
Management of Iron and Manganese Toxicities of Lentil Crops Grown in Central Chile

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Sampling of Plant Material and Soils
2.2. Pot Experiment
2.3. Data Analysis
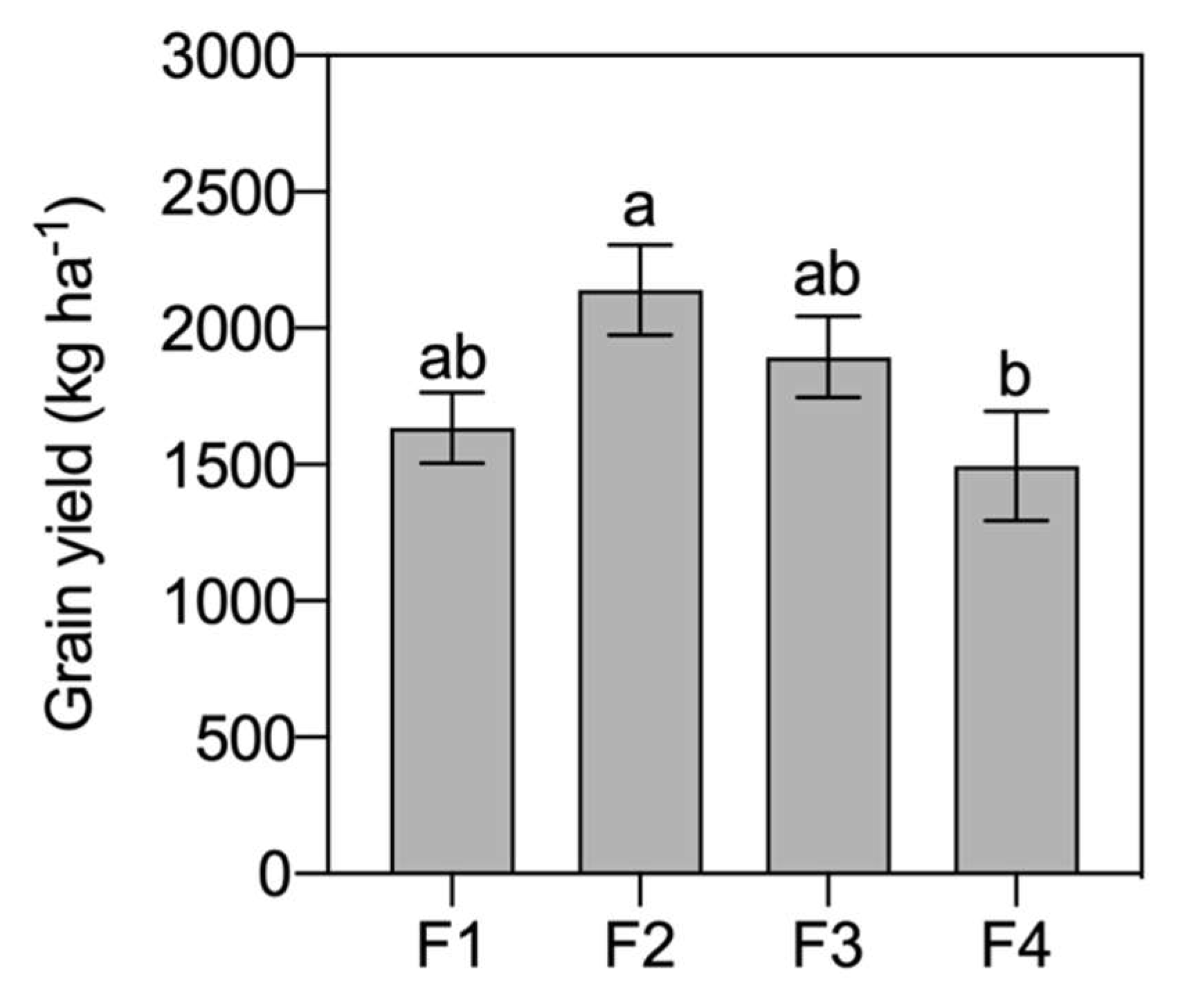
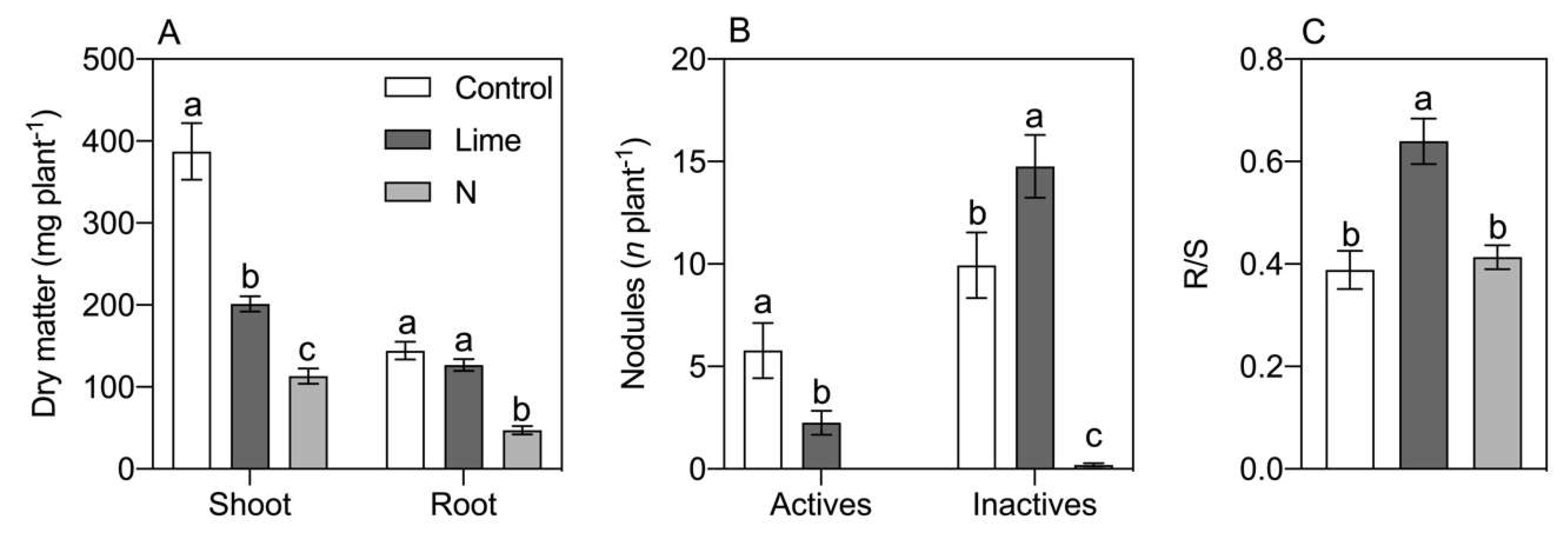
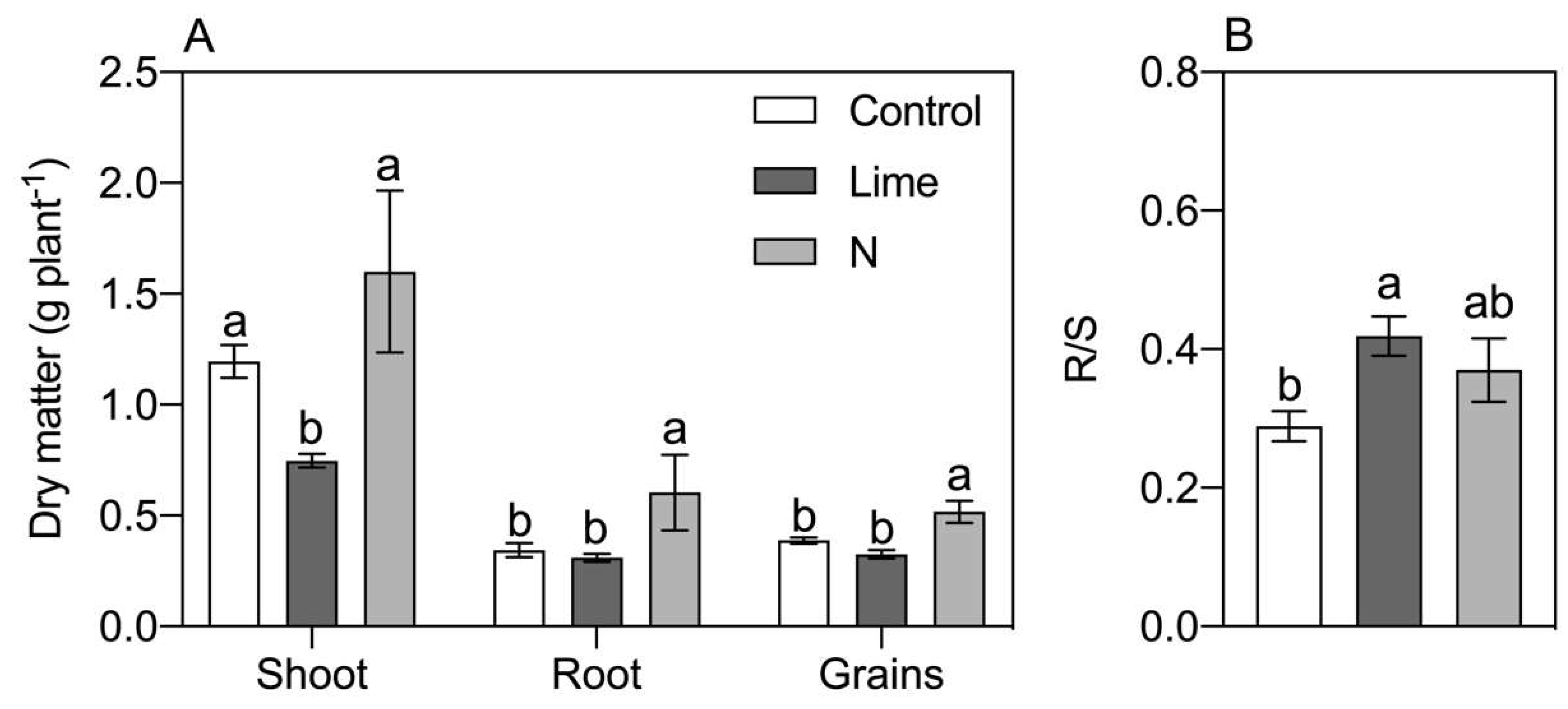
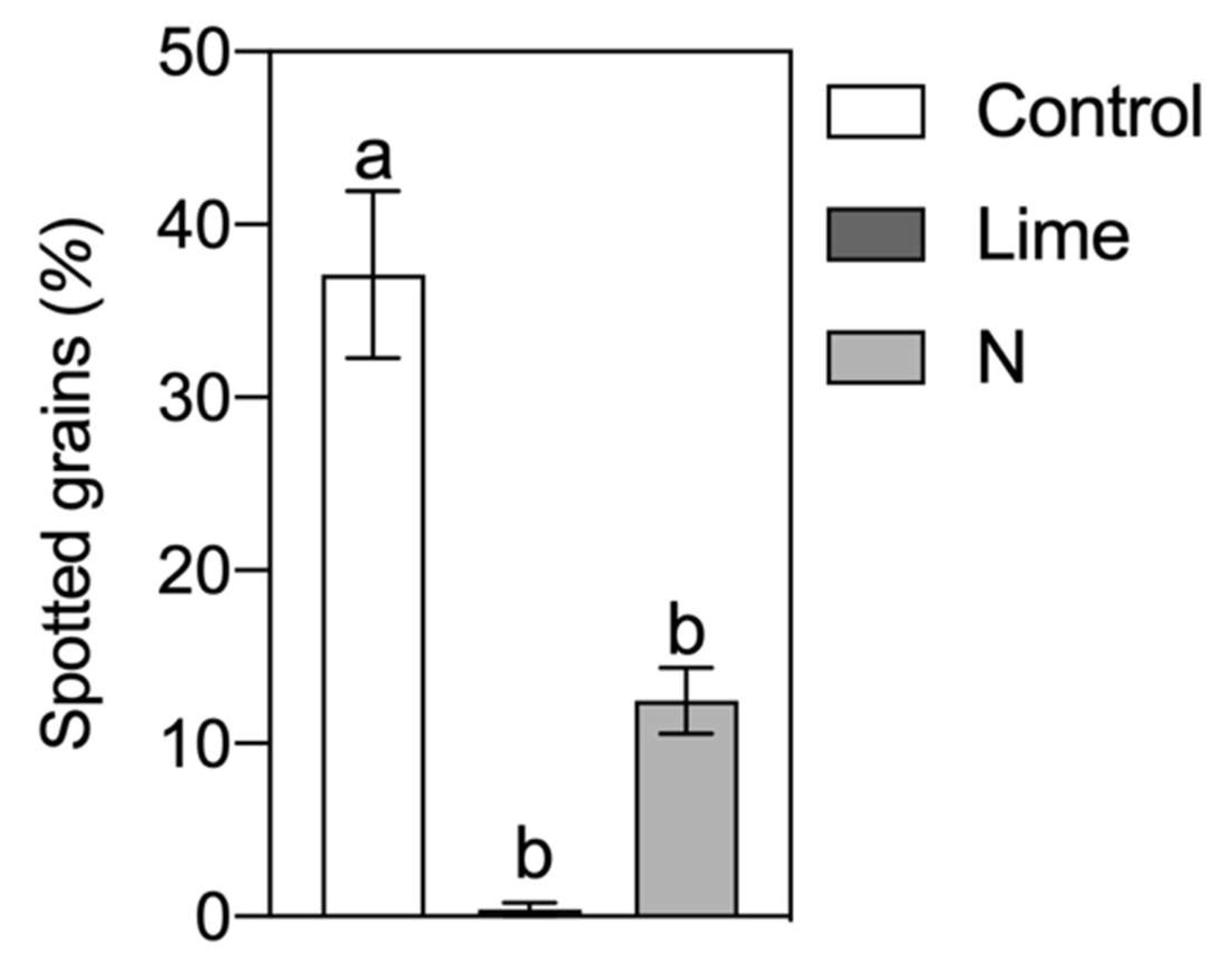
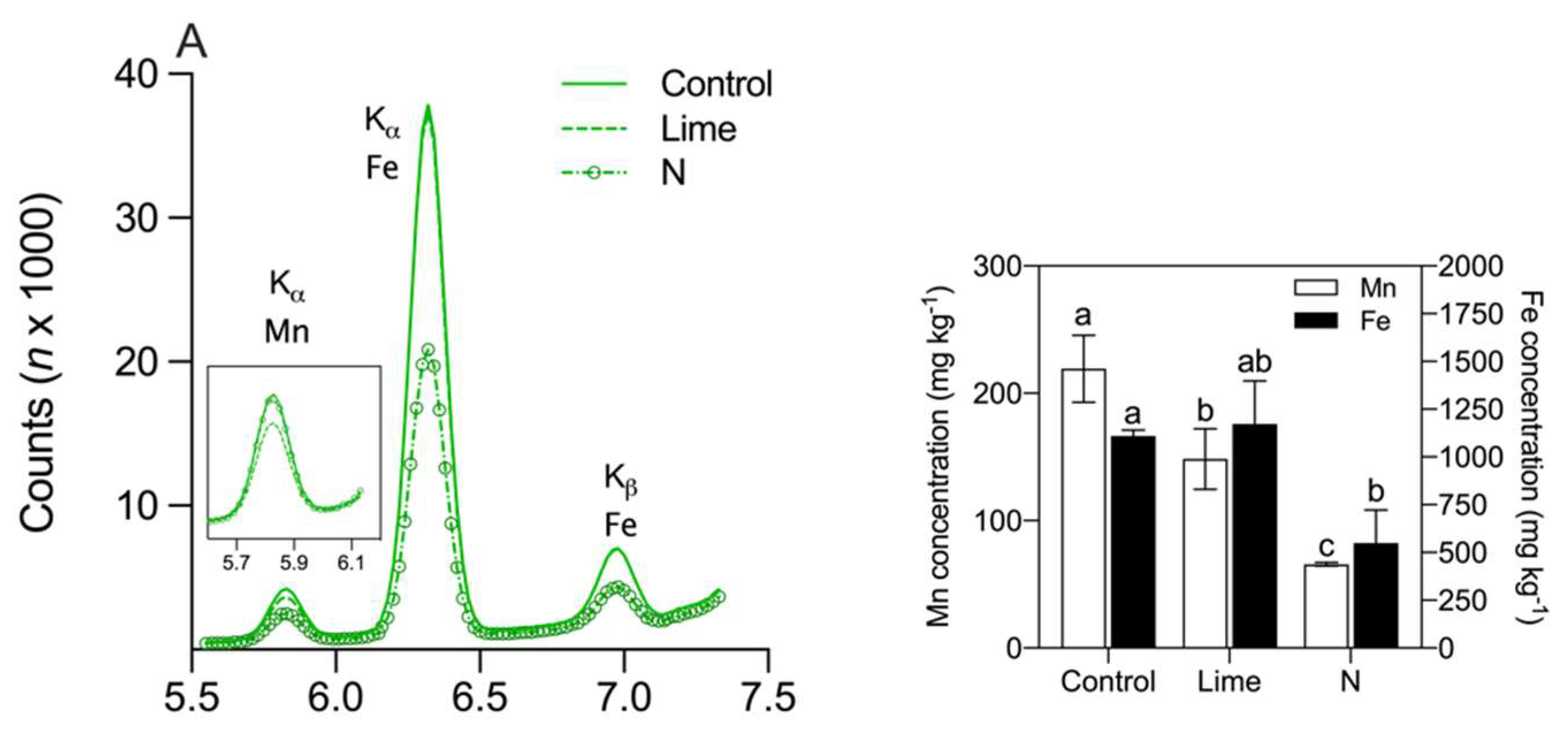
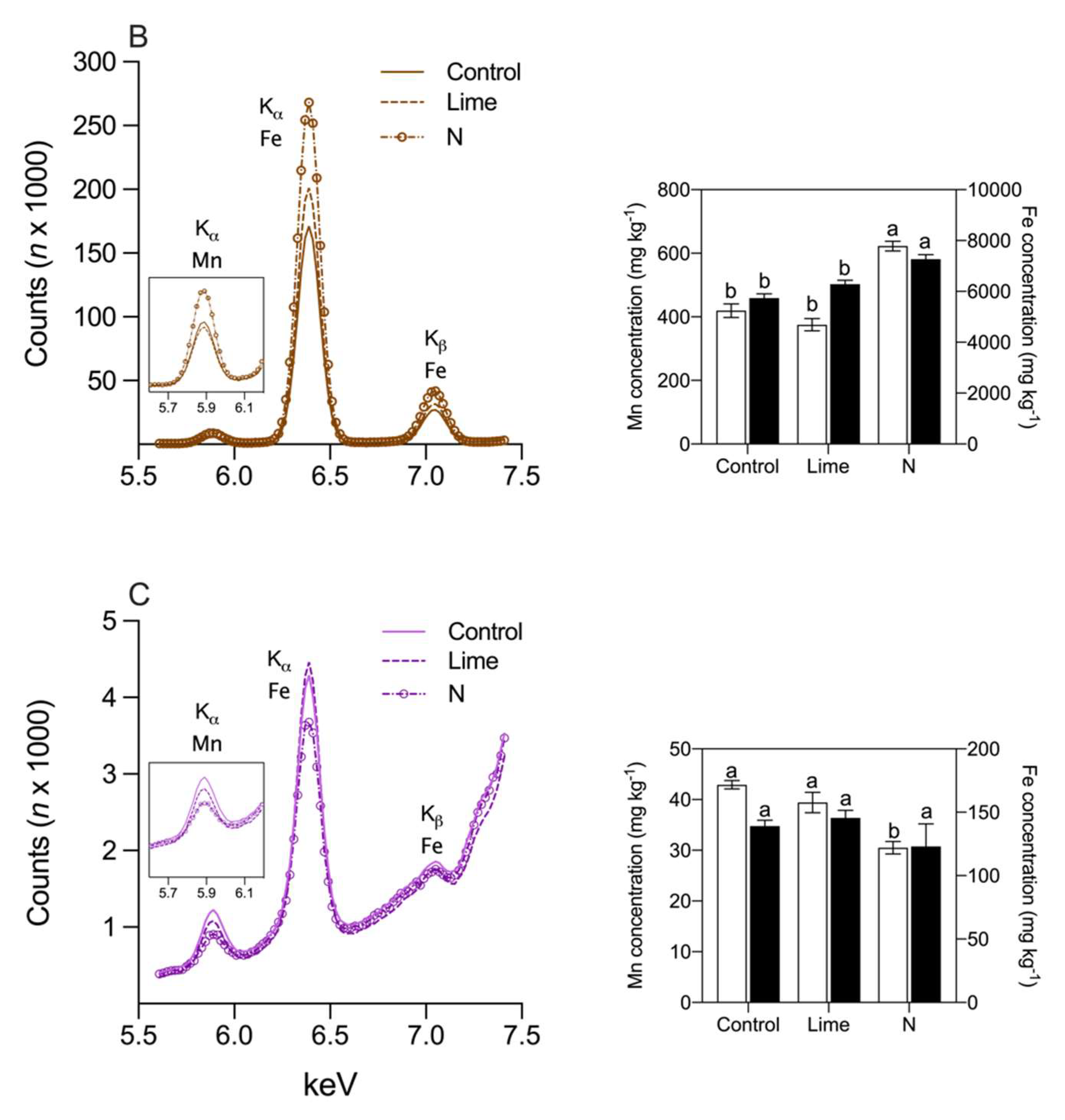
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Casanova, M.; Salazar, O.; Seguel, O.; Luzio, W. The Soils of Chile; World Soils Book Series; Springer: Dordrecht, The Netherlands, 2013; ISBN 978-94-007-5948-0. [Google Scholar]
- Sepúlveda-Caamaño, M.; Gerding, M.; Vargas, M.; Moya-Elizondo, E.; Oyarzúa, P.; Campos, J. Lentil (Lens Culinaris L.) Growth Promoting Rhizobacteria and Their Effect on Nodulation in Coinoculation with Rhizobia. Arch. Agron. Soil Sci. 2018, 64, 244–256. [Google Scholar] [CrossRef]
- Tay, J.; France, A.; Paredes, M. Súper Araucana-INIA: Una Nueva Variedad de Lenteja (Lens Culinaris Med.) Chilena de Grano Grande. Agric. Tec. 2001, 61, 385–389. [Google Scholar] [CrossRef]
- Peñaloza, E.; Tay, J.; France, A. Calpún-INIA, Cultivar de Lenteja (Lens Culinaris Medik.) de Grano Grande y Resistente a Roya. Agric. Tec. 2007, 67, 68–71. [Google Scholar] [CrossRef]
- Peoples, M.B.; Bowman, A.M.; Gault, R.R.; Herridge, D.F.; McCallum, M.H.; McCormick, K.M.; Norton, R.M.; Rochester, I.J.; Scammell, G.J.; Schwenke, G.D. Factors Regulating the Contributions of Fixed Nitrogen by Pasture and Crop Legumes to Different Farming Systems of Eastern Australia. Plant Soil 2001, 228, 29–41. [Google Scholar] [CrossRef]
- Pala, M.; Ryan, J.; Zhang, H.; Singh, M.; Harris, H.C. Water-Use Efficiency of Wheat-Based Rotation Systems in a Mediterranean Environment. Agric. Water Manag. 2007, 93, 136–144. [Google Scholar] [CrossRef]
