1. Introduction
The accurate estimation of leaf photosynthetic pigments concentration is crucial to check the plant´s health, particularly in agricultural systems, where growth and quality are directly related to plant status [
1].
Chlorophylls (Chl) are the most important photosynthetic pigments since they are responsible for the harvesting of light energy, transferring excitation energy to reaction centers, and driving charge separation reactions in reaction centers [
2]. Chlorophyll a is present in the light harvesting complexes, photosystems (I and II) while chlorophyll b is necessary for stabilizing the major light-harvesting chlorophyll binding proteins [
3,
4]. Carotenoids (Ct) also participate in harvesting light energy for photosynthesis [
5]. In addition, they are also involved in the defense mechanism against oxidative stress [
6] and play an essential role in the dissipation of excess light energy and provide protection to reaction centers. As a photo-protection mechanism, they are retained during the process of chlorophyll degeneration at leaf senescence [
7].
Traditional methods of measuring photosynthetic pigments involve complex procedures of solvent extraction followed by spectrophotometric determinations, which make them destructive, labor intensive, time-consuming and expensive [
8].
During recent years, portable plant instruments such as Soil Plant Analysis Development (SPAD) meters have become increasingly important to monitor plant health conditions in a non-destructive way. These devices offer a modest, fast and non-destructive approach to determine relative values of total chlorophyll content, but the main drawback of these devices is the need of calibration for measurement in absolute units of chlorophyll concentration per unit of leaf area [
9].
The relationships between SPAD readings and extractable leaf pigments in various plant species have been the focus of several studies in different crops such as soybean [
10], wheat [
11], and tomato [
12]. Nevertheless, it is necessary to point out that the relationship between SPAD values and leaf pigments concentration is not universal and varies with the extraction procedure, sensor type, leaf direction and exposure, and plant species (sometimes even within the same plant species) mainly associated with different leaf optical properties dependent on concentration of light absorbing compounds and the internal scattering of light [
13,
14].
Nowadays, the choice between different species based on the growing period has increased hugely in the ornamental market and the competitiveness at a worldwide level between international growers led to producing a higher variety of saleable plants with higher aesthetic value depending on shape, size, and leaf color [
15]; therefore in ornamental plants it is important to measure chlorophyll by color from a commercial point of view. Although in literature there are some references relating SPAD readings and extractable leaf pigments in ornamental plants [
16,
17], it is nevertheless necessary to obtain species-specific calibration equations for SPAD measurements. Therefore, we have conducted the following experiment to discern the relationship between SPAD values and pigment concentration in several ornamental species of high market value in the Mediterranean area grown under different conditions (greenhouse in spring season and open field in summer season). Moreover, the useful of the regression equations in practice can be the quick determination of chlorophyll concentration and consequently the healthy status of the garden plants in several species based on SPAD readings.
4. Discussion
The chlorophyll content is an important experimental parameter in agronomy and plant biology research [
20]. The amount of chlorophyll can vary among the species and also in the same species being related to the levels of irradiance received, plant nutritional status, and stress conditions as well as depending on the genetic factor [
21].
It should also be considered that there are a lot of methodologies to determine the chlorophyll concentration in leaf tissues in laboratory: N,N-dimethylformamide (DMF) by Rami and Dan [
22]; acetone by Arnon [
23]; dimethylsulfoxide (DMSO) by Houborg et al. [
24] and methanol, and that the used in this work proposed by Wellburn [
19] that offers different values of chlorophyll quantification. In both trials, we used the methodology proposed by Wellburn. This implies that the data of both trials are comparable but should be interpreted if another quantification method has been used.
