, 4, 73–84. [Google Scholar]
- Niemi, M.; Vestberg, M. Inoculation of commercially grown strawberry with VA mycorrhizal fungi. Plant Soil 1992, 144, 133–142. [Google Scholar] [CrossRef]
- European Pharmacopoeia, 7th ed; Edition 2.8.18; Council of Europe: Strasbourg, France, 2010; Volume 1, p. 284.
- Shelepova, O.V.; Semenova, M.V.; Enina, O.L.; Schanzer, I.A. Genetic, phenotypic and phytochemical polymorphism in Eastern European populations of Mentha arvensis L. Russ. J. Gen. 2017, 1, 59–66. [Google Scholar] [CrossRef]
- McLafferty, F.W. Wiley Registry of Mass Spectral Data 7th Edition with NIST 2005 Spectral Data; John Wiley and Sons: New York, NY, USA, 2005. [Google Scholar]
- Brand, W.; Cuvelier, W.; Berset, M.E. Use of a free radical method to evaluate antioxydant activity. Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Vasile, L.; Maria, Z.; Simona, V.; Eliza, A. Use of nodal explants in “in vitro” micro-propagation of Mentha piperita L. Fasc. Protecţia Mediu. 2011, 16, 247–251. [Google Scholar]
- Santoro, M.V.; Nievas, F.; Zygadlo, J.; Giordano, W.; Banchio, E. Effects of growth regulators on biomass and the production of secondary metabolites in peppermint (Mentha piperita) micropropagated in vitro. Am. J. Plant Sci. 2013, 4, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Islam, A.T.M.R.; Islam, M.; Alam, M.F. Rapid in vitro clonal propagation of herbal spice, Mentha piperita L. using shoot tip and nodal explants. Res. Plant Sci. 2017, 5, 43–50. [Google Scholar]
- Akter, K.T.; Hoque, M.A.; Yeasmin, T. Effect of IBA Concentration on in vitro root regeneration of different mint explants. Ann. Bangladesh Agric. 2016, 20, 49–59. [Google Scholar]
- Zuzarte, M.R.; Dinis, A.M.; Cavaleiro, C.; Salgueiro, L.R.; Canhoto, J.M. Trichomes, essential oils and in vitro propagation of Lavandula pedunculata (Lamiaceae). Ind. Crops Prod. 2010, 32, 580–587. [Google Scholar] [CrossRef]
- Benahmed, A.; Harfi, B.; Belkhiri, A. Biological activity of essential oils of Mentha pulegium from field-grown and acclimated in vitro plants. Curr. Sci. 2019, 116, 1897–1904. [Google Scholar] [CrossRef]
- Chekurova, G.V.; Evtukhova, L.A.; Slyusarenko, A.G. Reproduction of cranberry in vitro culture. Bull. Main Bot. Gard. 1990, 157, 90–95. (In Russian) [Google Scholar]
- Heck, M.A.; Lüth, V.M.; van Gessel, N.; Krebs, M.; Kohl, M.; Prager, A.; Joosten, H.; Decker, E.L.; Reski, R. Axenic in vitro cultivation of 19 peat moss (Sphagnum L.) species as a resource for basic biology, biotechnology, and paludiculture. New Phytol. 2021, 229, 861–876. [Google Scholar] [CrossRef] [PubMed]
- The Federal Electronic Medical Library. The State Pharmacopoeia of the Russian Federation, 13th ed.; Moscow, Russia; 2015; Volumes 1–3. Available online: http://femb.ru/femb/pharmacopea13.php (accessed on 18 October 2021). (In Russian)
- Baser, K.; Husnu, C.; Gerhard, B. Handbook of Essential Oils. Science, Technology, and Applications, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016; p. 1128. [Google Scholar]