- Gregory, P.J. Alternative Crops for Duplex Soils: Growth and Water Use of Some Cereal, Legume, and Oilseed Crops, and Pastures. Aust J. Agric. Res. 1998, 49, 21–32. [Google Scholar] [CrossRef]
- Sepulveda, P.; Baherle, P. Marea Negra En Lenteja. Investig. Prog. Agropecu. Quilamapu. 1986, 36, 20–23. [Google Scholar]
- France, A.; Tay, J. La Roña o Marea Negra de La Lenteja. Investig. Prog. Agropecu. Quilamapu. 1986, 29, 18–21. [Google Scholar]
- France, A.; Tay, J. Problema Nutricional En Lentejas (Lens Culinaris Med.) Asociado a Altos Contenidos de Manganeso y Hierro. Agric. Tec. 1986, 46, 379–383. [Google Scholar]
- Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press: Cambridge, MA, USA, 2012; ISBN 978-0-12-384905-2. [Google Scholar]
- Havlin, J.; Beaton, J.D.; Tisdale, S.L.; Nelson, W.L. Soil Fertility and Fertilizers: An Introduction to Nutrient Management; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2005. [Google Scholar]
- Sparks, D.L. Environmental Soil Chemistry; Elsevier: Amsterdam, The Netherlands, 2003; ISBN 978-0-08-049480-7. [Google Scholar]
- Lindsay, W. Chemical Equilibria in Soils; Wiley-Interscience: New York, NY, USA, 1979. [Google Scholar]
- Morrissey, J.; Guerinot, M.L. Iron Uptake and Transport in Plants: The Good, the Bad, and the Ionome. Chem. Rev. 2009, 109, 4553–4567. [Google Scholar] [CrossRef] [PubMed]
- Ciudad, C.; Ite, R. Disponibilidad de Hierro y Manganeso En Suelos Con Aireación Restringida. Agric. Tec. 1994, 54, 199–203. [Google Scholar]
- Hue, N.V. A Possible Mechanism for Manganese Phytotoxicity in Hawaii Soils Amended with a Low-Manganese Sewage Sludge. J. Environ. Qual. 1988, 17, 473–479. [Google Scholar] [CrossRef]