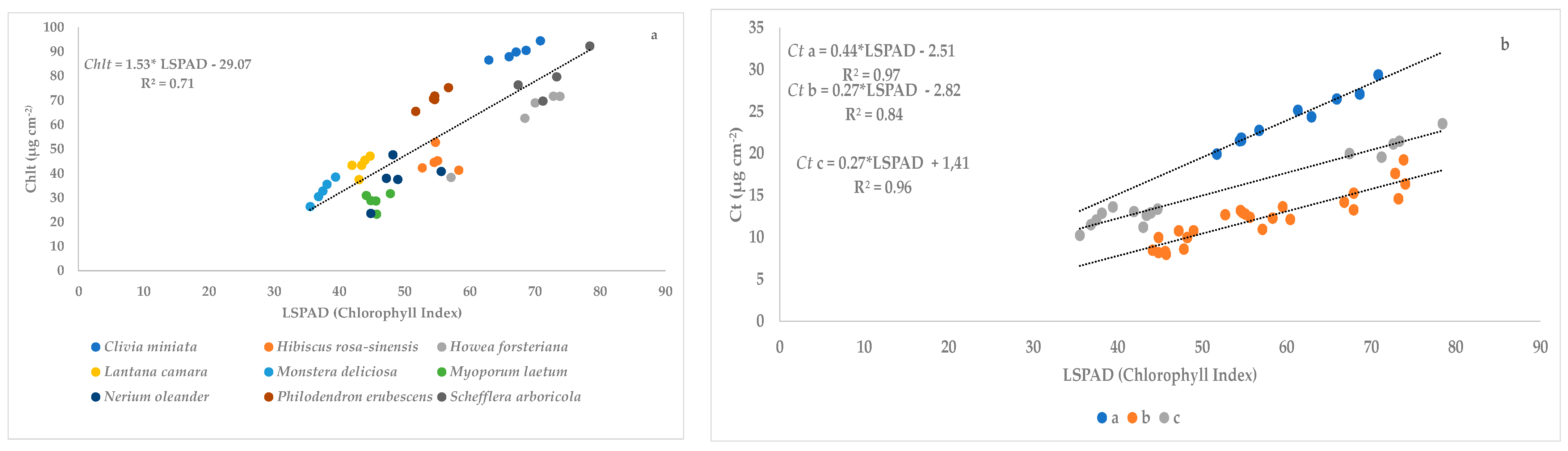
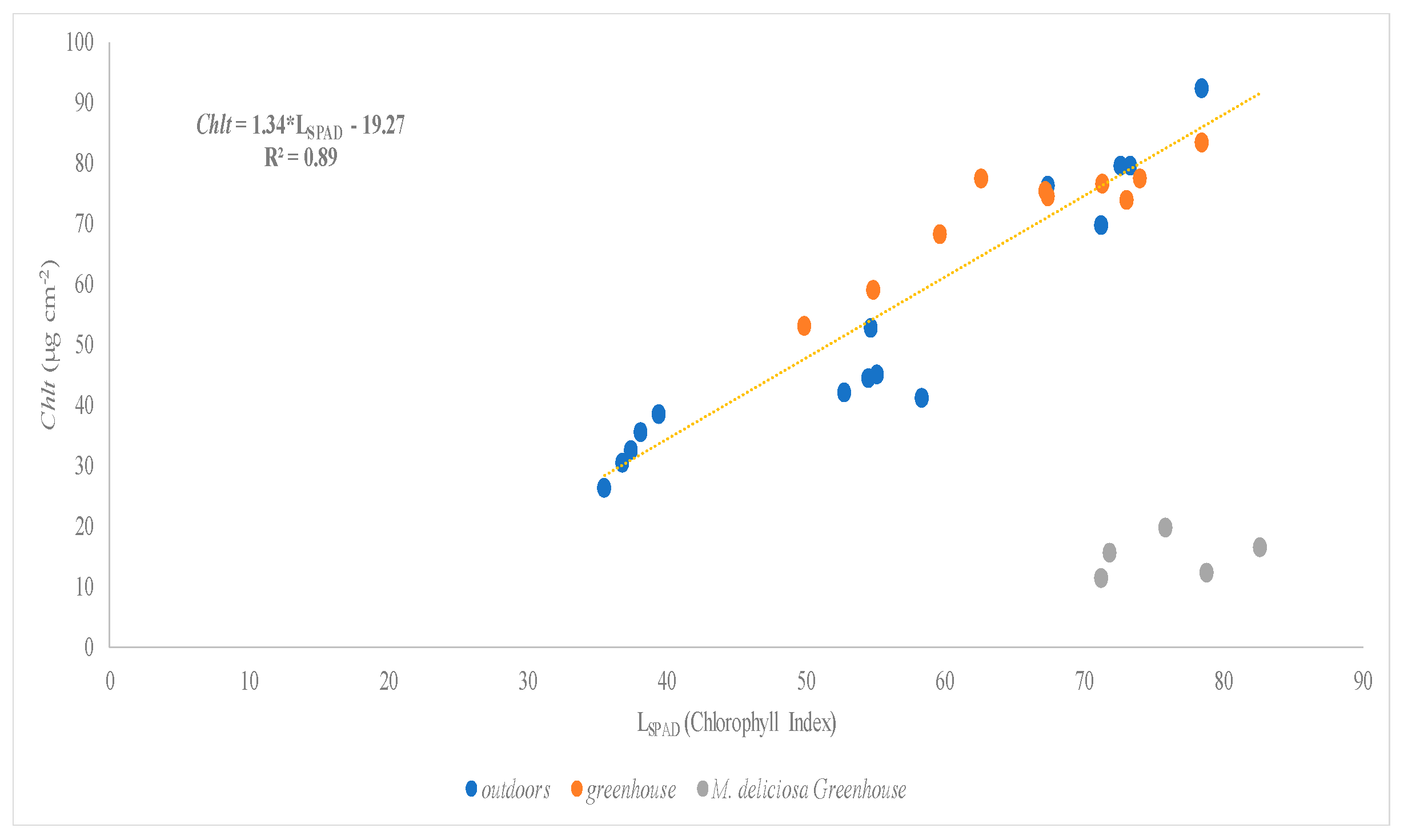
In our experiment, we found that the Chlt concentration varies significantly between species because plant species (sometimes even within the same plant species) have different pigment concentrations [
13]. Comparing the results of both trials, we can see lower values of Chlt in the second trial (summer season) with respect to the first trial (spring season). This fact can be explained by the effects of the high temperature under summer conditions which result in a decrease of photosynthesis rate and consequently in a reduction of pigments concentrations as proposed by Shanmugam et al. [
25]. Moreover, higher levels of irradiance at open field conditions (second trial) may be another possible factor responsible for the pigment reduction a result of photooxidative damage [
26]. Comparing our results obtained with already published data, the Chlt concentration in
C. miniata at field conditions (trial 2) was lower than those reported by Youssef et al. [
