
The Contribution of Microorganisms to Soil Organic Carbon Accumulation under Fertilization Varies among Aggregate Size Classes

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Soil Sampling
2.2. Aggregate Fractionation
2.3. Determination of Soil Physical and Chemical Properties
2.4. Analysis of Enzyme Activity
2.5. PLFA Extraction and Analysis
2.6. Data Calculation and Statistical Analysis
3. Results
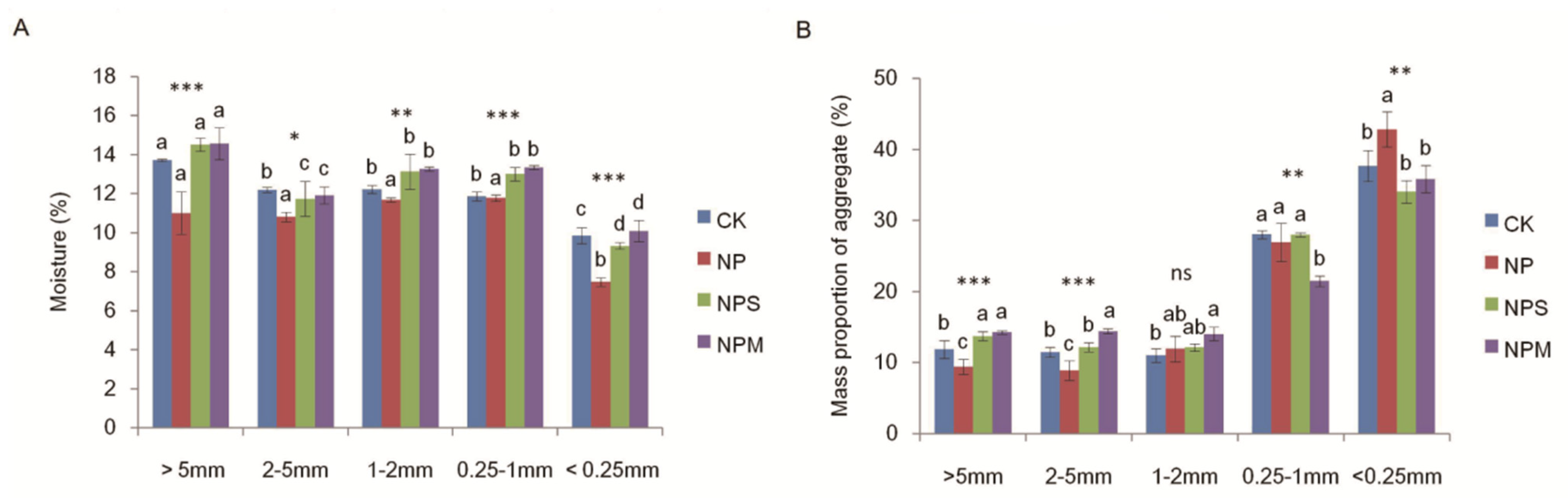
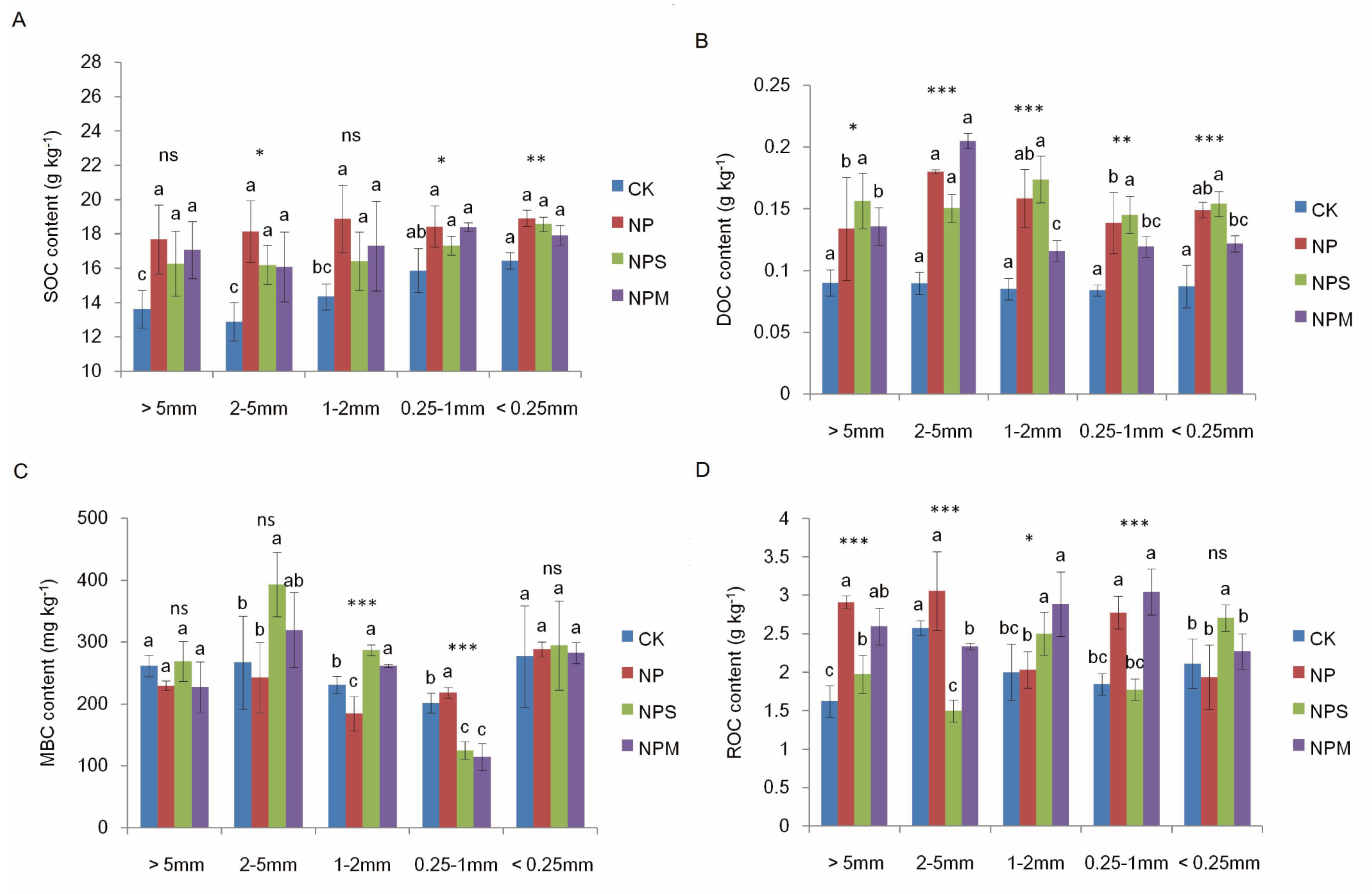
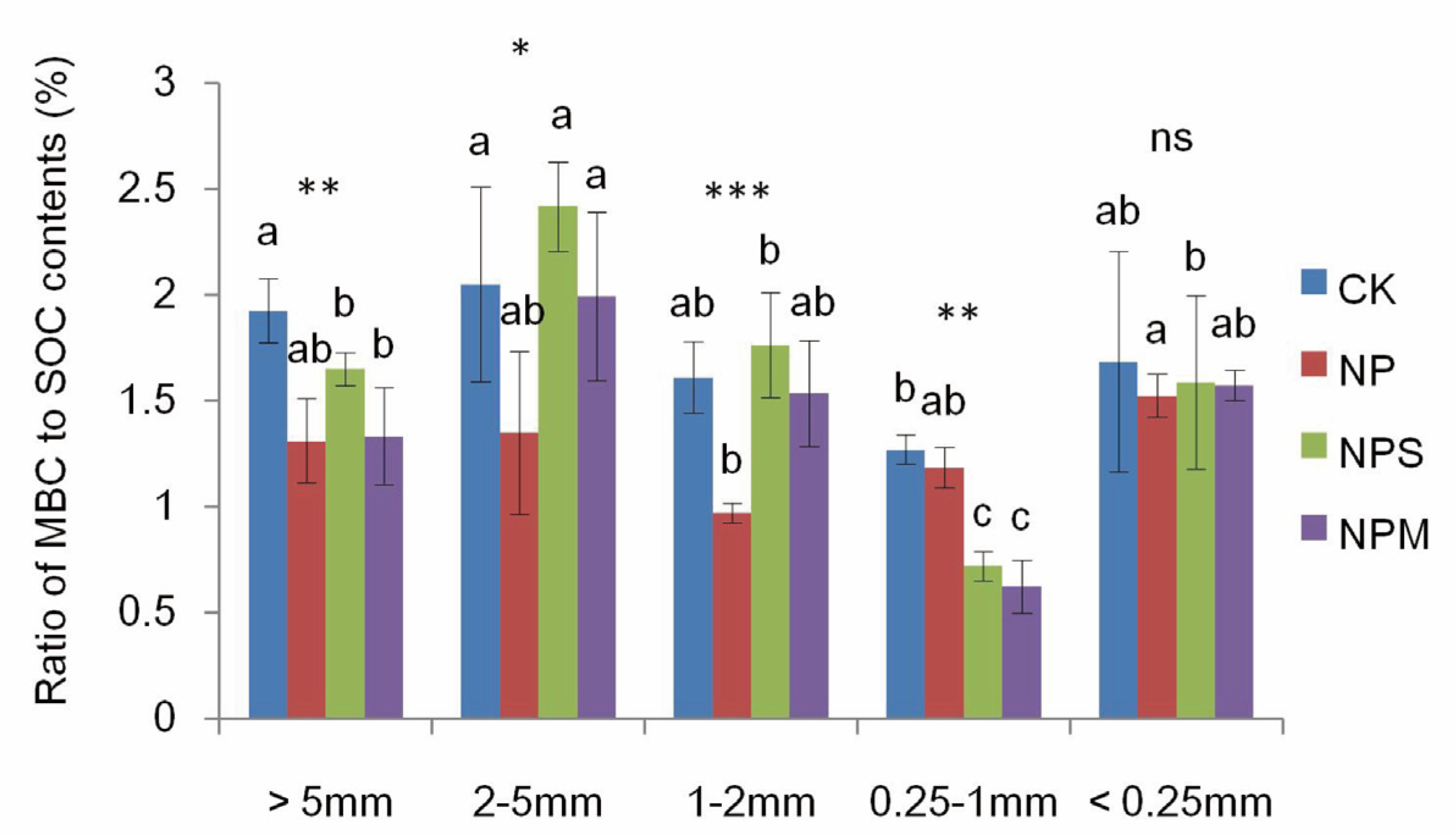
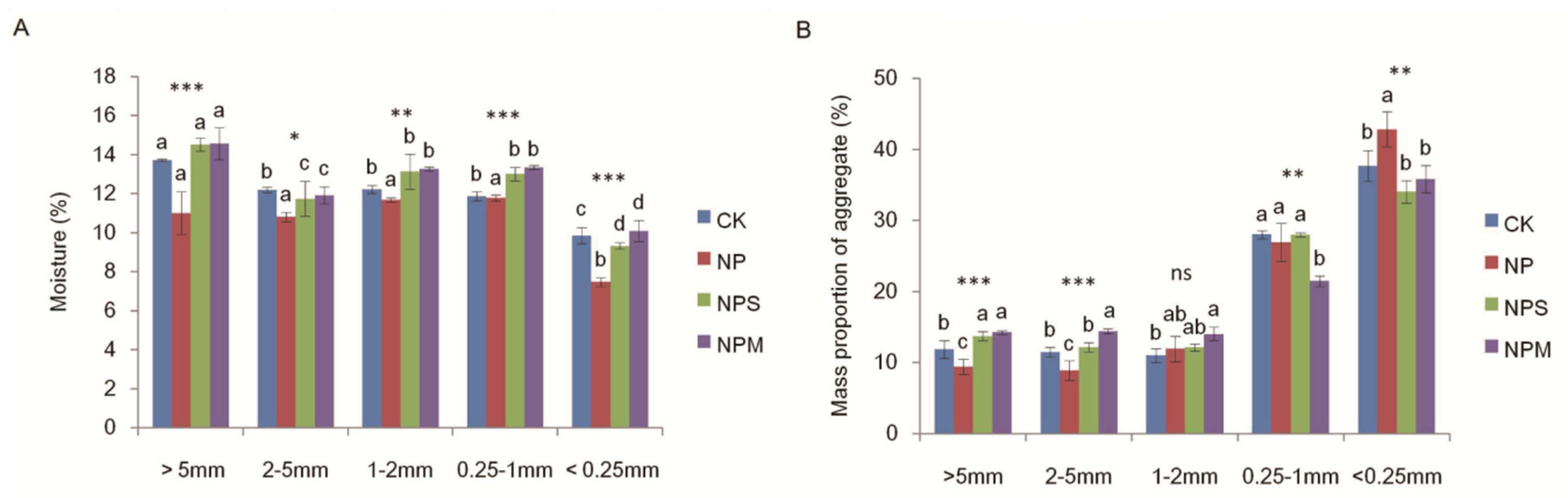
3.1. Soil Organic C and Moisturein Aggregate and Aggregate Proportions under Different Fertilizations
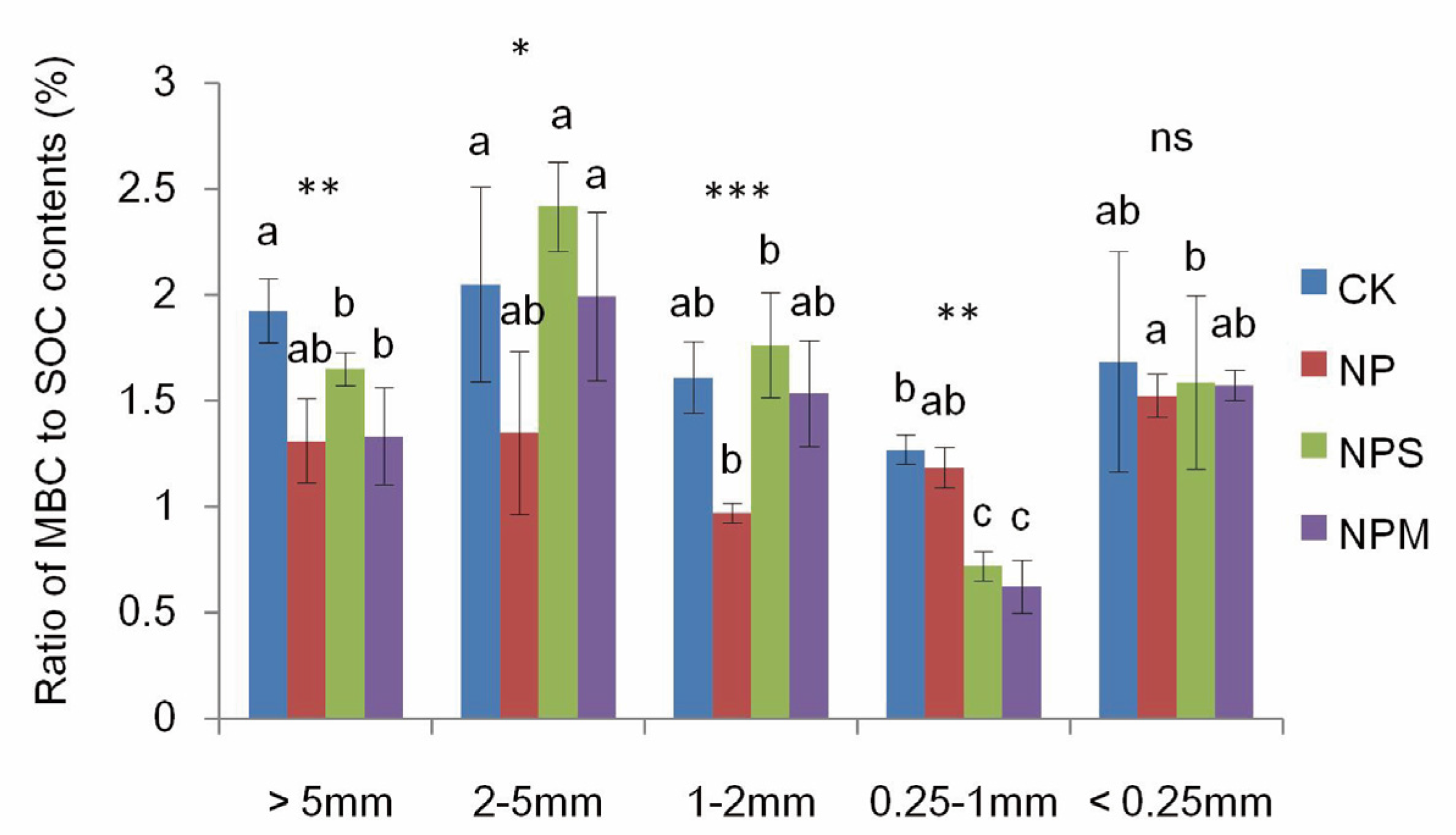
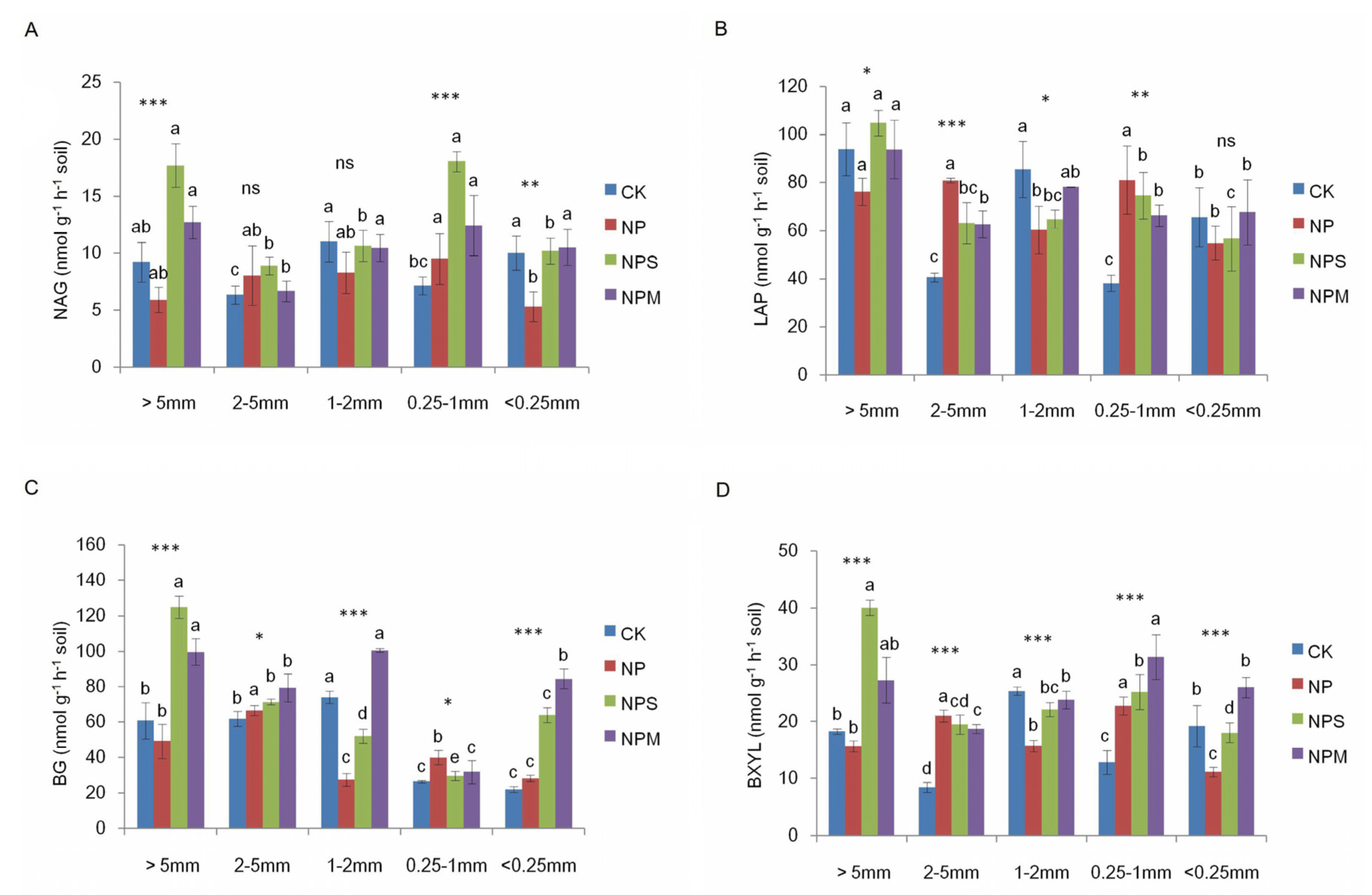
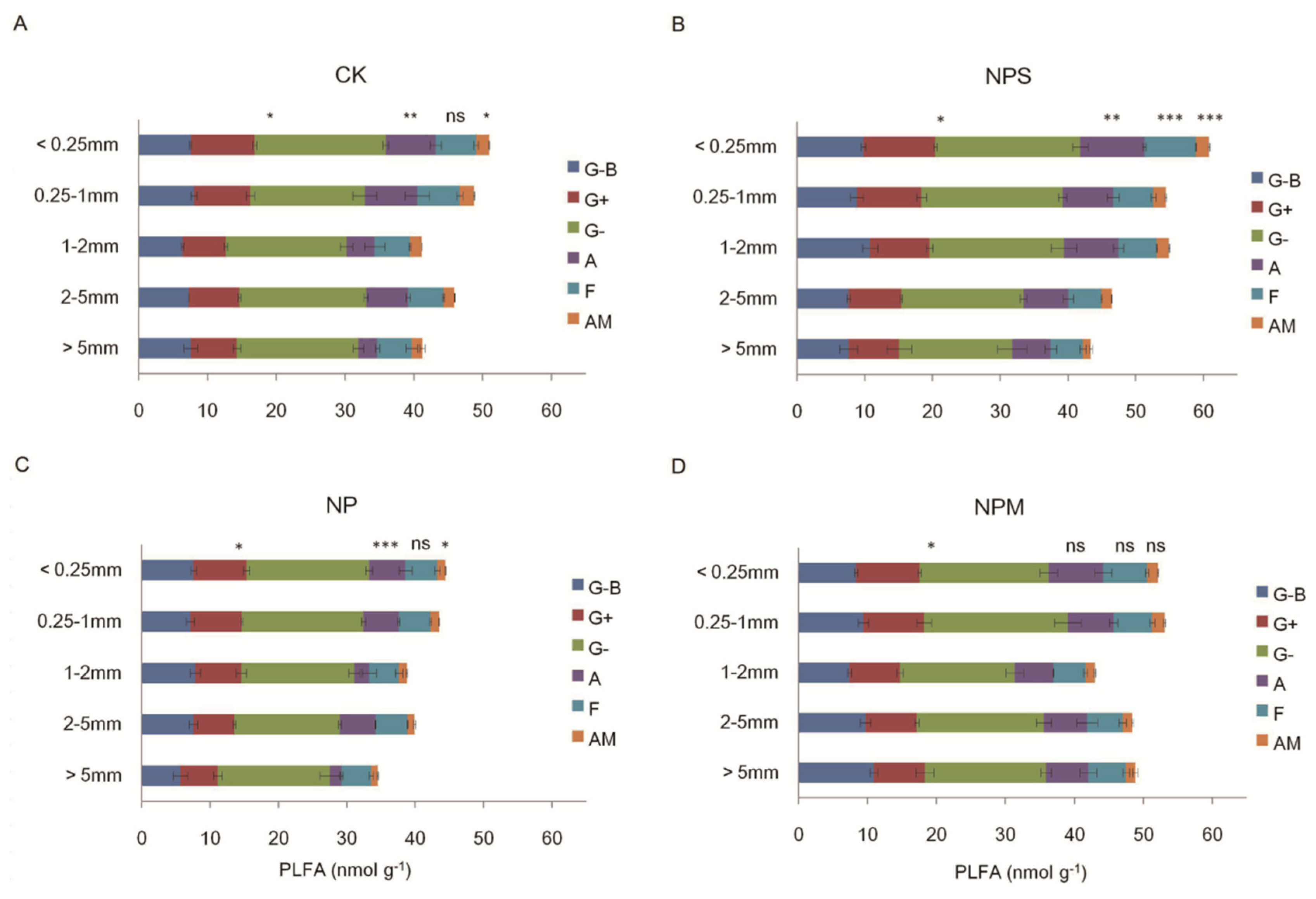
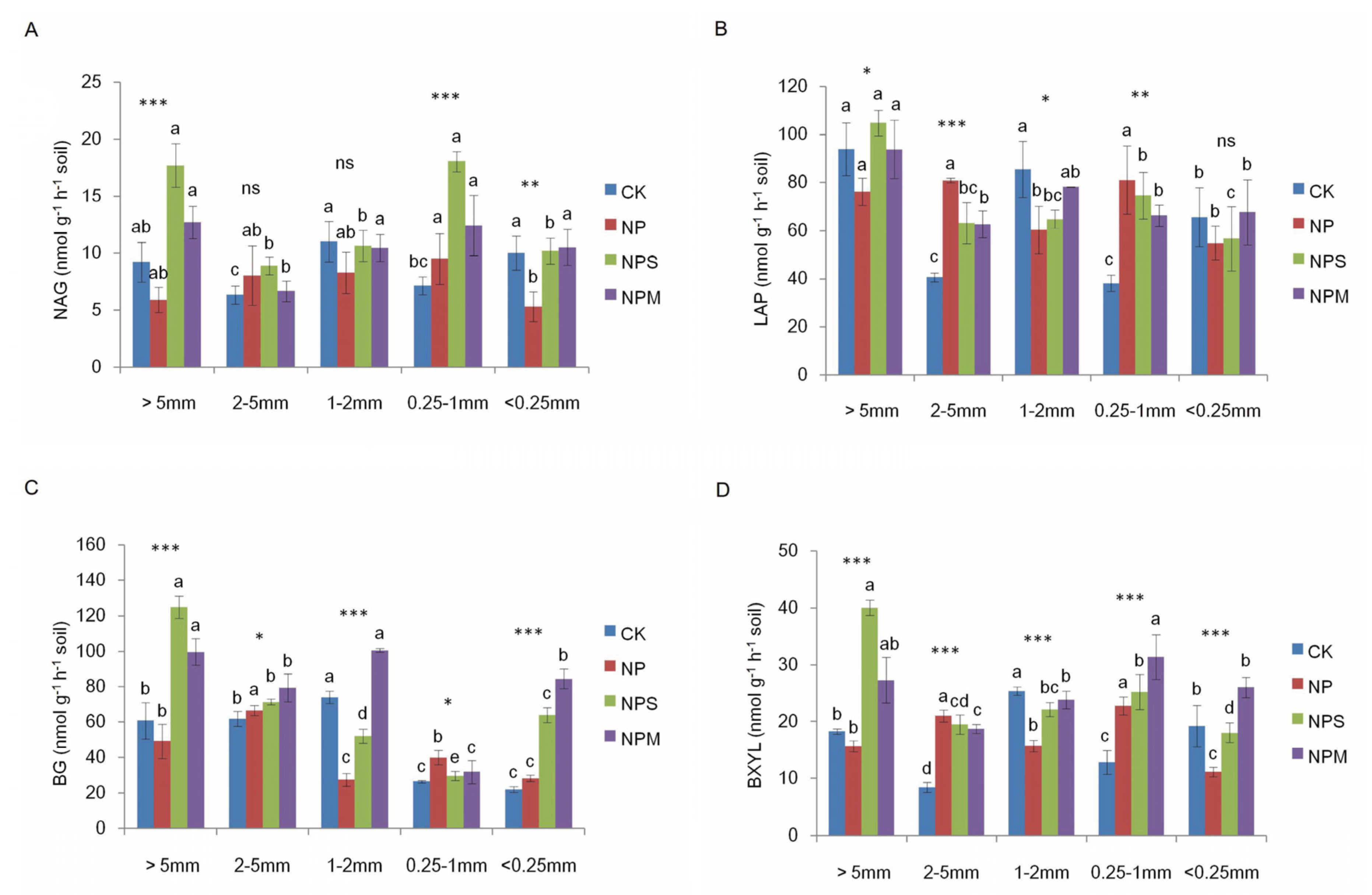
3.2. Soil Microbial Biomass and Diversity and Enzyme Activity
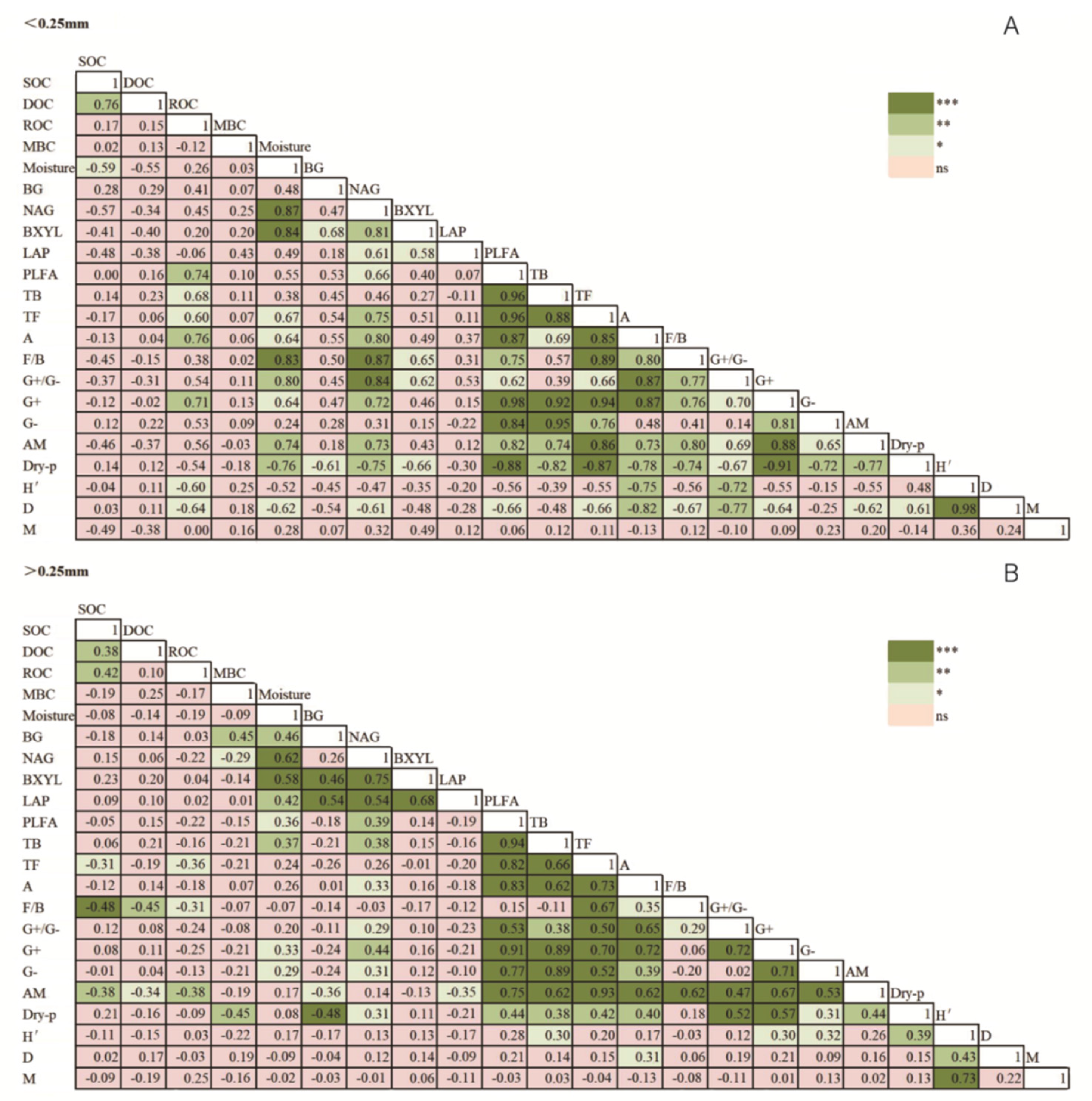
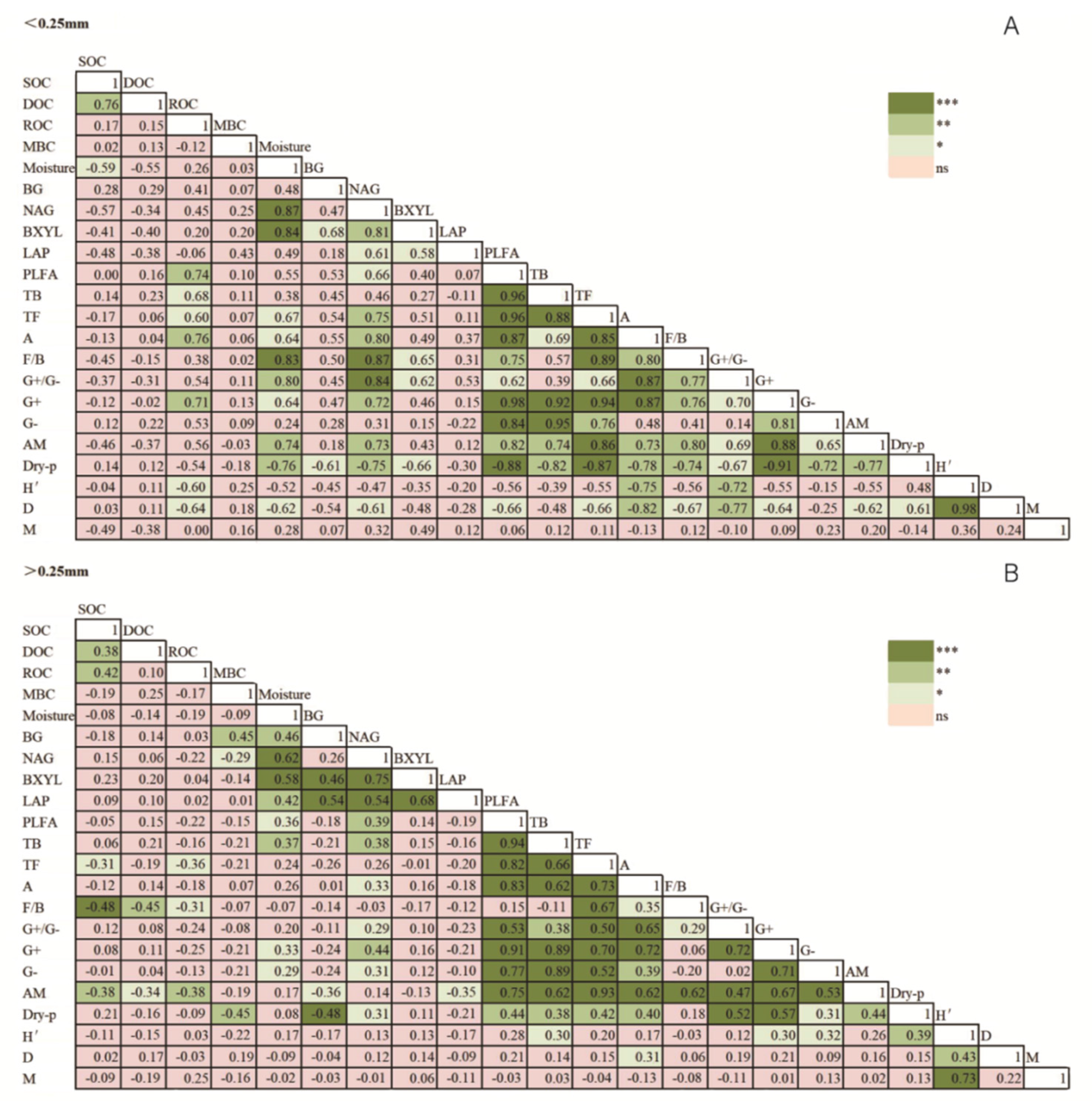
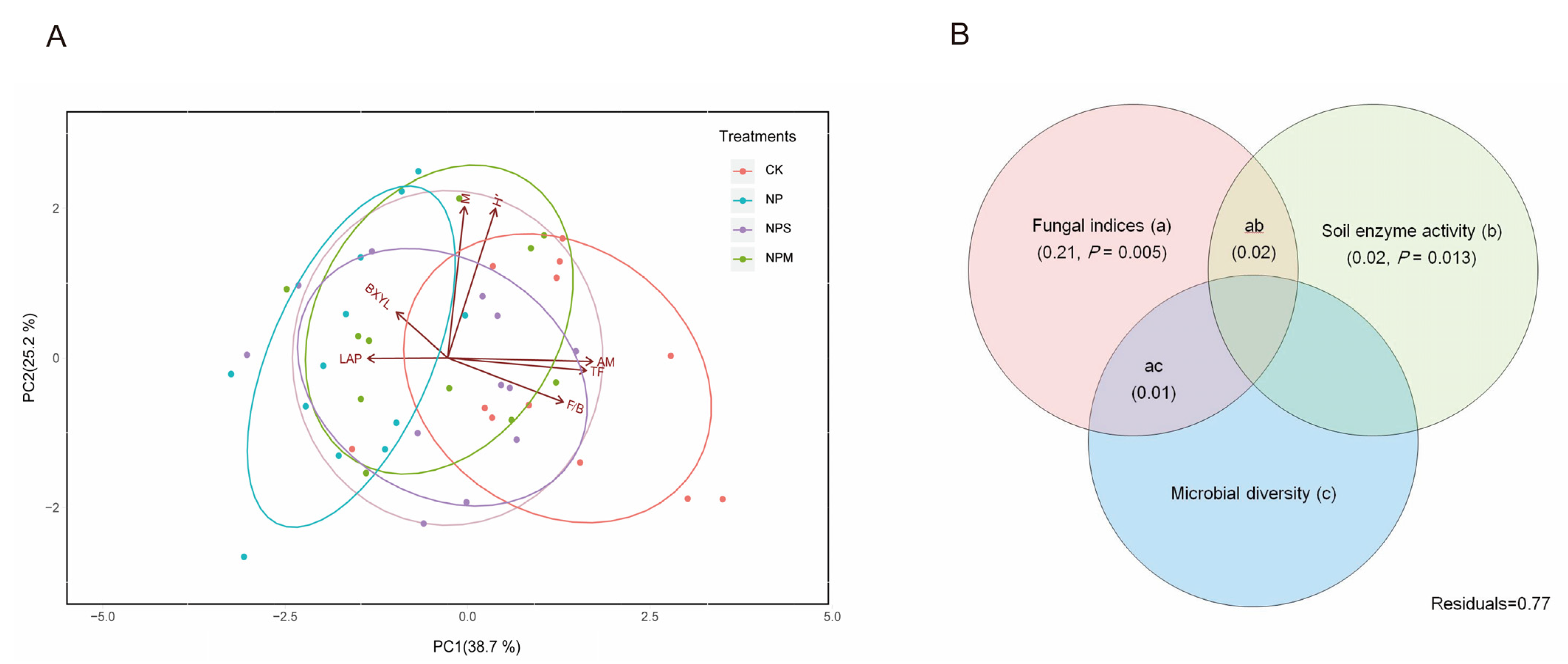
3.3. Contributions of Microbial Community and Soil Enzyme to SOC Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Yao, Z.; Zhan, Y.; Zheng, X.; Zhou, M.; Yan, G.; Wang, L.; Werner, C.; Butterbach-Bahl, K. Potential benefits of liming to acid soils on climate change mitigation and food security. Glob. Chang. Biol. 2021, 27, 2807–2821. [Google Scholar] [CrossRef] [PubMed]
- Kallenbach, C.M.; Frey, S.D.; Grandy, A.S. Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Crowther, T.W.; Van den Hoogen, J.; Wan, J.; Mayes, M.A.; Keiser, A.D.; Mo, L.; Averill, C.; Maynard, D.S. The global soil community and its influence on biogeochemistry. Science 2019, 365, eaav0550. [Google Scholar] [CrossRef] [PubMed]
- Melillo, J.M.; Frey, S.D.; DeAngelis, K.M.; Werner, W.J.; Bernard, M.J.; Bowles, F.P.; Pold, G.; Knorr, M.A.; Grandy, A.S. Long-term pattern and magnitude of soil carbon feedback to the climate system in a warming world. Science 2017, 358, 101–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, S.; Sun, H.; Yang, J.; Qin, J.; Shen, D.; Chen, Y. Variations in soil enzyme activities and microbial communities along an altitudinal gradient on the eastern Qinghai–Tibetan plateau. Forests 2021, 12, 681. [Google Scholar] [CrossRef]
- Merino, C.; Matus, F.; Kuzyakov, Y.; Dyckmans, J.; Stock, S.; Dippold, M.A. Contribution of the Fenton reaction and ligninolytic enzymes to soil organic matter mineralisation under anoxic conditions. Sci. Total Environ. 2021, 760, 143397. [Google Scholar] [CrossRef] [PubMed]
- Bond-Lamberty, B.; Bailey, V.L.; Chen, M.; Gough, C.M.; Vargas, R. Globally rising soil heterotrophic respiration over recent decades. Nature 2018, 560, 80–83. [Google Scholar] [CrossRef]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Juan-Ovejero, R.; Briones, M.J.I.; Öpik, M. Fungal diversity in peatlands and its contribution to carbon cycling. Appl. Soil Ecol. 2020, 146, 103393. [Google Scholar] [CrossRef]
