Sheep Grazing and Wildfire: Disturbance Effects on Dry Grassland Vegetation in the Western Italian Alps

 , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
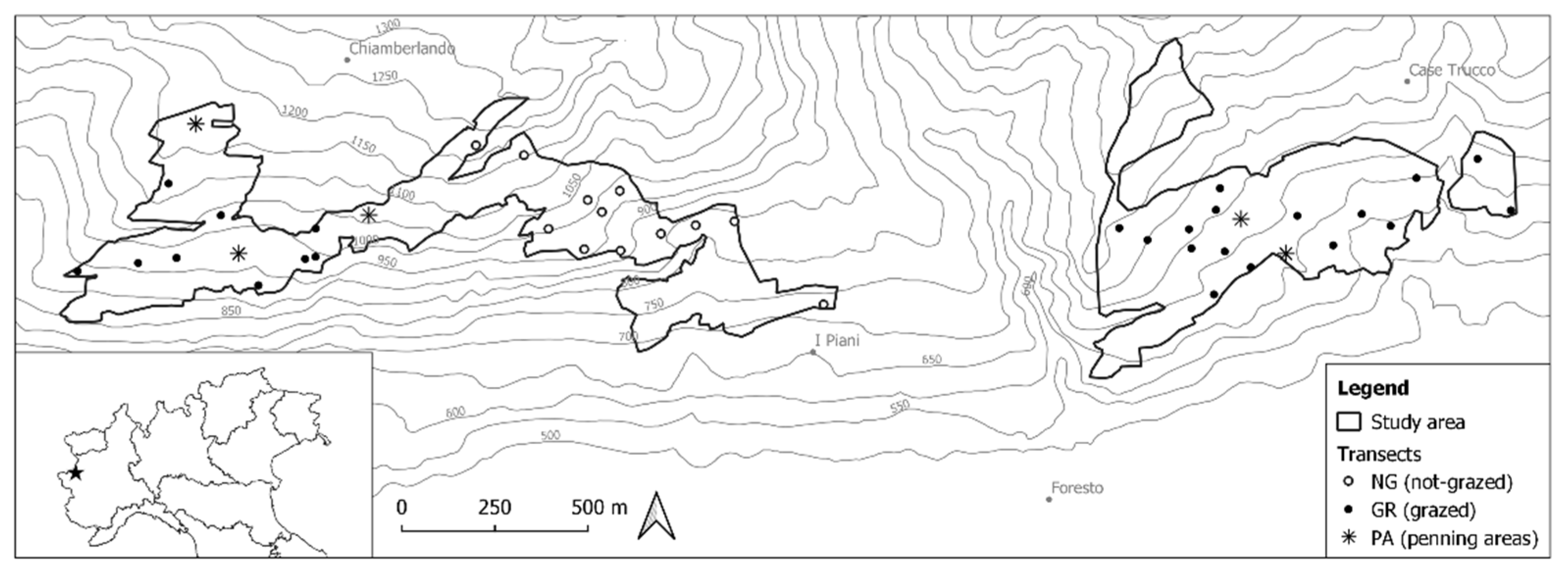
2.1. Study Area
2.2. Experimental Design
2.3. Vegetation Transects
2.4. Data Analysis
2.5. Statistical Analysis
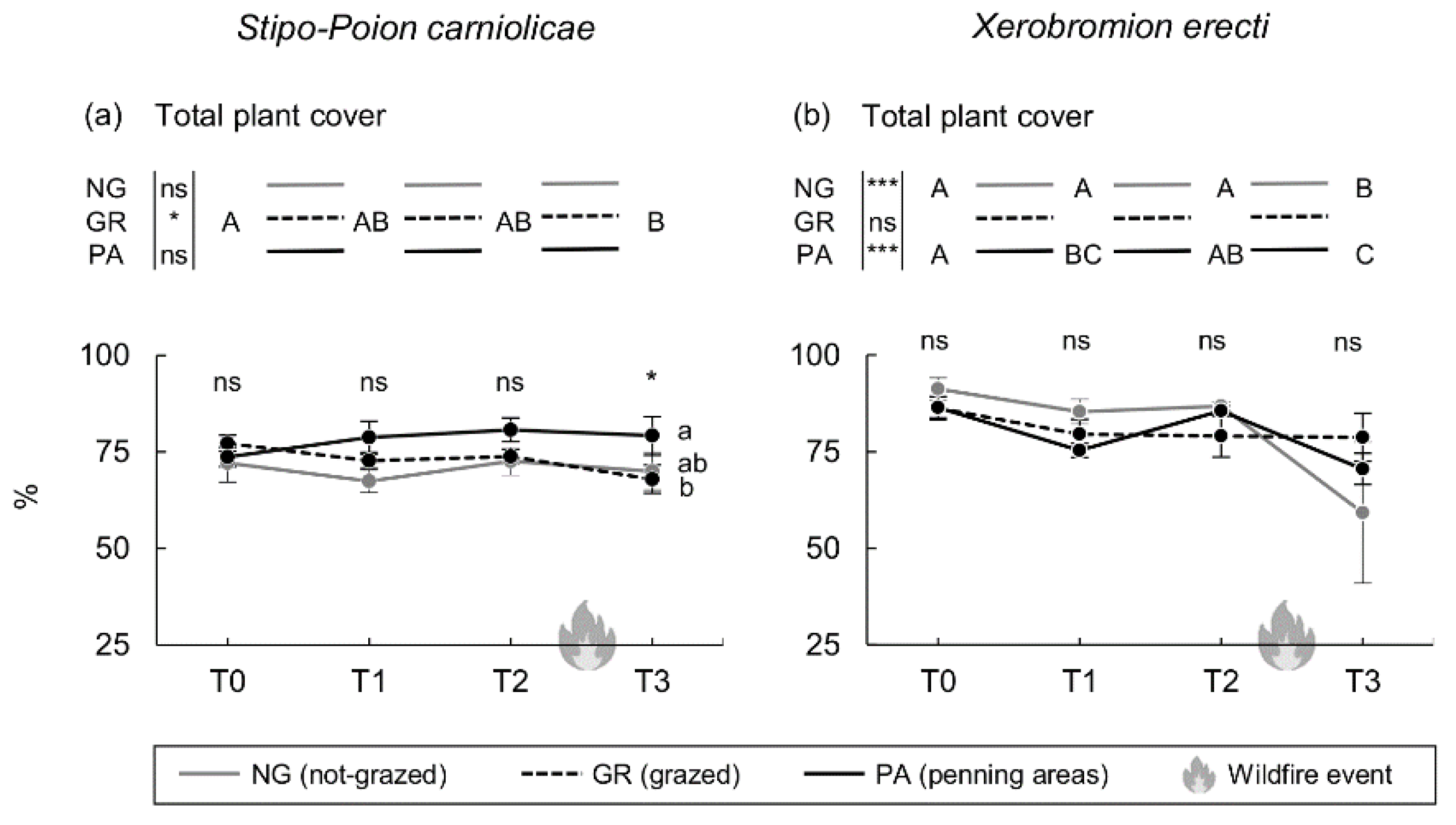
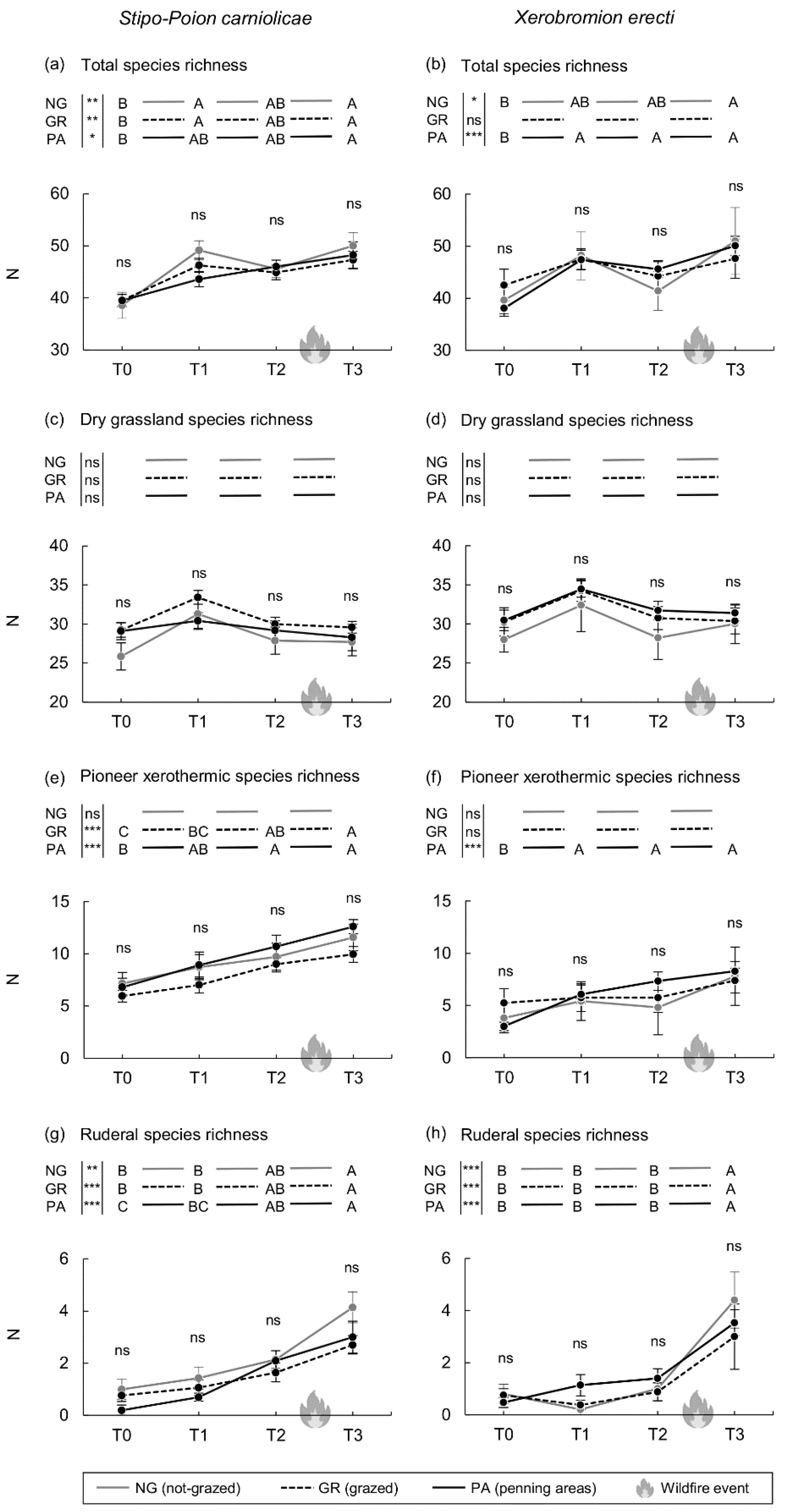
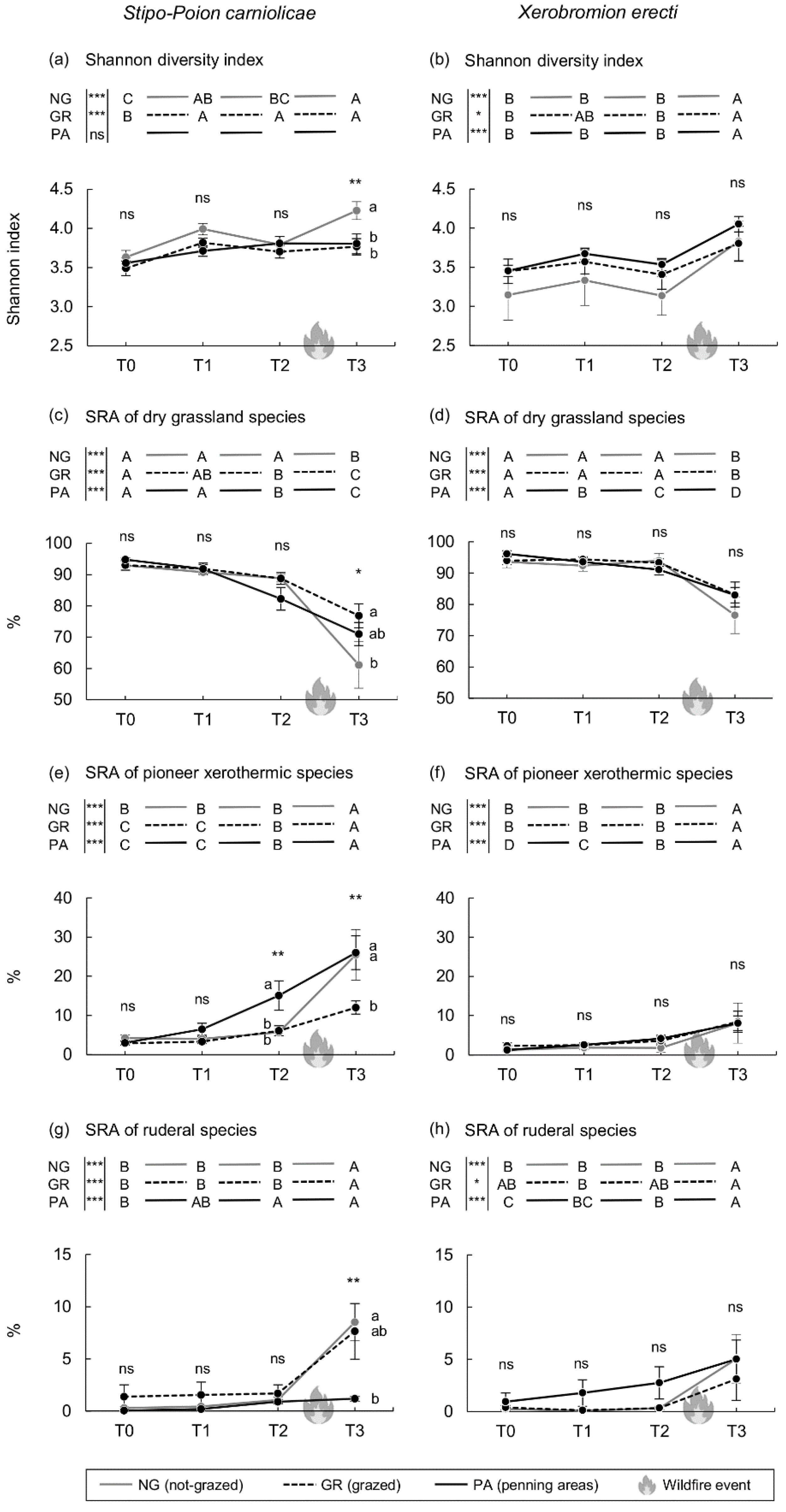
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poschlod, P.; Wallis De Vries, M.F. The historical and socioeconomic perspective of calcareous grasslands—lessons from the distant and recent past. Biol. Conserv. 2002, 104, 361–376. [Google Scholar] [CrossRef]
