
Cytological and Molecular Characterization for Ploidy Determination in Yams (Dioscorea spp.)

 , , , , , ,
, , , , , ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
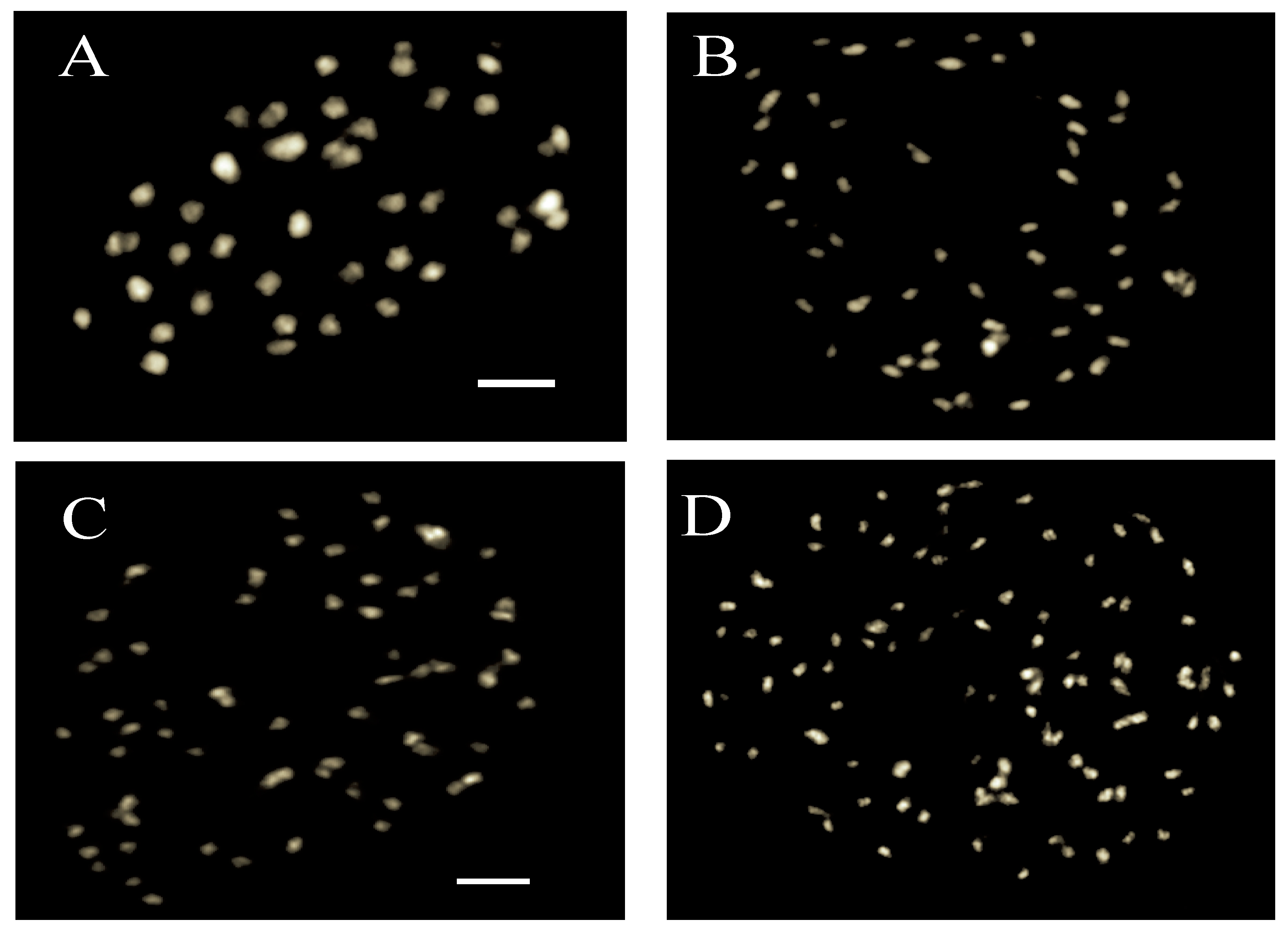
2.2. Chromosome Counting
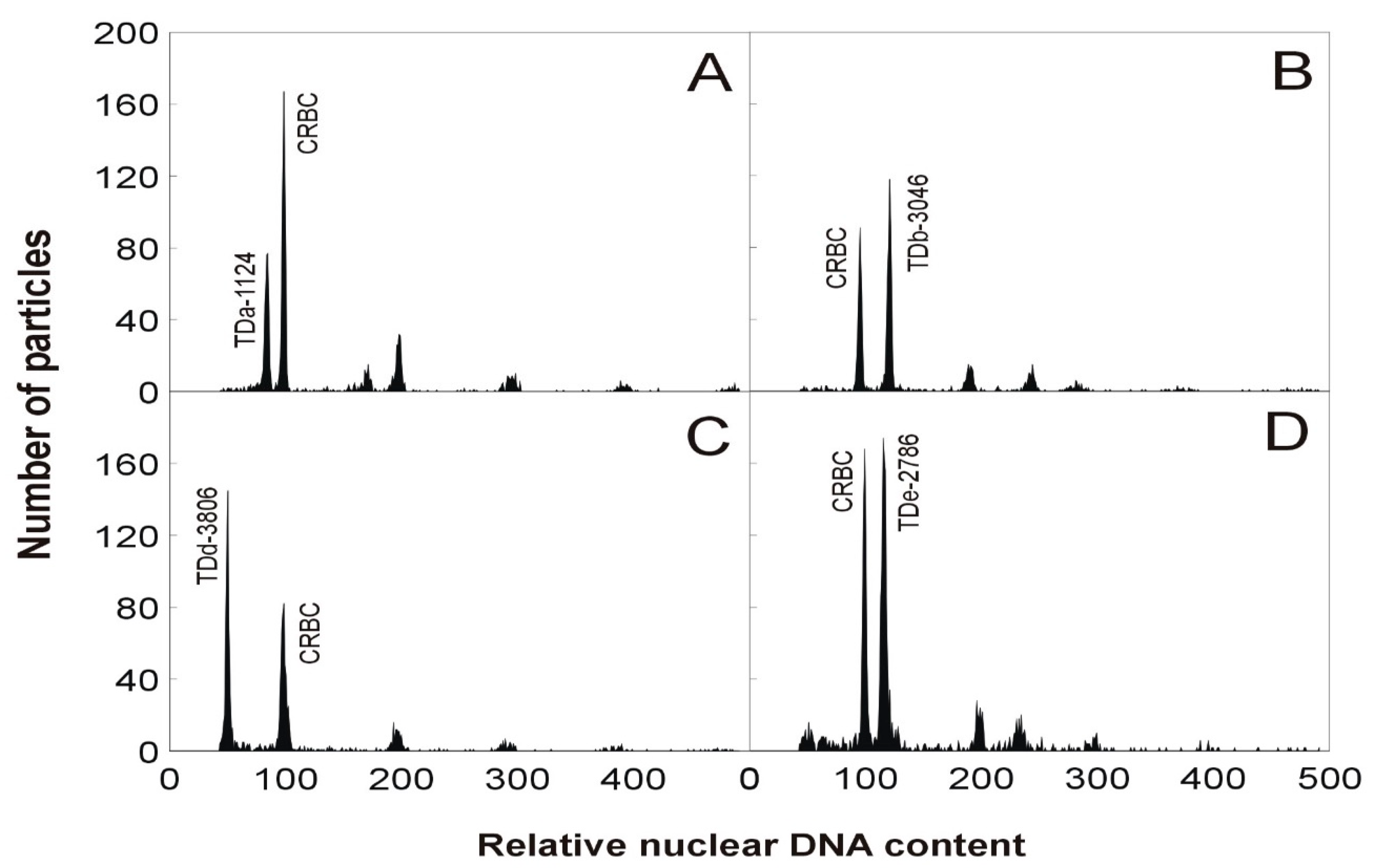
2.3. Flow Cytometry
2.4. DNA Extraction and Quantification
2.5. Analysis of Molecular Data Using DArTseq SNP Markers
3. Results
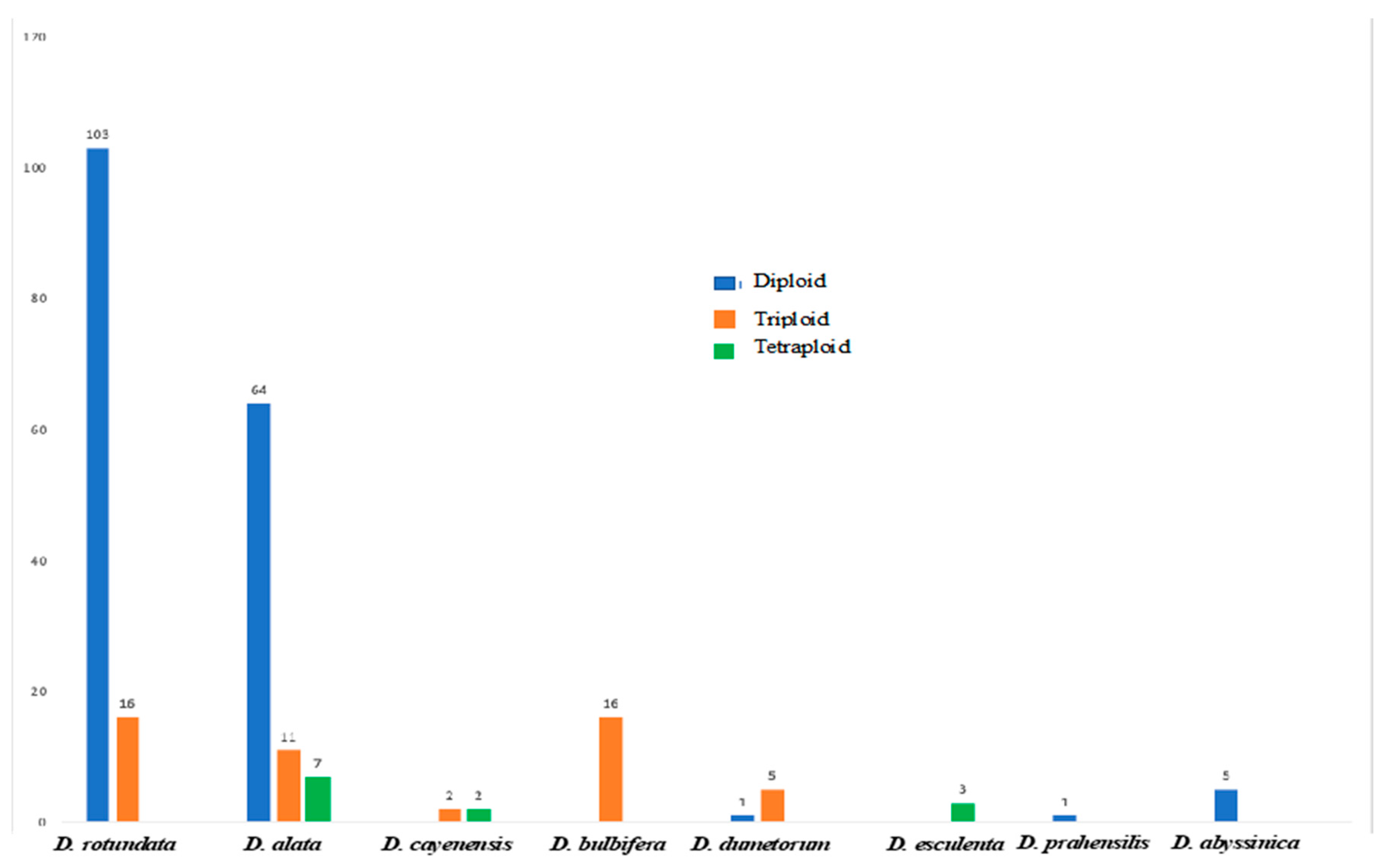
3.1. Ploidy Estimation Using Chromosome Counting