- Cao, X.; Chen, Y.; Wang, X.; Deng, X. Effects of Redox Potential and pH Value on the Release of Rare Earth Elements from Soil. Chemosphere 2001, 44, 655–661. [Google Scholar] [CrossRef]
- FAO; ITPS. Status of the World’s Soil Resources: Main Report; FAO: Rome, Italy, 2015; ISBN 978-92-5-109004-6. [Google Scholar]
- Rashid, D.A.; Ryan, J. Micronutrient Constraints to Crop Production in Soils with Mediterranean-Type Characteristics: A Review. J. Plant Nutr. 2004, 27, 959–975. [Google Scholar] [CrossRef]
- Ovalle, C.; Del Pozo, A.; Avendaño, J.; Aronson, J. Caracteristicas Fenologicas y Productivas de Treinta y Cuatro Accesiones de Hualputra (Medicago Polymorpha) Colectadas En La Zona Mediterranea de Chile. Agric. Tec. 1997, 57, 261–271. [Google Scholar]
- Husson, O. Redox Potential (Eh) and pH as Drivers of Soil/Plant/Microorganism Systems: A Transdisciplinary Overview Pointing to Integrative Opportunities for Agronomy. Plant Soil 2013, 362, 389–417. [Google Scholar] [CrossRef]
- Sen Gupta, D.; McPhee, K.; Kumar, S. Development of Molecular Markers for Iron Metabolism Related Genes in Lentil and Their Expression Analysis under Excess Iron Stress. Front Plant Sci. 2017, 8, 579. [Google Scholar] [CrossRef]
- Zuo, Y.; Zhang, F. Soil and Crop Management Strategies to Prevent Iron Deficiency in Crops. Plant Soil 2011, 339, 83–95. [Google Scholar] [CrossRef]
- Marschner, H.; Römheld, V.; Kissel, M. Different Strategies in Higher Plants in Mobilization and Uptake of Iron. J. Plant Nutr. 1986, 9, 695–713. [Google Scholar] [CrossRef]
- Marschner, H.; Römheld, V.; Horst, W.J.; Martin, P. Root-Induced Changes in the Rhizosphere: Importance for the Mineral Nutrition of Plants. Z Pflanz Bodenkd. 1986, 149, 441–456. [Google Scholar] [CrossRef]