- Prévosto, B.; Kuiters, L.; Bernhardt-Römermann, M.; Dölle, M.; Schmidt, W.; Hoffmann, M.; Van Uytvanck, J.; Bohner, A.; Kreiner, D.; Stadler, J.; et al. Impacts of Land Abandonment on Vegetation: Successional Pathways in European Habitats. Folia Geobot. 2011, 46, 303–325. [Google Scholar] [CrossRef]
- Valkó, O.; Venn, S.; Żmihorski, M.; Biurrun, I.; Labadessa, R.; Loos, J. The challenge of abandonment for the sustainable management of Palaearctic natural and semi-natural grasslands. Hacquetia 2018, 17, 5–16. [Google Scholar] [CrossRef]
- Bohner, A.; Karrer, J.; Walcher, R.; Brandl, D.; Michel, K.; Arnberger, A.; Frank, T.; Zaller, J.G. Ecological responses of semi-natural grasslands to abandonment: Case studies in three mountain regions in the Eastern Alps. Folia Geobot. 2020, 54, 211–225. [Google Scholar] [CrossRef]
- Enyedi, Z.M.; Ruprecht, E.; Deák, M. Long-term effects of the abandonment of grazing on steppe-like grasslands. Appl. Veg. Sci. 2008, 11, 55–62. [Google Scholar] [CrossRef]