- Shelepova, O.V.; Kondrat’eva, V.V.; Olehnovich, L.S.; Zaithik, B.T.; Ruzhitsky, A.O. Variations in composition of essential oil and in accumulation of salicylic acid in Mentha arvensis var. piperascens (Lamiaceae) when introduced in Moscow region. Rastit. Resur. 2016, 3, 94–104. (In Russian) [Google Scholar]
- Shigeto, A.; Wada, A.; Kumazawa, K. Identification of the novel odor active compounds “p-menthane lactones” responsible for the characteristic aroma of fresh peppermint leaf. Biosci. Biotechnol. Biochem. 2020, 84, 421–427. [Google Scholar] [CrossRef] [PubMed]
- da Ramos, R.S.; Rodrigues, A.B.L.; Farias, A.L.F.; Simões, R.C.; Pinheiro, M.T.; dos Ferreira, R.M.A.; Barbosa, L.M.C.; Souto, R.N.P.; Fernandes, J.B.; Lourivaldo da Silva Santos, L.S.; et al. Chemical composition and in vitro antioxidant, cytotoxic, antimicrobial, and larvicidal activities of the essential oil of Mentha piperita L. (Lamiaceae). Sci. World J. 2017, e 4927214. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemotype | No of New Lateral Shoots | Plant Height (cm) | No of New Nodal Segments | Root Length (cm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V-K | V-1 | V-2 | V-K | V-1 | V-2 | V-K | V-1 | V-2 | V-K | V-1 | V-2 | |
| Zabava | 1.3 ± 0.1 c | 2.7 ± 0.1 b | 3.3 ± 0.2 a | 5.0 ± 0.2 a | 4.5 ± 0.1 b | 4.1 ± 0.3 bc | 4.1 ± 0.4 a | 4.2 ± 0.5 a | 3.5 ± 0.3 b | 2.1 ± 0.1 b | 3.5 ± 0.2 a | 1.5 ± 0.2 c |
| Tik-Tak | 1.5 ± 0.2 c | 3.2 ± 0.4 ab | 3.9 ± 0.2 a | 6.5 ± 0.2 a | 5.8 ± 0.2 b | 5.1 ± 0.4 c | 5.5 ± 0.3 ab | 5.9 ± 0.4 a | 4.5 ± 0.6 b | 4.8 ± 0.3 b | 6.5 ± 0.3 a | 4.2 ± 0.2 bc |
| Serebristaya | 1.1 ± 0.1 b | 2.5 ± 0.3 a | 2.8 ± 0.2 a | 4.3 ± 0.1 a | 3.9 ± 0.2 bc | 3.3 ± 0.3 c | 3.9 ± 0.2 a | 3.7 ± 0.5 a | 3.1 ± 0.3 b | 2.6 ± 0.2 ab | 3.0 ± 01 a | 1.8 ± 0.1 c |
| Compound | Zabava | Tik-Tak | Serebristaya | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | In vitro | Control | In vitro | Control | In vitro | ||||
| 1st Vegetation Year | 2nd VegetationYear | 1st Vegetation Year | 2nd Vegetation Year | 1st Vegetation Year | 2nd Vegetation Year | ||||
| α-pinene | 0.7 ab | 0.8 a | 0.6 b | 0.6 a | 0.2 c | 0.4 b | 0.9 ab | 1.0 a | 0.8 b |
| sabinene | 0.3 ab | 0.3 ab | 0.4 a | 0.3 a | 0.2 ab | 0.2 ab | 0.6 a | 0.6 a | 0.5 ab |
| β-pinene | 0.7 a | 0.7 a | 0.8 a | 0.6 a | 0.3 b | 0.3 b | 1.2 a | 1.2 a | 1.0 b |
| myrcene | 0.5 ab | 0.6 a | 0.4 b | 0.4 a | 0.3 a | 0.3 a | 0.3 a | 0.2 a | 0.2 a |
| 3-octanol | 0.5 a | 0.4 a | 0.4 a | 0.6 a | 0.3 b | 0.5 a | 0.3 b | 0.2 bc | 0.6 a |
| α-terpinene | 0.1 a | 0.2 a | 0.2 a | 0.1 a | 0.1 a | 0.1 a | 0.3 a | 0.2 a | 0.2 a |
| ρ-cymene | 0.1 a | 0.1 a | 0.1 a | 0.1 a | t | 0.1 a | 0.4 ab | 0.1 | 0.5 a |
| d-limonene | 1.5 c | 2.6 b | 2.8 a | 4.0 a | 1.7 b | 1.1 bc | 2.1 bc | 2.3 b | 3.0 a |
| 1,8-cineole | 1.1 b | 1.2 b | 2.2 a | 1.2 c | 1.8 b | 2.3 a | 6.5 a | 5.5 b | 5.1 bc |
| trans-ocimene | 0.1 a | 0.1 a | t | 0.1 b | 0.3 a | 0.1 b | 0.2 a | 0.2 a | 0.1 a |
| γ-terpinene | 0.1 a | 0.2 a | 0.1 a | 0.1 a | 0.1 a | 0.1 a | 0.6 a | 0.7 a | 0.5 b |