- Jensen, E.S.; Hauggaard-Nielsen, H. How Can Increased Use of Biological N2 Fixation in Agriculture Benefit the Environment? Plant Soil 2003, 252, 177–186. [Google Scholar] [CrossRef]
- Sadzawka, A.; Carrasco, M.A.; Gez, R.; Mora, M.D.L.L.; Flores, H.; Neaman, A. Metodos de Análisis Recomendados Para Los Suelos de Chile. Revision 2006; Instituto de Investigaciones Agropecuarias: Santiago, Chile, 2006. [Google Scholar]
- Nelson, D.; Sommers, L. Total carbon, organic carbon and organic matter. In Methods of Soil Analysis; Page, A., Miller, R., Keeney, D., Eds.; ASA, SSSA, CSSA: Madison, WI, USA, 1982; pp. 539–580. [Google Scholar]
- Lindsay, W.L.; Norvell, W.A. Development of a DTPA Soil Test for Zinc, Iron, Manganese, and Copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- CIREN. Descripción de Suelos, Materiales y Símbolos, Estudio Agrológico VII Región; CIREN: Santiago, Chile, 1997. [Google Scholar]
- Tennant, D. A Test of a Modified Line Intersect Method of Estimating Root Length. J. Ecol. 1975, 63, 995–1001. [Google Scholar] [CrossRef]
- Dijk, D. van Wageningen Evaluating Programmes for Analytical Laboratories (WEPAL): A World of Experience. Commun in Soil Sci. Plant Anal. 2002, 33, 2457–2465. [Google Scholar] [CrossRef]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Mahmoudi, H.; Ksouri, R.; Gharsalli, M.; Lachaâl, M. Differences in Responses to Iron Deficiency between Two Legumes: Lentil (Lens Culinaris) and Chickpea (Cicer Arietinum). J. Plant Physiol. 2005, 162, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Gupta, U.C.; Wu, K.; Liang, S. Micronutrients in Soils, Crops, and Livestock. Earth Sci. Front 2008, 15, 110–125. [Google Scholar] [CrossRef]