- Baumann, K.; Dignac, M.F.; Rumpel, C.; Bardoux, G.; Sarr, A.; Steffens, M.; Maron, P.A. Soil microbial diversity affects soil organic matter decomposition in a silty grassland soil. Biogeochemistry 2013, 114, 201–212. [Google Scholar] [CrossRef]
- Zhang, S.; Li, Q.; Lü, Y.; Zhang, X.; Liang, W. Contributions of soil biota to C sequestration varied with aggregate fractions under different tillage systems. Soil Biol. Biochem. 2013, 62, 147–156. [Google Scholar] [CrossRef]
- Ma, Q.; Wen, Y.; Wang, D.; Sun, X.; Hill, P.W.; Macdonald, A.; Chadwick, D.R.; Wu, L.; Jones, D.L. Farmyard manure applications stimulate soil carbon and nitrogen cycling by boosting microbial biomass rather than changing its community composition. Soil Biol. Biochem. 2020, 144, 107760. [Google Scholar]
- Romaní, A.M.; Fischer, H.; Mille-Lindblom, C.; Tranvik, L.J. Interactions of bacteria and fungi on decomposing litter: Differential extracellular enzyme activities. Ecology 2006, 87, 2559–2569. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Yuan, M.; Jian, S.; Gamage, L.; Parajuli, M.; Dzantor, K.E.; Hui, D.; Fay, P.A.; Li, J. Soil extracellular oxidases mediated nitrogen fertilization effects on soil organic carbon sequestration in bioenergy croplands. Glob. Chang. Biol. 2021, 13, 1303–1318. [Google Scholar] [CrossRef]
- Xu, C.Y.; Du, C.; Jian, J.S.; Hou, L.; Wang, Z.K.; Wang, Q.; Geng, Z.C. The interplay of labile organic carbon, enzyme activities and microbial communities of two forest soils across seasons. Sci. Rep. 2021, 11, 1–12. [Google Scholar]
- He, L.; Lu, S.; Wang, C.; Mu, J.; Zhang, Y.; Wang, X. Changes in soil organic carbon fractions and enzyme activities in response to tillage practices in the Loess Plateau of China. Soil Tillage Res. 2021, 209, 104940. [Google Scholar] [CrossRef]
- Somasundaram, J.; Reeves, S.; Wang, W.; Heenan, M.; Dalal, R. Impact of 47 years of no tillage and stubble retention on soil aggregation and carbon distribution in a vertisol. L. Degrad. Dev. 2017, 28, 1589–1602. [Google Scholar] [CrossRef]
- Elliott, E.T. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Sci. Soc. Am. J. 1986, 50, 627–633. [Google Scholar] [CrossRef]
- Young, I.M.; Ritz, K. Tillage, habitat space and function of soil microbes. Soil Tillage Res. 2000, 53, 201–213. [Google Scholar] [CrossRef]
- Briar, S.S.; Fonte, S.J.; Park, I.; Six, J.; Scow, K.; Ferris, H. The distribution of nematodes and soil microbial communities across soil aggregate fractions and farm management systems. Soil Biol. Biochem. 2011, 43, 905–914. [Google Scholar] [CrossRef]