3.2. Flow Cytometry Analysis of Dioscorea Accessions
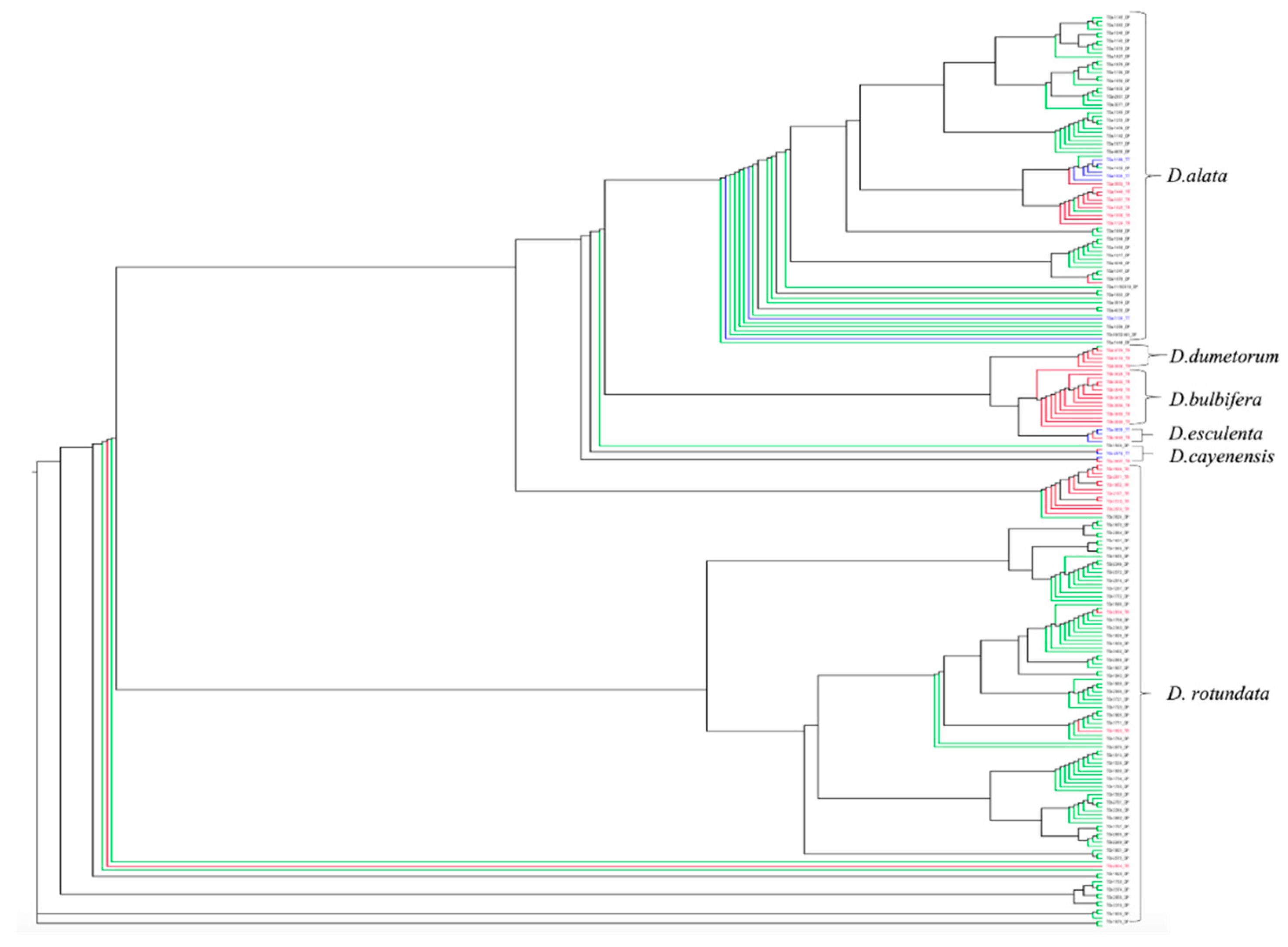
3.3. Ploidy Status and Phylogenetic Relationship of Dioscorea spp. Based on DArTseq SNP Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, L.; Xu, J.; Luo, L.; Hu, H.; Meng, X.; Li, X.; Chen, S. Predicting the potential global distribution of diosgenin-contained Dioscorea species. Chin. Med. 2018, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- Sonibare, M.A.; Asiedu, R.; Albach, D.C. Genetic diversity of Dioscorea dumetorum (Kunth) Pax using amplified fragment length polymorphisms (AFLP) and cpDNA. Biochem. Syst. Ecol. 2010, 38, 320–334. [Google Scholar] [CrossRef]
- Darkwa, K.; Olasanmi, B.; Asiedu, R.; Asfaw, A. Review of empirical and emerging breeding methods and tools for yam (Dioscorea spp.) improvement: Status and prospects. Plant Breed. 2020, 139, 474–497. [Google Scholar] [CrossRef]
- Asiedu, R.; Sartie, A. Crops that feed the world 1. Yams: Yams for income and food security. Food Secur. 2010, 2, 305–315. [Google Scholar] [CrossRef]
- Price, E.J.; Bhattacharjee, R.; Lopez-Montes, A.; Fraser, P.D. Metabolite profiling of yam (Dioscorea spp.) accessions for use in crop improvement programmes. Metabolomics 2017, 13, 1–12. [Google Scholar] [CrossRef]
- FAO. Food and Agriculture Organization of the United Nations Statistics Database, FAOSTAT. 2017. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 10 March 2020).