- Rosales-Rodríguez, J.A.; Esquivel-Segura, E.A.; Acevedo-Tapia, M.A.; González-Ortega, M.; Cartes-Rodríguez, E. Pre and Post-Fire Situation, of an Ecosystem of the Roble-Hualo Forest Type, Maule Region, Chile. Rev. For. Mes. Kurú. 2019, 16, 55–68. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C.; Clark, R.B. Micronutrients in Crop Production. Adv. Agron. 2002, 77, 185–268. [Google Scholar] [CrossRef]
- Ray, H.; Bett, K.; Tar’an, B.; Vandenberg, A.; Thavarajah, D.; Warkentin, T. Mineral Micronutrient Content of Cultivars of Field Pea, Chickpea, Common Bean, and Lentil Grown in Saskatchewan, Canada. Crop Sci. 2014, 54, 1698–1708. [Google Scholar] [CrossRef]
- Streeter, J. Inhibition of Legume Nodule Formation and N2 Fixation by Nitrate. Crit. Rev. Plant Sci. 1988, 7, 1–23. [Google Scholar] [CrossRef]
- Cabeza, R.A.; Koester, B.; Liese, R.; Lingner, A.; Baumgarten, V.; Dirks, J.; Salinas-Riester, G.; Pommerenke, C.; Dittert, K.; Schulze, J. An RNA Sequencing Transcriptome Analysis Reveals Novel Insights into Molecular Aspects of the Nitrate Impact on the Nodule Activity of Medicago Truncatula. Plant Physiol. 2014, 164, 400–411. [Google Scholar] [CrossRef]
- Schulze, J.; Liese, R.; Ballesteros, G.; Casieri, L.; Salinas, G.; Cabeza, R.A. Ammonium Acts Systemically While Nitrate Exerts an Additional Local Effect on Medicago Truncatula Nodules. Plant Sci. 2020, 292, 110383. [Google Scholar] [CrossRef]
- Schilling, G.; Adgo, E.; Schulze, J. Carbon Costs of Nitrate Reduction in Broad Bean (Vicia Faba L.) and Pea (Pisum Sativum L.) Plants. J. Plant Nutr. Soil Sci. 2006, 169, 691–698. [Google Scholar] [CrossRef]
- Foy, C.D.; Chaney, R.L.; White, M.C. The Physiology of Metal Toxicity in Plants. Annu. Rev. Plant Physiol. 1978, 29, 511–566. [Google Scholar] [CrossRef]
- Hell, R.; Stephan, U.W. Iron Uptake, Trafficking and Homeostasis in Plants. Planta 2003, 216, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Blamey, F.P.C.; Joyce, D.C.; Edwards, D.G.; Asher, C.J. Role of Trichomes in Sunflower Tolerance to Manganese Toxicity. Plant Soil 1986, 91, 171–180. [Google Scholar] [CrossRef]