- Larkin, A.A.; Martiny, A.C. Microdiversity shapes the traits, niche space, and biogeography of microbial taxa. Environ. Microbiol. Rep. 2017, 9, 55–70. [Google Scholar] [CrossRef]
- Wisz, M.S.; Pottier, J.; Kissling, W.D.; Pellissier, L.; Lenoir, J.; Damgaard, C.F.; Dormann, C.F.; Forchhammer, M.C.; Grytnes, J.; Guisan, A. The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorodnikov, M.; Blagodatskaya, E.; Blagodatsky, S.; Marhan, S.; Fangmeier, A.; Kuzyakov, Y. Stimulation of microbial extracellular enzyme activities by elevated CO2 depends on soil aggregate size. Glob. Chang. Biol. 2009, 15, 1603–1614. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Philippot, L. Insights into the resistance and resilience of the soil microbial community. FEMS Microbiol. Rev. 2013, 37, 112–129. [Google Scholar] [PubMed] [Green Version]
- Liang, Q.; Chen, H.; Gong, Y.; Yang, H.; Fan, M.; Kuzyakov, Y. Effects of 15 years of manure and mineral fertilizers on enzyme activities in particle-size fractions in a North China Plain soil. Eur. J. Soil Biol. 2014, 60, 112–119. [Google Scholar] [CrossRef]
- Liu, M.; Li, P.; Liu, M.; Wang, J.; Chang, Q. The trend of soil organic carbon fractions related to the successions of different vegetation types on the tableland of the Loess Plateau of China. J. Soils Sediments. 2021, 21, 203–214. [Google Scholar]
- Inselsbacher, E.; Umana, N.H.-N.; Stange, F.C.; Gorfer, M.; Schüller, E.; Ripka, K.; Zechmeister-Boltenstern, S.; Hood-Novotny, R.; Strauss, J.; Wanek, W. Short-term competition between crop plants and soil microbes for inorganic N fertilizer. Soil Biol. Biochem. 2010, 42, 360–372. [Google Scholar]
- Li, S.X.; Xiao, L. Distribution and management of drylands in the People’s Republic of China. Adv. Soil Sci. 1992, 147–302. [Google Scholar]
- Yu, Q.; Hu, X.; Ma, J.; Ye, J.; Sun, W.; Wang, Q.; Lin, H. Effects of long-term organic material applications on soil carbon and nitrogen fractions in paddy fields. Soil Tillage Res. 2020, 196, 104483. [Google Scholar] [CrossRef]
- Qiu, G.; Zhu, M.; Contin, M.; De Nobili, M.; Luo, Y.; Xu, J.; Brookes, P.C. Evaluating the triggering responsein soils, using 13C-glucose, and effects on dynamics of microbial biomass. Soil Biol. Biochem. 2020, 33, 107843. [Google Scholar] [CrossRef]
- Wang, Y.; Li, C.; Tu, C.; Hoyt, G.D.; DeForest, J.L.; Hu, S. Long-term no-tillage and organic input management enhanced the diversity and stability of soil microbial community. Sci. Total Environ. 2017, 609, 341–347. [Google Scholar] [CrossRef]
- Tripathi, R.; Nayak, A.K.; Bhattacharyya, P.; Shukla, A.K.; Shahid, M.; Raja, R.; Panda, B.B.; Mohanty, S.; Kumar, A.; Thilagam, V.K. Soil aggregation and distribution of carbon and nitrogen in different fractions after 41 years long-term fertilizer experiment in tropical rice–rice system. Geoderma 2014, 213, 280–286. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- DeForest, J.L. The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and l-DOPA. Soil Biol. Biochem. 2009, 41, 1180–1186. [Google Scholar] [CrossRef]