- Alabi, T.R.; Adebola, P.O.; Asfaw, A.; De Koeyer, D.; Lopez-Montes, A.; Asiedu, R. Spatial multivariate cluster analysis for defining target population of environments in West Africa for yam breeding. Int. J. Appl. Geospa. Res. 2019, 10, 1–30. [Google Scholar] [CrossRef]
- Baimey, H.; Coyne, D.; Labuschagne, N. Effect of fertiliser application on yam nematode (Scutellonema bradys) multiplication and consequent damage to yam (Dioscorea spp.) under field and storage conditions in Benin. Int. J. Pest Manag. 2006, 52, 63–70. [Google Scholar] [CrossRef]
- Mignouna, H.D.; Abang, M.M.; Asiedu, R. Genomics of yams, a common source of food and medicine in the tropics. Gen. Trop. Crop Plants. 2008, 549–570. [Google Scholar] [CrossRef]
- Girma, G.; Natsume, S.; Carluccio, A.V.; Takagi, H.; Matsumura, H.; Uemura, A.; Muranaka, S.; Takagi, H.; Stavolone, L.; Gedil, M.; et al. Identification of candidate flowering and sex genes in white Guinea yam (D. rotundata Poir.) by SuperSAGE transcriptome profiling. PLoS ONE 2019, 14, e0216912. [Google Scholar] [CrossRef] [PubMed]
- Siadjeu, C.; Pucker, B.; Viehöver, P.; Albach, D.C.; Weisshaar, B. High contiguity de novo genome sequence assembly of Trifoliate yam (Dioscorea dumetorum) using long read sequencing. Genes 2020, 11, 274. [Google Scholar] [CrossRef] [PubMed]
- Girma, G.; Hyma, K.E.; Asiedu, R.; Mitchell, S.E.; Gedil, M.; Spillane, C. Next generation sequencing based genotyping, cytometry and phenotyping for understanding diversity and evolution of guinea yams. Theor. Appl. Genet. 2014, 127, 1783–1794. [Google Scholar] [CrossRef]
- Pickering, R.A.; Hayes, J.D. Partial incompatibility in crosses between Hordeum vulgare L. and H. bulbosum L. Euphytica 1976, 25, 671–678. [Google Scholar] [CrossRef]
- Abraham, K.; Nair, P.G. Polyploidy and sterility in relation to sex in Dioscorea alata L. (Dioscoreaceae). Genetica 1991, 83, 93–97. [Google Scholar] [CrossRef]
- Gamiette, F.; Bakry, F.; Ano, G. Ploidy determination of some yam species (Dioscorea spp.) by flow cytometry and conventional chromosomes counting. Genet. Resour. Crop Evol. 1999, 46, 19–27. [Google Scholar] [CrossRef]
- Egesi, C.N.; Pillay, M.; Asiedu, R.; Egunjobi, J.K. Ploidy analysis in water yam, Dioscorea alata L. germplasm. Euphytica 2002, 128, 225–230. [Google Scholar] [CrossRef]
- Sattler, M.C.; Carvalho, C.R.; Clarindo, W.R. The polyploidy and its key role in plant breeding. Planta 2016, 243, 281–296. [Google Scholar] [CrossRef]
- Malapa, R.; Arnau, G.; Noyer, J.L.; Lebot, V. Genetic diversity of the greater yam (Dioscorea alata L.) and relatedness to D. nummularia Lam. and D. transversa Br. as revealed with AFLP markers. Genet. Resour. Crop Evol. 2005, 52, 919–929. [Google Scholar] [CrossRef]
- Arnau, G.; Abraham, K.; Sheela, M.N.; Chair, H.; Sartie, A.; Asiedu, R. Yams. Root and Tuber Crops. In Handbook of Plant Breeding; Springer: New York, NY, USA, 2010; Volume 7, pp. 127–148. [Google Scholar]
- Zlesak, D.C. Pollen diameter and guard cell length as predictors of ploidy in diverse rose cultivars, species and breeding lines. J. Floric. Ornam. Biotech. 2009, 3, 53–70. [Google Scholar]