- Page, V.; Weisskopf, L.; Feller, U. Heavy Metals in White Lupin: Uptake, Root-to-Shoot Transfer and Redistribution within the Plant. New Phytol. 2006, 171, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Horst, W.J. Factors Responsible for Genotypic Manganese Tolerance in Cowpea (Vigna Unguiculata). Plant Soil 1983, 72, 213–218. [Google Scholar] [CrossRef]
- Blamey, F.P.C.; Paterson, D.J.; Walsh, A.; Afshar, N.; McKenna, B.A.; Cheng, M.; Tang, C.; Horst, W.J.; Menzies, N.W.; Kopittke, P.M. Time-Resolved X-Ray Fluorescence Analysis of Element Distribution and Concentration in Living Plants: An Example Using Manganese Toxicity in Cowpea Leaves. Environ. Exper. Bot. 2018, 156, 151–160. [Google Scholar] [CrossRef]
- González, A.; Lynch, J.P. Subcellular and Tissue Mn Compartmentation in Bean Leaves under Mn Toxicity Stress. Funct. Plant Biol. 1999, 26, 811–822. [Google Scholar] [CrossRef]
- Shigaki, T.; Pittman, J.K.; Hirschi, K.D. Manganese Specificity Determinants in the Arabidopsis Metal/H+ Antiporter CAX2. J. Biol. Chem. 2003, 278, 6610–6617. [Google Scholar] [CrossRef] [PubMed]
- Lanquar, V.; Ramos, M.S.; Lelièvre, F.; Barbier-Brygoo, H.; Krieger-Liszkay, A.; Krämer, U.; Thomine, S. Export of Vacuolar Manganese by AtNRAMP3 and AtNRAMP4 Is Required for Optimal Photosynthesis and Growth under Manganese Deficiency. Plant Physiol. 2010, 152, 1986–1999. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H.; Römheld, V. In Vivo Measurement of Root-Induced pH Changes at the Soil-Root Interface: Effect of Plant Species and Nitrogen Source. Z Pflanz. 1983, 111, 241–251. [Google Scholar] [CrossRef]