- German, D.P.; Weintraub, M.N.; Grandy, A.S.; Lauber, C.L.; Rinkes, Z.L.; Allison, S.D. Optimization of hydrolytic and oxidative enzyme methods for ecosystem studies. Soil Biol. Biochem. 2011, 43, 1387–1397. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M.; Gunapala, N.; Department, K.J.G. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef]
- Mo, F.; Zhang, Y.Y.; Liu, Y.; Liao, Y.C. Microbial carbon-use efficiency and straw-induced priming effect within soil aggregates are regulated by tillage history and balanced nutrient supply. Biol. Fertil. Soils. 2021, 57, 409–420. [Google Scholar] [CrossRef]
- Chakraborty, A.; Chakrabarti, K.; Chakraborty, A.; Ghosh, S. Effect of long-term fertilizers and manure application on microbial biomass and microbial activity of a tropical agricultural soil. Biol. Fertil. Soils. 2011, 47, 227–233. [Google Scholar] [CrossRef]
- Yang, Q.; Zheng, F.; Jia, X.; Liu, P.; Dong, S.; Zhang, J.; Zhao, B. The combined application of organic and inorganic fertilizers increases soil organic matter and improves soil microenvironment in wheat-maize field. J. Soils Sediments. 2020, 20, 2395–2404. [Google Scholar] [CrossRef]
- Manzoni, S.; Schimel, J.P.; Porporato, A. Responses of soil microbial communities to water stress: Results from a meta-analysis. Ecology 2012, 93, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Ye, G.; Kuzyakov, Y.; Liu, D.; Fan, J.; Ding, W. Long-term manure application increases soil organic matter and aggregation, and alters microbial community structure and keystone taxa. Soil Biol. Biochem. 2019, 134, 187–196. [Google Scholar] [CrossRef]
- Trivedi, P.; Rochester, I.J.; Trivedi, C.; Van Nostrand, J.D.; Zhou, J.; Karunaratne, S.; Anderson, I.C.; Singh, B.K. Soil aggregate size mediates the impacts of cropping regimes on soil carbon and microbial communities. Soil Biol. Biochem. 2015, 91, 169–181. [Google Scholar] [CrossRef]
- Wei, L.; Vosátka, M.; Cai, B.; Ding, J.; Lu, C.; Xu, J.; Yan, W.; Li, Y.; Liu, C. The role of arbuscular mycorrhiza fungi in the decomposition of fresh residue and soil organic carbon: A Mini-Review. Soil Sci. Soc. Am. J. 2019, 83, 511–517. [Google Scholar] [CrossRef]
- Legrand, F.; Picot, A.; Cobo-Díaz, J.F.; Carof, M.; Chen, W.; Le Floch, G. Effect of tillage and static abiotic soil properties on microbial diversity. Appl. Soil Ecol. 2018, 132, 135–145. [Google Scholar] [CrossRef]
- Zhang, J.; Sayer, E.J.; Zhou, J.; Li, Y.; Li, Y.; Li, Z.; Wang, F. Long-term fertilization modifies the mineralization of soil organic matter in response to added substrate. Sci. Total Environ. 2021, 798, 149341. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, K.; Wang, W.; Ren, G.; Khan, A.; Feng, Y.; Yang, G. Changes in soil enzymes, soil properties, and maize crop productivity under wheat straw mulching in Guanzhong, China. Soil Tillage Res. 2018, 182, 94–102. [Google Scholar] [CrossRef]