27] (2.14 mg g
−1 FW). Our values in
N. oleander were similar to the values proposed by Mugnai et al. [
28]. Lower values were found in
L. camara (10 times less) than those presented by Singh et al. [
29], which can be associated with the use of acetone as extractant. Similar values have been found in
N. exaltata compared to the data obtained by Parminder et al. [
30].
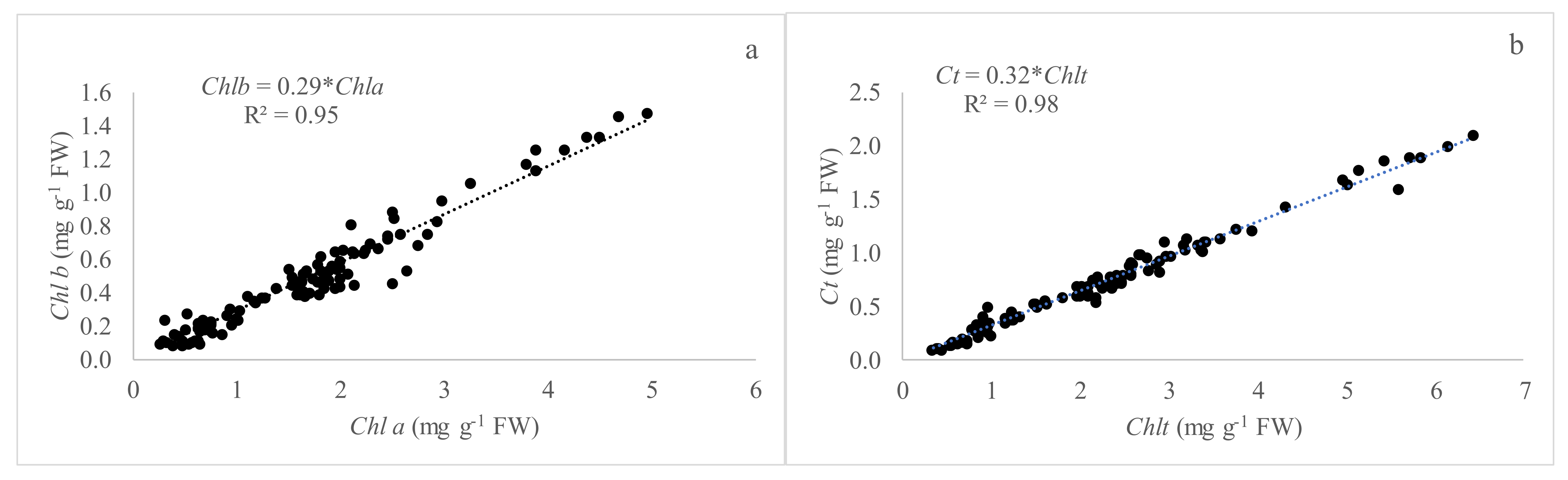
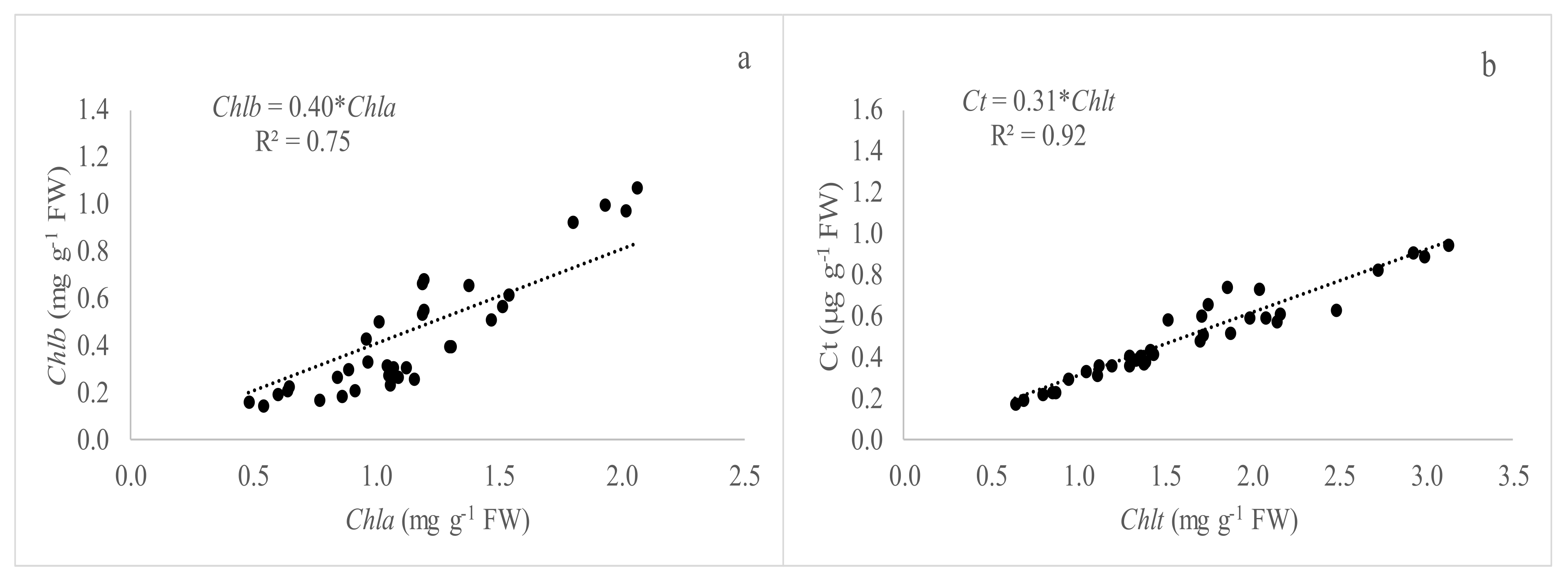
We examined the relationships between Chla, Chlb, Chlt, and Ct content of the species studied. Not surprisingly, Chla and Chlb were found to be closely associated with each other. In our experiment, the relationships between Chla and Chlb presented a high correlation with values of R
2 of 0.95 and 0.75 (
Figure 1 and
Figure 3) and the estimated ratio Chla/Chlb of 3.45 and 2.5 as an inverse of the slope of the regression line, under greenhouse and open field conditions, respectively. In this sense, Shah et al. [
14] reported a close relationship between both photosynthetic pigments, Chla being 2–4 times higher than Chlb. The differences in Chla/Chlb ratio depend on several factors like plant species, growth stage, and environmental conditions [
31]. It is well known that the Chla/Chlb ratio decreases under shading conditions [
32,
33]. In the case of the three species that were tested under both conditions, we noted two different trends:
H. rosa-sinensis presented similar ratios of Chla/Chlb under both climatic conditions whereas
M. deliciosa and
S. arboricola decreased their ratios under outdoor conditions. These results agreed with Casierra-Posada et al. [
34] who reported in
Calendula officinalis a higher Chla/Chlb ratio in leaves under shaded conditions since, under conditions of excess lighting, the photosystem II with a high presence of Chlb is more unstable than photosystem I rich with high presence of Chla. Moreover, PSII is more sensitive to heat stress than PSI; therefore, under heat stress, there is severe damage to the reaction center-binding protein D1 of PSII [
35].