| trans-sabinene hydrate | 0.2 a | 0.3 a | 0.3 a | t | t | t | 0.4 a | 0.1 | 0.3 ab |
| linalool | 0.1 a | 0.2 a | 0.1 a | 0.1 a | 0.1 a | 0.1 a | t | t | t |
| menthone | 8.3 c | 14.4 a | 9.9 b | 10.8 c | 18.0 a | 11.2 b | 27.1 c | 35.0 a | 26.8 b |
| menthofuran | 0.1 a | 0.1 a | t | t | 1.0 | t | 3.6 c | 7.3 a | 4.9 b |
| iso-menthone | 3.2 a | 2.4 b | 2.0 bc | 4.0 c | 11.6 a | 5.3 b | 3.2 b | 4.4 a | 2.7 c |
| neo-menthol | 0.1 a | 0.1 a | 0.2 a | 0.2 bc | 0.5 a | 0.3 b | 0.7 a | 0.4 bc | 0.5 b |
| menthol | 56.5 a | 23.3 d | 51.3 b | 66.6 a | 29.8 d | 59.9 b | 36.6 a | 10.6 d | 32.1 b |
| iso-menthol | 3.7 b | 2.4 c | 5.5 a | 1.5 b | 1.0 c | 3.1 a | 2.5 b | 2.3 bc | 4.4 a |
| α-terpineol | 0.1 b | 0.3 a | 0.1 b | 0.1 a | 0.2 a | 0.2 a | 0.3 a | 0.3 a | 0.4 a |
| pulegone | 1.0 d | 25.3 a | 2.4 c | 0.7 d | 22.5 a | 1.8 c | 3.1 d | 19.9 a | 5.7 c |
| carvone | 0.2 a | 0.1 a | 0.1 a | 0.1 a | 0.1 a | t | 0.1 c | 0.9 a | 0.3 b |
| p-menthane-8-ol | t | t | t | t | t | t | 0.1 ab | 0.2 a | 0.2 a |
| piperitone | 2.9 d | 4.4 a | 3.5 c | 0.1 c | 1.0 a | 1.1 a | 0.5 bc | 1.6 a | 0.7 b |
| Menthyl acetate | 5.8 a | 3.3 bc | 3.9 b | 0.1 d | 6.7 a | 1.2 c | 0.1 a | 0.1 a | 0.1 a |
| β-caryophyllene | 0.1 b | t | 0.3 a | 0.2 b | 0.4 a | 0.4 a | t | t | t |
| germacrene D | 0.2 a | 0.2 a | 0.2 a | 0.8 a | t | 0.7 a | t | t | t |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shelepova, O.V.; Dilovarova, T.A.; Gulevich, A.A.; Olekhnovich, L.S.; Shirokova, A.V.; Ushakova, I.T.; Zhuravleva, E.V.; Konovalova, L.N.; Baranova, E.N. Chemical Components and Biological Activities of Essential Oils of Mentha × piperita L. from Field-Grown and Field-Acclimated after In Vitro Propagation Plants. Agronomy 2021, 11, 2314. https://doi.org/10.3390/agronomy11112314
Shelepova OV, Dilovarova TA, Gulevich AA, Olekhnovich LS, Shirokova AV, Ushakova IT, Zhuravleva EV, Konovalova LN, Baranova EN. Chemical Components and Biological Activities of Essential Oils of Mentha × piperita L. from Field-Grown and Field-Acclimated after In Vitro Propagation Plants. Agronomy. 2021; 11(11):2314. https://doi.org/10.3390/agronomy11112314
Chicago/Turabian StyleShelepova, Olga V., Tatyana A. Dilovarova, Alexander A. Gulevich, Ludmila S. Olekhnovich, Anna V. Shirokova, Irina T. Ushakova, Ekaterina V. Zhuravleva, Ludmila N. Konovalova, and Ekaterina N. Baranova. 2021. "Chemical Components and Biological Activities of Essential Oils of Mentha × piperita L. from Field-Grown and Field-Acclimated after In Vitro Propagation Plants" Agronomy 11, no. 11: 2314. https://doi.org/10.3390/agronomy11112314
APA StyleShelepova, O. V., Dilovarova, T. A., Gulevich, A. A., Olekhnovich, L. S., Shirokova, A. V., Ushakova, I. T., Zhuravleva, E. V., Konovalova, L. N., & Baranova, E. N. (2021). Chemical Components and Biological Activities of Essential Oils of Mentha × piperita L. from Field-Grown and Field-Acclimated after In Vitro Propagation Plants. Agronomy, 11(11), 2314. https://doi.org/10.3390/agronomy11112314