- Liu, E.; Yan, C.; Mei, X.; He, W.; Bing, S.H.; Ding, L.; Liu, Q.; Liu, S.; Fan, T. Long-term effect of chemical fertilizer, straw, and manure on soil chemical and biological properties in northwest China. Geoderma 2010, 158, 173–180. [Google Scholar] [CrossRef]
- Caldwell, B.A. Enzyme activities as a component of soil biodiversity: A review. Pedobiologia 2005, 49, 637–644. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Drought decreases soil enzyme activity in a Mediterranean Quercus ilex L. forest. Soil Biol. Biochem. 2005, 37, 455–461. [Google Scholar] [CrossRef]
- Six, J.; Elliott, E.T.; Paustian, K. Soil macroaggregate turnover and microaggregate formation: A mechanism for C sequestration under no-tillage agriculture. Soil Biol. Biochem. 2000, 32, 2099–2103. [Google Scholar] [CrossRef]
- Hurisso, T.T.; Davis, J.G.; Brummer, J.E.; Stromberger, M.E.; Mikha, M.M.; Haddix, M.L.; Booher, M.R.; Paul, E.A. Rapid changes in microbial biomass and aggregate size distribution in response to changes in organic matter management in grass pasture. Geoderma 2013, 193–194, 68–75. [Google Scholar] [CrossRef]
- Demoling, F.; Figueroa, D.; Bååth, E. Comparison of factors limiting bacterial growth in different soils. Soil Biol. Biochem. 2007, 39, 2485–2495. [Google Scholar] [CrossRef]
- Gupta, V.V.S.R.; Germida, J.J. Soil aggregation: Influence on microbial biomass and implications for biological processes. Soil Biol. Biochem. 2015, 80, A3–A9. [Google Scholar] [CrossRef]
- Tisdall, J.M.; Oades, J. Organic matter and water-stable aggregates in soils. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ranalli, M.G.; Haddix, M.L.; Six, J.; Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 2019, 12, 989–994. [Google Scholar] [CrossRef]
- Kögel-Knabner, I.; Guggenberger, G.; Kleber, M.; Kandeler, E.; Kalbitz, K.; Scheu, S.; Eusterhues, K.; Leinweber, P. Organo-mineral associations in temperate soils: Integrating biology, mineralogy, and organic matter chemistry. J. Plant Nutr. Soil Sci. 2008, 171, 61–82. [Google Scholar] [CrossRef]
- Jasinska, E.; Wetzel, H.; Baumgartl, T.; Horn, R. Heterogeneity of physico-chemical properties in structured soils and its consequences. Pedosphere 2006, 16, 284–296. [Google Scholar] [CrossRef]
- Guggenberger, G.; Frey, S.D.; Six, J.; Paustian, K.; Elliott, E.T. Bacterial and fungal cell-wall residues in conventional and no-tillage agroecosystems. Soil Sci. Soc. Am. J. 1999, 63, 1188–1198. [Google Scholar] [CrossRef]