- Chambers, A.H.; Pollard, H.; Folta, K.M. Limitations of morphological ploidy estimation methods in Fragaria. J. Berry Res. 2013, 3, 135–149. [Google Scholar] [CrossRef]
- Dansi, A.; Mignouna, H.D.; Pillay, M.; Zok, S. Ploidy variation in the cultivated yams (Dioscorea cayenensis-Dioscorea rotundata complex) from Cameroon as determined by flow cytometry. Euphytica 2001, 119, 301–307. [Google Scholar] [CrossRef]
- Obidiegwu, J.; Rodriguez, E.; Ene-Obong, E.; Loureiro, J.; Muoneke, C.; Santos, C.; Kolesnikova-Allen, M.; Asiedu, R. Ploidy levels of Dioscorea alata L. germplasm determined by flow cytometry. Genet. Resour. Crop Evol. 2010, 57, 351–356. [Google Scholar] [CrossRef]
- Suda, J.; Trávníček, P. Reliable DNA ploidy determination in dehydrated tissues of vascular plants by DAPI flow cytometry-new prospects for plant research. Cytom. Part A 2006, 69, 273–280. [Google Scholar] [CrossRef]
- Delaat, A.M.M.; Gohde, W.; Vogelzakg, M.J.D.C. Determination of ploidy of single plants and plant populations by flow cytometry. Plant Breed. 1987, 99, 303–307. [Google Scholar] [CrossRef]
- Dolezel, J. Application of flow cytometry for the study of plant genomes. J. Appl. Genet. 1997, 3, 285–302. [Google Scholar]
- Besnard, G.; Baali-Cherif, D. Coexistence of diploids and triploids in a Saharan relict olive: Evidence from nuclear microsatellite and flow cytometry analyses. C. R. Biol. 2009, 332, 1115–1120. [Google Scholar] [CrossRef]
- Nemorin, A.; David, J.; Maledon, E.; Nudol, E.; Dalon, J.; Arnau, G. Microsatellite and flow cytometry analysis to help understand the origin of Dioscorea alata polyploids. Ann. Bot. 2013, 112, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Donkpegan, A.S.L.; Doucet, J.-L.; Dainou, K.; Hardy, O.J. Microsatellite development and flow cytometry in the African tree genus Afzelia (Fabaceae, Caesalpinioideae) reveal a polyploid complex. Appl. Plant Sci. 2015, 3, 1400097. [Google Scholar] [CrossRef]
- Viruel, J.; Conejero, M.; Hidalgo, O.; Pokorny, L.; Powell, R.F.; Forest, F.; Kantar, M.B.; Soto Gomez, M.; Graham, S.W.; Gravendeel, B.; et al. A target capture-based method to estimate ploidy from herbarium specimens. Front. Plant Sci. 2019, 10, 937. [Google Scholar] [CrossRef] [PubMed]
- Gardina, P.J.; Lo, K.C.; Lee, W.; Cowell, J.K.; Turpaz, Y. Ploidy status and copy number aberrations in primary glioblastomas defined by integrated analysis of allelic ratios, signal ratios and loss of heterozygosity using 500K SNP Mapping Arrays. BMC Genom. 2008, 9, 1–16. [Google Scholar] [CrossRef]
- Rungis, D.E.; Voronova, A.; Kokina, A.; Veinberga, I.; Skrabule, I.; Rostoks, N. Assessment of genetic diversity and relatedness in the Latvian potato genetic resources collection by DArT genotyping. Plant Genet. Resour. 2017, 15, 72–78. [Google Scholar] [CrossRef]
- Gatarira, C.; Paterne, A.; Matsumoto, R.; Edemondu, A.; Adetimirin, V.; Bhattacharjee, R.; Asiedu, R.; Asfaw, A. Genome-wide association analysis for tuber dry matter and oxidative browning in water yam (Dioscorea alata L.). Plants 2020, 9, 969. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233. [Google Scholar] [CrossRef]