- Döbereiner, J. Manganese Toxicity Effects on Nodulation and Nitrogen Fixation of Beans (Phaseolus Vulgaris L.), in Acid Soils. Plant Soil 1966, 24, 153–166. [Google Scholar] [CrossRef]
- Bordeleau, L.M.; Prévost, D. Nodulation and Nitrogen Fixation in Extreme Environments. Plant Soil 1994, 161, 115–125. [Google Scholar] [CrossRef]
- Cabeza, R.A.; Liese, R.; Fischinger, S.A.; Sulieman, S.; Avenhaus, U.; Lingner, A.; Hein, H.; Koester, B.; Baumgarten, V.; Dittert, K.; et al. Long-Term Non-Invasive and Continuous Measurements of Legume Nodule Activity. Plant J. 2015, 81, 637–648. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site and Geographic Location | * Sowing, Harvest and Threshing Date | Previous Crops | Sowing Surface | Nutrient Applied (kg ha−1) | |||
|---|---|---|---|---|---|---|---|
| (ha) | N | P | K | S | |||
| Farmer 1 35°42′44.6″ S 72°31′07.0″ W | 22 June 7 December 25 December | L/P/W | 0.5 | - | 30 | 41.5 | 18 |
| Farmer 2 35°42′44.6″ S 72°31′07.0″ W | 22 June 7 December 25 December | W | 0.9 | - | 30 | 41.5 | 18 |
| Farmer 3 35°41′38.8″ S 72°30′10.6″ W | 24 June 7 December 25 December | W | 2.0 | 46 | - | - | - |
| Farmer 4 35°40′50.1″ S 72°29′12.1″ W | 24 June 7 December 25 December | W | 2.0 | - | - | - | - |
| pHw | EC | SOM | K | Ca | Mg | Na | N | P | Fe | Mn | Zn | Cu | B | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| dS/m | g 100 g−1 | cmol(+) kg−1 | mg kg−1 | |||||||||||
| F1, F2 | 5.7 | 0.14 | 4.8 | 0.92 | 7.87 | 2.73 | 0.14 | 11.8 | 15.9 | 100.1 | 16.7 | 3.42 | 1.24 | 0.74 |
| F3 | 5.7 | 0.06 | 3.5 | 0.57 | 8.26 | 1.84 | 0.14 | 13.0 | 34.0 | 134.3 | 16.2 | 3.09 | 1.94 | 0.29 |
| F4 | 5.8 | 0.05 | 1.8 | 0.18 | 5.82 | 3.14 | 0.20 | 1.0 | 11.0 | 90.2 | 19.0 | 1.14 | 1.34 | 0.12 |
| Sites | N | P | K | Ca | Mg | B | Cu | Zn | Mn | Fe |
|---|---|---|---|---|---|---|---|---|---|---|