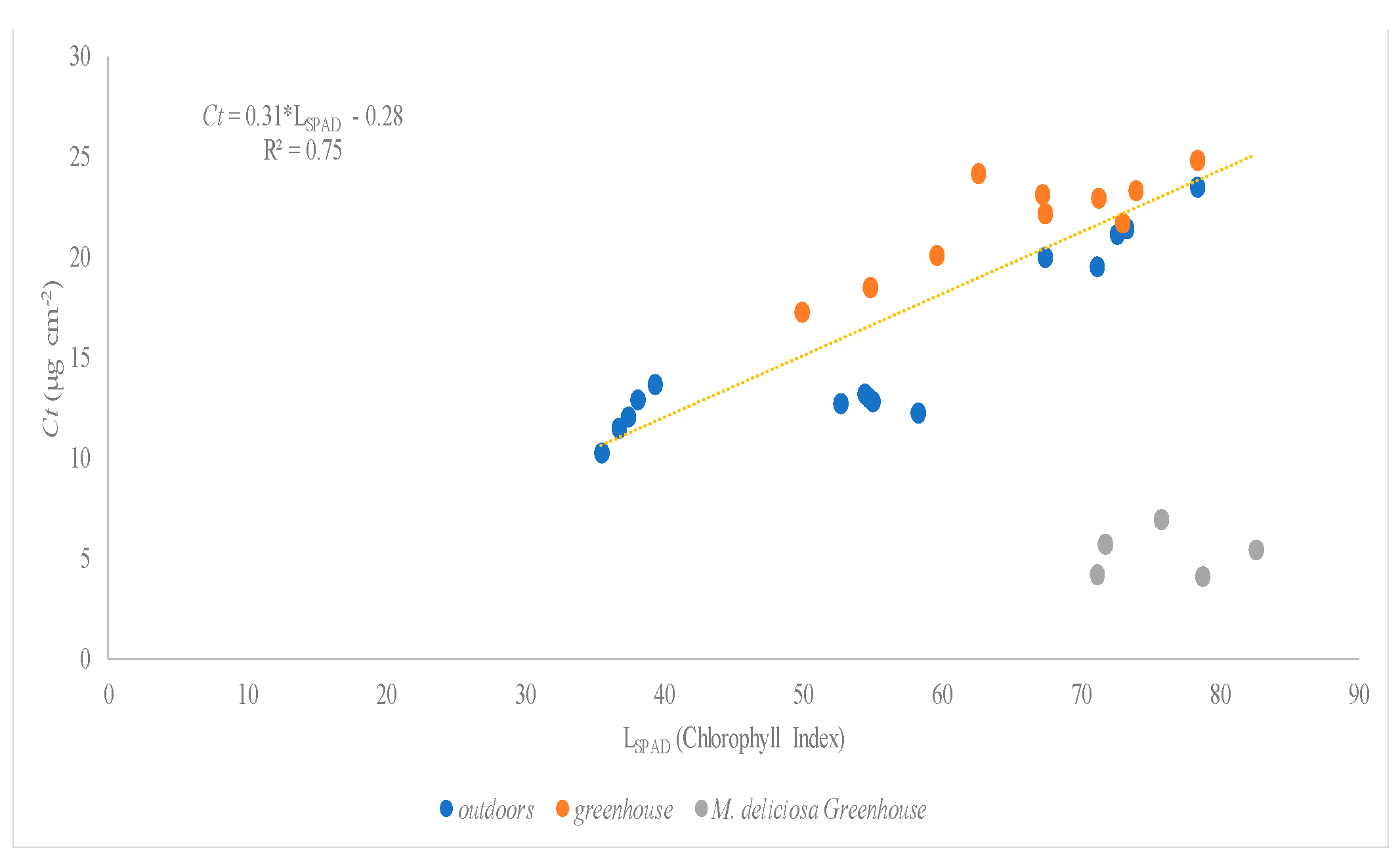
In our experiment, the relationships between Chlt and Ct showed values of R
2 of 0.98 and 0.91 under greenhouse and open field conditions, respectively. The slope was similar under the two assessed conditions (0.32 and 0.31) meaning that the total chlorophyll and carotenoid concentration ratio can be considered constant for all the species studied. This would allow us to make relationships between SPAD readings and carotenoid concentrations assessed. These pigments act as accessory pigments in photosystems and as photoprotective agents reducing the damaging effects of high light [
36].
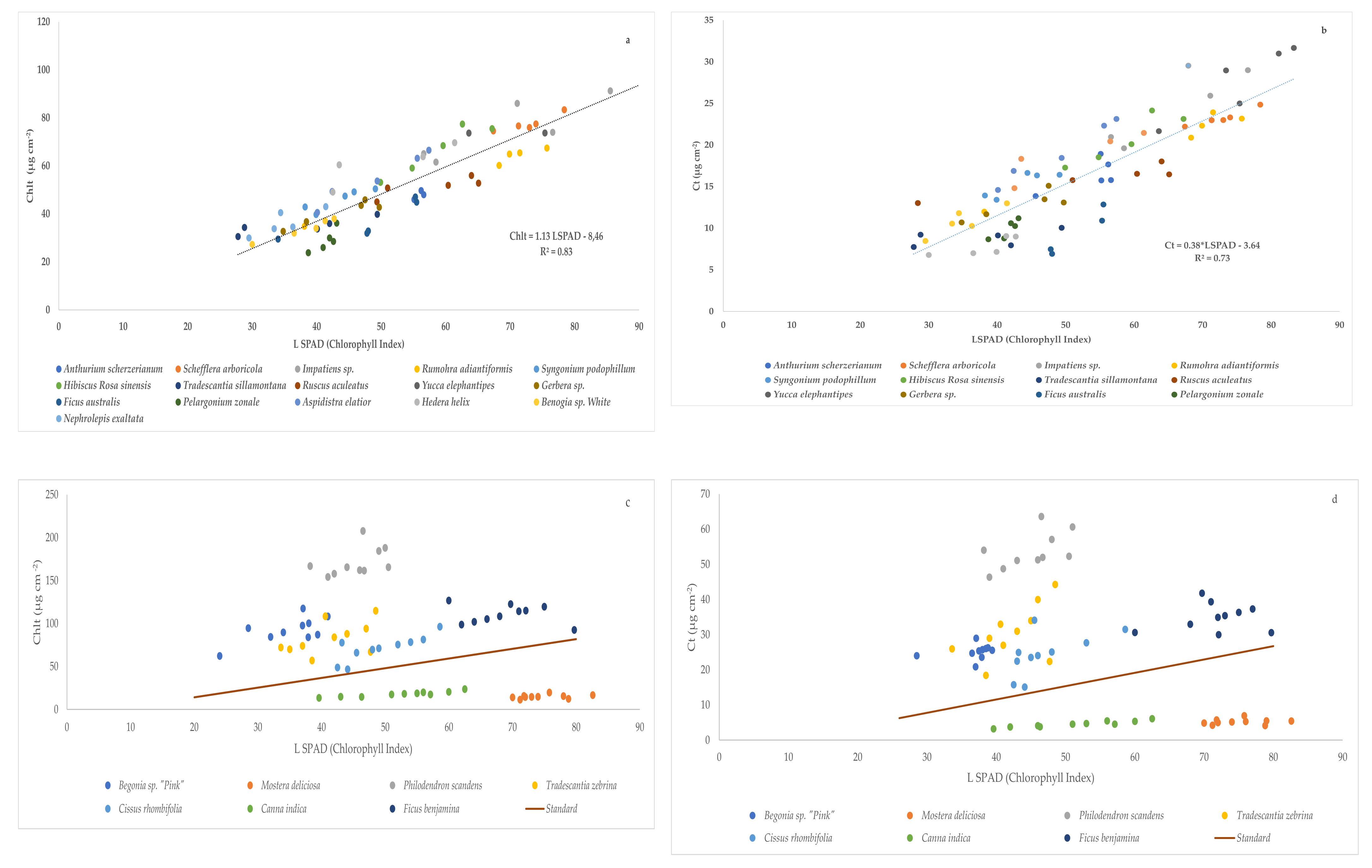
SPAD determinations are widely used to assess the absolute chlorophyll content per leaf area in research settings and agricultural systems [
13]. The relationship between Chlorophyll index or SPAD readings and Chlt content has been widely studied in different species such as
Euphorbia pulcherrima [
37],
Elaies guineensis [
38] and
Fraxinus chinensis,
Ginkgo biloba, and
Magnolia denudata [
39], between others, where there was a good correlation between both parameters when the readings of SPAD oscillated from 26 to 60 and also in the functioning of the chlorophyll extraction procedure assessed. Comparing the results obtained in our experiment with the previous literature, we can report that higher values of chlorophyll Index or SPAD readings were found in