- Cheng, L.; Booker, F.L.; Tu, C.; Burkey, K.O.; Zhou, L.; Shew, H.D.; Rufty, T.W.; Hu, S. Arbuscular mycorrhizal fungi increase organic carbon decomposition under elevated CO2. Science 2012, 337, 1084–1087. [Google Scholar] [CrossRef]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Zhou, G.; Chang, D.; Gao, S.; Liang, T.; Liu, R.; Cao, W. Co-incorporating leguminous green manure and rice straw drives the synergistic release of carbon and nitrogen, increases hydrolase activities, and changes the composition of main microbial groups. Biol. Fertil. Soils. 2021, 57, 547–561. [Google Scholar] [CrossRef]
- Ren, Y.; Zhang, L.; Yang, K.; Li, Z.; Yin, R.; Tan, B.; Wang, L.; Liu, Y.; Li, H.; You, C.; et al. Short-term effects of snow cover manipulation on soil bacterial diversity and community composition. Sci. Total Environ. 2020, 741, 140454. [Google Scholar] [CrossRef] [PubMed]
- Baumert, V.L.; Vasilyeva, N.A.; Vladimirov, A.A.; Meier, I.C.; Kögel-Knabner, I.; Mueller, C.W. Root exudates induce soil macroaggregation facilitated by fungi in subsoil. Front. Environ. Sci. 2018, 6, 1–17. [Google Scholar]
- Shi, Y.; Li, Y.; Xiang, X.; Sun, R.; Yang, T.; He, D.; Zhang, K.; Ni, Y.; Zhu, Y.G.; Adams, J.M.; et al. Spatial scale affects the relative role of stochasticity versus determinism in soil bacterial communities in wheat fields across the North China Plain. Microbiome 2018, 6, 1–12. [Google Scholar]
- Shi, W.; Zhao, H.Y.; Chen, Y.; Wang, J.S.; Han, B.; Li, C.P.; Lu, J.Y.; Zhang, L.M. Organic manure rather than phosphorus fertilization primarily determined asymbiotic nitrogen fixation rate and the stability of diazotrophic community in an upland red soil. Agric. Ecosyst. Environ. 2021, 319, 107535. [Google Scholar]
- Cai, A.; Xu, M.; Wang, B.; Zhang, W.; Liang, G.; Hou, E.; Luo, Y. Manure acts as a better fertilizer for increasing crop yields than synthetic fertilizer does by improving soil fertility. Soil Tillage Res. 2019, 189, 168–175. [Google Scholar]
- Li, Y.; Chen, H.; Feng, H.; Dong, Q.; Wu, W.; Zou, Y.; Chau, H.W.; Siddique, K.H.M. Influence of straw incorporation on soil water utilization and summer maize productivity: A five-year field study on the Loess Plateau of China. Agric. Water Manag. 2020, 233, 106106. [Google Scholar]
- Mondini, C.; Contin, M.; Leita, L.; De Nobili, M. Response of microbial biomass to air-drying and rewetting in soils and compost. Geoderma 2002, 105, 111–124. [Google Scholar] [CrossRef]
- Thormann, M.N. Diversity and function of fungi in peatlands: A carbon cycling perspective. Can. J. Soil Sci. 2006, 86, 281–293. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indices | CK | NP | NPS | NPM |
|---|---|---|---|---|
| pH | 7.80 | 7.83 | 7.79 | 7.77 |
| Bulk density (g cm−3) | 1.28 | 1.30 | 1.23 | 1.20 |
| SOC (g kg−1) | 12.35 | 18.26 | 17.67 | 17.15 |
| Indices | Treatment (T) F P | Aggregate Size (A) F P | T × A F P |