- Otto, F. DAPI staining of fixed cells for high-resolution flow cytometry of nuclear DNA. Methods Cell Biol. 1990, 33, 105–110. [Google Scholar]
- Galbraith, D.W.; Lambert, G.M.; Macas, J.; Dolezel, J. Analysis of nuclear DNA content and ploidy in higher plants. Curr. Protoc. Cytom. 1997, 2, 7.6.1–7.6.22. [Google Scholar] [CrossRef]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Kilian, A.; Wenzl, P.; Huttner, E.; Carling, J.; Xia, L.; Blois, H.; Caig, V.; Heller-Uszynska, K.; Jaccoud, D.; Hopper, C.; et al. Diversity arrays technology: A generic genome profiling technology on open platforms. In Data Production and Analysis in Population Genomics; Humana Press: Totowa, NJ, USA, 2012; pp. 67–89. [Google Scholar]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Gompert, Z.; Mock, K.E. Detection of individual ploidy levels with genotyping-by-sequencing (GBS) analysis. Mol. Ecol. Resour. 2017, 17, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.; Daly, M.J. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2018. [Google Scholar]
- Bhattacharjee, R.; Gedil, M.; Sartie, A.; Otoo, E.; Dumet, D.; Kikuno, H.; Kumar, P.L.; Asiedu, R. Dioscorea. In Wild Crop Relatives: Genomic and Breeding Resources; Springer: Berlin/Heidelberg, Germany, 2011; pp. 71–96. [Google Scholar]
- Norman, P.E.; Tongoona, P.; Shanahan, P.E. Diversity in chromosome number and raphide morphology of yam (Dioscorea spp.) genotypes from Sierra Leone. Afr. J. Plant. Sci. 2011, 6, 157–160. [Google Scholar]
- Roux, N.; Toloza, A.; Radecki, Z.; Zapata-Arias, F.J.; Dolezel, J. Rapid detection of aneuploidy in Musa using flow cytometry. Plant Cell Rep. 2003, 21, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Doležel, J.; Lysák, M.A.; Van den Houwe, I.; Dolezelová, M.; Roux, N. Use of flow cytometry for rapid ploidy determination in Musa species. Infomusa 1997, 6, 6–9. [Google Scholar]
- Praça-Fontes, M.M.; Carvalho, C.R.; Clarindo, W.R.; Cruz, C.D. Revisiting the DNA C-values of the genome size-standards used in plant flow cytometry to choose the “best primary standards”. Plant Cell Rep. 2011, 30, 1183–1191. [Google Scholar] [CrossRef]
- Doležel, J.; Bartoš, J. Plant DNA flow cytometry and estimation of nuclear genome size. Ann. Bot. 2005, 95, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Dolezelová, M.; Doležel, J.; Roux, N.; Swennen, R. Ploidy levels revealed. InfoMusa 2005, 14, 34–36. [Google Scholar]
- Christelová, P.; De Langhe, E.; Hřibová, E.; Čížková, J.; Sardos, J.; Hušáková, M.; Sutanto, A.; Kepler, A.K.; Swennen, R.; Roux, N.; et al. Molecular and cytological characterization of the global Musa germplasm collection provides insights into the treasure of banana diversity. Biodivers. Conserv. 2017, 26, 801–824. [Google Scholar] [CrossRef]
- Rayburn, A.L.; Birdar, D.P.; Bullock, D.G.; Nelson, R.L.; Gourmet, C.; Wetzel, J.B. Nuclear DNA content diversity in Chinese soybean introductions. Ann. Bot. 1997, 80, 321–325. [Google Scholar] [CrossRef][Green Version]