F. benjamina under greenhouse conditions and
S. arboricola under open field conditions than those reported by Sardoei et al. [
40], but under greenhouse conditions, the value of CI in
S. arboricola reported by them was similar to our results. We also noted higher SPAD values (66.0) than those found by Massa et al. [
41] in
Impatiens sp. Nevertheless, the values reported by Parminder et al. [
29] in
N. exaltata agree with our results. The fitting of our data in both trials reported a linear relationship and values of R
2 close to 1 between chlorophyll index (LSPAD) readings and total chlorophyll (Chlt) and carotenoids (Ct) concentration. In this sense, Campbell et al. [
42] also found a linear model of SPAD–chlorophyll relationships between different experiments and environmental conditions. Nevertheless, Houborg et al. [
24] fitted an exponential model to the relationship between SPAD readings and dimethylsulfoxide (DMSO) extractable Chlt per leaf area.
Reviewing previous literature, there are scarce references about the relationship between SPAD-readings and carotenoid concentration. For instance, Shah et al. [
14] found a linear relationship in
Triticum aestivum with a slope of 0.12 and a coefficient of determination (R
2) on 0.85. In our experiment, we also noted a linear relationship between SPAD readings and Ct content per leaf area under greenhouse conditions with a slope of 0.38 and R
2 of 0.73 and at open field conditions with slope values ranging from 0.27 to 0.44 and values of R
2 ranging from 0.84 to 0.97. These differences in slope and values of R
2 can be ascribed to leaf anatomical modifications which may alter the spectral response and SPAD readings [
43]. However, in this specific study, the changes found in the regression coefficients of the fitted models did not vary significantly.
In our experiment, the three species studied in both growing conditions (greenhouse and open field conditions) have also been assessed to discern the relationships between chlorophyll index (LSPAD) readings and total chlorophyll (Chlt) and carotenoid (Ct) concentration. The results obtained reported that there are two species
H. rosa-sinensis and
S. arboricola, whose results obtained for the values of the content in Chlt and Ct conform to the same model of correlation with SPAD in both conditions of cultivation. On the contrary, the species,
M. deliciosa, shows very different correlations between these values in both conditions of cultivation, the correlation obtained in the greenhouse being significantly different from the general correlation. These results depicted that these relationships between pigments and SPAD readings can vary among the species due to several factors like leaf optical properties [
21].


 and
and 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}