|---|---|---|---|
| SOC | 19.1 *** | 4.81 ** | 0.55 |
| DOC | 57.15 *** | 8.01 *** | 4.33 *** |
| ROC | 14.82 *** | 0.35 | 7.6 *** |
| MBC | 2.08 | 17.73 *** | 4.51 *** |
| BG | 35.32 *** | 5.54 ** | 2.66 * |
| BXYL | 39.34 *** | 14.74 *** | 2.11 * |
| NAG | 12.23 *** | 17.37 *** | 1.61 |
| LAP | 4.08 * | 54.97 *** | 3 ** |
| Total PLFA | 35.05 *** | 17.59 *** | 3.00 ** |
| Bacteria | 22.39 *** | 9.91 *** | 3.22 ** |
| Fungi | 23.08 *** | 12.23 *** | 2.44 * |
| AM | 46.27 *** | 15.89 *** | 2.18 * |
| Actinomycetes | 21.84 *** | 12.38 *** | 1.72 |
| G+ | 20.73 *** | 22.59 *** | 0.99 |
| G− | 8.16 *** | 5.17 ** | 2.26 * |
| G+/G− | 5.87 ** | 10.33 *** | 1.91 |
| F/B | 10.46 *** | 1.69 | 1.56 |
| H′ | 0.95 | 6 *** | 3.48 ** |
| D | 0.41 | 5.2 ** | 2.11 * |
| M | 1.92 | 2.13 | 2.62 * |
| Moisture | 54.91 *** | 137.16 *** | 4.85 *** |
| Dry-p | 0 | 879.68 *** | 14.77 *** |
| Microbial Indices | Soil Fraction (mm) | Fertilizer Management | p Value | |||
|---|---|---|---|---|---|---|
| CK | NP | NPS | NPM | |||
| F/B | >5 mm | 21.71 ± 2.19 Ab | 18.00 ± 5.07 Aa | 18.75 ± 0.52 Ab | 19.67 ± 2.80 Aa | ns |
| 2–5 mm | 20.42 ± 0.18 Ab | 19.72 ± 1.22 Aa | 19.22 ± 0.20 Ab | 18.59 ± 2.12 Aa | ns | |
| 1–2 mm | 22.53 ± 0.75 Aab | 17.74 ± 2.29 Ba | 19.09 ± 1.48 Bb | 19.21 ± 1.02 Ba | * | |
| 0.25–1 mm | 25.17 ± 1.22 Aa | 18.19 ± 0.46 Ba | 19.78 ± 0.61 Bb | 19.05 ± 0.99 Ba | *** | |
| <0.25 mm | 21.68 ± 1.00 Ab | 17.69 ± 1.11 Ba | 22.86 ± 0.86 Aa | 22.03 ± 1.36 Aa | ** | |
| p value | * | ns | *** | ns | ||
| G+/G− | >5 mm | 0.37 ± 0.22 ABb | 0.34 ± 0.04 Bb | 0.45 ± 0.03 Ab | 0.42 ± 0.06 ABa | ns |
| 2–5 mm | 0.40 ± 0.02 ABb | 0.39 ± 0.02 Bab | 0.43 ± 0.01 Ab | 0.40 ± 0.02 ABa | ns | |
| 1–2 mm | 0.36 ± 0.03 Ab | 0.41 ± 0.05 Aa | 0.44 ± 0.02 Ab | 0.44 ± 0.05 Aa | ns | |
| 0.25–1 mm | 0.50 ± 0.06 Aa | 0.42 ± 0.01 Aa | 0.46 ± 0.02 Ab | 0.42 ± 0.02 Aa | ns | |
| <0.25 mm | 0.49 ± 0.03 Aa | 0.44 ± 0.02 Ba | 0.50 ± 0.01 Aa | 0.50 ± 0.03 Aa | ns | |
| p value | ** | * | * | ns | ||
| Microbial diversity | ||||||
| Shannon–Wiener (H′) | >5 mm | 3.21 ± 0.09 ABb | 3.10 ± 0.07 Bc | 3.26 ± 0.05 Aa | 3.23 ± 0.08 ABb | ns |
| 2–5 mm | 3.38 ± 0.03 Aa | 3.16 ± 0.04 Bc | 3.19 ± 0.13 Ba | 3.25 ± 0.03 ABab | * | |
| 1–2 mm | 3.25 ± 0.07 Aab | 3.28 ± 0.05 Ab | 3.24 ± 0.05 Aa | 3.25 ± 0.12 Aab | ns | |
| 0.25–1 mm | 3.23 ± 0.06 Bb | 3.41 ± 0.02 Aa | 3.26 ± 0.10 Ba | 3.40 ± 0.05 Aa | * | |
| <0.25 mm | 3.32 ± 0.09 Aab | 3.38 ± 0.07 Aa | 3.26 ± 0.05 Aa | 3.29 ± 0.07 Aab | ns | |
| p value | ns | *** | ns | ns | ||
| Simpson (D) | > 5 mm | 0.94 ± 0.01 Aa | 0.93 ± 0.006 Bb | 0.95 ± 0.001 Aa | 0.94 ± 0.006 ABa | * |
| 2–5 mm | 0.95 ± 0.001 Aa | 0.96 ± 0.01 Aa | 0.96 ± 0.01 Aa | 0.95 ± 0.005 Aa | ns | |
| 1–2 mm | 0.95 ± 0.005 Aa | 0.94 ± 0.006 Aa | 0.95 ± 0.004 Aa | 0.94 ± 0.01 Aa | ns | |