- Bartoš, J.; Alkhimova, O.; Doleželová, M.; De Langhe, E.; Doležel, J. Nuclear genome size and genomic distribution of ribosomal DNA in Musa and Ensete (Musaceae): Taxonomic implications. Cytogenet. Genome Res. 2005, 109, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Čížková, J.; Hřibová, E.; Humplíková, L.; Christelová, P.; Suchánková, P.; Doležel, J. Molecular analysis and genomic organization of major DNA satellites in banana (Musa spp.). PLoS ONE 2013, 8, e54808. [Google Scholar] [CrossRef]
- Scarcelli, N.; Daïnou, O.; Agbangla, C.; Tostain, S.; Pham, J.L. Segregation patterns of isozyme loci and microsatellite markers show the diploidy of African yam Dioscorea rotundata (2n = 40). Theor. Appl. Genet. 2005, 111, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Arnau, G.; Némorin, A.; Maledon, E.; Abraham, K. Revision of ploidy status of Dioscorea alata L. (Dioscoreaceae) by cytogenetic and microsatellite segregation analysis. Theor. Appl. Genet. 2009, 118, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Bousalem, M.; Arnau, G.; Hochu, I.; Arnolin, R.; Viader, V.; Santoni, S.; David, J. Microsatellite segregation analysis and cytogenetic evidence for tetrasomic inheritance in the American yam Dioscorea trifida and a new basic chromosome number in the Dioscoreacae. Theor. Appl. Genet. 2006, 113, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Seberg, O.; Petersen, G. A critical review of concepts and methods used in classical genome analysis. Bot. Rev. 1998, 64, 372–417. [Google Scholar] [CrossRef]
- Edet, O.U.; Gorafi, Y.S.; Nasuda, S.; Tsujimoto, H. DArTseq-based analysis of genomic relationships among species of tribe Triticeae. Sci. Rep. 2018, 8, 1–11. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gatarira, C.; Sládeková, L.; Němečková, A.; Šimoníková, D.; Paliwal, R.; Asfaw, A.; Abberton, M.; Badara, G.; Asiedu, R.; Čížková, J.; et al. Cytological and Molecular Characterization for Ploidy Determination in Yams (Dioscorea spp.). Agronomy 2021, 11, 1897. https://doi.org/10.3390/agronomy11101897
Gatarira C, Sládeková L, Němečková A, Šimoníková D, Paliwal R, Asfaw A, Abberton M, Badara G, Asiedu R, Čížková J, et al. Cytological and Molecular Characterization for Ploidy Determination in Yams (Dioscorea spp.). Agronomy. 2021; 11(10):1897. https://doi.org/10.3390/agronomy11101897
Chicago/Turabian StyleGatarira, Cobes, Lucia Sládeková, Alžběta Němečková, Denisa Šimoníková, Rajneesh Paliwal, Asrat Asfaw, Michael Abberton, Gueye Badara, Robert Asiedu, Jana Čížková, and et al. 2021. "Cytological and Molecular Characterization for Ploidy Determination in Yams (Dioscorea spp.)" Agronomy 11, no. 10: 1897. https://doi.org/10.3390/agronomy11101897
APA StyleGatarira, C., Sládeková, L., Němečková, A., Šimoníková, D., Paliwal, R., Asfaw, A., Abberton, M., Badara, G., Asiedu, R., Čížková, J., Hřibová, E., Doležel, J., DeKoeyer, D., Adetimirin, V., & Bhattacharjee, R. (2021). Cytological and Molecular Characterization for Ploidy Determination in Yams (Dioscorea spp.). Agronomy, 11(10), 1897. https://doi.org/10.3390/agronomy11101897