| g 100 g−1 | mg kg−1 | |||||||||
| F1 | 3.77 (0.19) | 0.31 (0.04) b | 0.77 (0.04) | 0.06 (0.02) b | 0.11 (0.01) | 5.75 (0.96) | 6.75 (0.96) | 41.00 (4.97) | 15.50 (2.65) b | 49.25 (5.56) b |
| F2 | 3.86 (0.48) | 0.34 (0.10) b | 0.82 (0.10) | 0.05 (0.03) b | 0.12 (0.02) | 4.75 (1.26) | 8.25 (1.26) | 45.50 (9.57) | 14.00 (0.82) b | 53.50 (5.45) b |
| F3 | 3.86 (0.08) | 0.48 (0.05) a | 0.86 (0.15) | 0.07 (0.01) ab | 0.13 (0.01) | 13.25 (10.97) | 6.75 (0.96) | 47.00 (3.92) | 29.75 (6.24) a | 97.25 (10.28) a |
| F4 | 3.64 (0.07) | 0.30 (0.02) b | 0.69 (0.01) | 0.10 (0.02) a | 0.11 (0.01) | 5.75 (1.50) | 7.50 (0.58) | 44.25 (0.58) | 16.50 (1.73) b | 57.75 (4.11) b |
| p-Value | 0.6 | <0.01 | 0.1 | <0.05 | 0.1 | 0.1 | 0.1 | 0.5 | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amigo, R.; Méndez-Espinoza, A.M.; Schwember, A.R.; Cornejo, J.; Baettig, R.; Cabeza, R.A. Management of Iron and Manganese Toxicities of Lentil Crops Grown in Central Chile. Agronomy 2021, 11, 2051. https://doi.org/10.3390/agronomy11102051
Amigo R, Méndez-Espinoza AM, Schwember AR, Cornejo J, Baettig R, Cabeza RA. Management of Iron and Manganese Toxicities of Lentil Crops Grown in Central Chile. Agronomy. 2021; 11(10):2051. https://doi.org/10.3390/agronomy11102051
Chicago/Turabian StyleAmigo, Ramón, Ana María Méndez-Espinoza, Andrés R. Schwember, Jorge Cornejo, Ricardo Baettig, and Ricardo A. Cabeza. 2021. "Management of Iron and Manganese Toxicities of Lentil Crops Grown in Central Chile" Agronomy 11, no. 10: 2051. https://doi.org/10.3390/agronomy11102051
APA StyleAmigo, R., Méndez-Espinoza, A. M., Schwember, A. R., Cornejo, J., Baettig, R., & Cabeza, R. A. (2021). Management of Iron and Manganese Toxicities of Lentil Crops Grown in Central Chile. Agronomy, 11(10), 2051. https://doi.org/10.3390/agronomy11102051