| 0.25–1 mm | 0.94 ± 0.004 Ca | 0.95 ± 0.001 Aa | 0.95 ± 0.005 BCa | 0.95 ± 0.001 ABa | * | |
| <0.25 mm | 0.95 ± 0.006 ABa | 0.95 ± 0.004 Aa | 0.94 ± 0.002 Ba | 0.95 ± 0.005 Ba | ns | |
| p value | ns | ** | ns | ns | ||
| Margalef (M) | >5 mm | 10.79 ± 0.48 Ab | 11.25 ± 1.57 Ab | 12.04 ± 0.83 Aa | 11.83 ± 0.80 Aa | ns |
| 2–5 mm | 13.59 ± 0.05 Aa | 10.94 ± 0.43 Bb | 11.12 ± 1.66 Ba | 11.17 ± 0.92 Ba | * | |
| 1–2 mm | 11.93 ± 1.26 Aab | 12.03 ± 0.70 Aab | 10.73 ± 0.52 Aa | 12.13 ± 2.07 Aa | ns | |
| 0.25–1 mm | 11.17 ± 1.29 Bb | 13.34 ± 0.96 Aa | 10.85 ± 1.42 Ba | 13.45 ± 0.83 Aa | * | |
| <0.25 mm | 12.89 ± 0.63 Aa | 12.13 ± 0.22 Aab | 12.33 ± 0.46 Aa | 12.82 ± 0.90 Aa | ns | |
| p value | * | ns | ns | ns | ||
| Index | >5 mm | 2–5 mm | 1–2 mm | 0.25–1 mm | <0.25 mm |
|---|---|---|---|---|---|
| Total PLFA | −0.14 | −0.44 | −0.22 | 0.01 | 0.003 |
| Actinomycetes | −0.17 | −0.21 | −0.3 | −0.23 | −0.13 |
| Bacteria | 0.02 | −0.41 | −0.05 | 0.22 | 0.14 |
| Fungi | −0.53 | −0.43 | −0.64 * | −0.48 | −0.17 |
| AM | −0.67 * | −0.64 * | −0.68 * | −0.56 | −0.46 |
| F/B | −0.76 ** | −0.07 | −0.73 ** | −0.66 * | −0.45 |
| G+/G− | −0.17 | 0.04 | 0.33 | −0.32 | −0.37 |
| H′ | −0.27 | −0.75** | −0.27 | 0.61 * | −0.04 |
| D | −0.43 | 0.47 | −0.44 | 0.56 | 0.03 |
| M | 0.06 | −0.74 ** | −0.14 | 0.50 | −0.49 |
| BG | 0.12 | 0.17 | −0.28 | 0.56 | 0.28 |
| NAG | −0.03 | 0.54 | −0.37 | 0.26 | −0.57 |
| BXYL | 0.05 | 0.83 *** | −0.63 * | 0.65 * | −0.41 |
| LAP | −0.51 | 0.8 ** | −0.32 | 0.66 * | −0.48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, J.; Li, S.; Liang, G.; Wu, X.; Zhang, Q.; Gao, C.; Li, J.; Jin, D.; Zheng, F.; Zhang, M.; et al. The Contribution of Microorganisms to Soil Organic Carbon Accumulation under Fertilization Varies among Aggregate Size Classes. Agronomy 2021, 11, 2126. https://doi.org/10.3390/agronomy11112126
Lu J, Li S, Liang G, Wu X, Zhang Q, Gao C, Li J, Jin D, Zheng F, Zhang M, et al. The Contribution of Microorganisms to Soil Organic Carbon Accumulation under Fertilization Varies among Aggregate Size Classes. Agronomy. 2021; 11(11):2126. https://doi.org/10.3390/agronomy11112126
Chicago/Turabian StyleLu, Jinjing, Shengping Li, Guopeng Liang, Xueping Wu, Qiang Zhang, Chunhua Gao, Jianhua Li, Dongsheng Jin, Fengjun Zheng, Mengni Zhang, and et al. 2021. "The Contribution of Microorganisms to Soil Organic Carbon Accumulation under Fertilization Varies among Aggregate Size Classes" Agronomy 11, no. 11: 2126. https://doi.org/10.3390/agronomy11112126
APA StyleLu, J., Li, S., Liang, G., Wu, X., Zhang, Q., Gao, C., Li, J., Jin, D., Zheng, F., Zhang, M., Abdelrhman, A. A., & Degré, A. (2021). The Contribution of Microorganisms to Soil Organic Carbon Accumulation under Fertilization Varies among Aggregate Size Classes. Agronomy, 11(11), 2126. https://doi.org/10.3390/agronomy11112126