- Giarrizzo, E.; Burrascano, S.; Chiti, T.; de Bello, F.; Lepš, J.; Zavattero, L.; Blasi, C. Re-visiting historical semi-natural grasslands in the Apennines to assess patterns of changes in species composition and functional traits. Appl. Veg. Sci. 2017, 20, 247–258. [Google Scholar] [CrossRef]
- Hegedüšová, K.; Senko, D. Successional changes of dry grasslands in southwestern Slovakia after 46 years of abandonment. Plant Biosyst. 2011, 145, 666–687. [Google Scholar] [CrossRef]
- Pittarello, M.; Probo, M.; Lonati, M.; Lombardi, G. Restoration of sub-alpine shrub-encroached grasslands through pastoral practices: Effects on vegetation structure and botanical composition. Appl. Veg. Sci. 2016, 19, 381–390. [Google Scholar] [CrossRef]
- Orlandi, S.; Probo, M.; Sitzia, T.; Trentanovi, G.; Garbarino, M.; Lombardi, G.; Lonati, M. Environmental and land use determinants of grassland patch diversity in the western and eastern Alps under agro-pastoral abandonment. Biodivers. Conserv. 2016, 25, 275–293. [Google Scholar] [CrossRef]
- Görzen, E.; Borisova, K.; Fenesi, A.; Ruprecht, E.; Donath, T.W. Effects of woody species encroachment and fire on vegetation and the soil seed bank in dry grasslands of Transylvania. Appl. Veg. Sci. 2019, 22, 409–422. [Google Scholar] [CrossRef]
- Dengler, J.; Becker, T.; Ruprecht, E.; Szabó, A.; Becker, U.; Beldean, M.; Bita-Nicolae, C.; Dolnik, C.; Goia, I.; Peyrat, J.; et al. Festuco-Brometea communities of the Transylvanian Plateau (Romania)—A preliminary overview on syntaxonomy, ecology, and biodiversity. Tuexenia 2012, 32, 319–359. [Google Scholar]
- Wilson, J.B.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant species richness: The world records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Calaciura, B.; Spinelli, O. Management of Natura 2000 Habitats. 6210 Semi-Natural Dry Grasslands and Scrubland Facies on Calcareous Substrates (Festuco-Brometalia) (*Important Orchid Sites); European Commission: Brussels, Belgium, 2008. [Google Scholar]
- WallisDeVries, M.F.; Van Swaay, C.A.M. Grasslands as habitats for butterflies in Europe. In Grasslands in Europe of High Nature Value; Venn, P., Jefferson, R., de Smidt, J., van der Straaten, J., Eds.; KNNV Publishing: Utrecht, The Netherlands, 2009; pp. 27–34. [Google Scholar]
- Dostálek, J.; Frantík, T. Dry grassland plant diversity conservation using low-intensity sheep and goat grazing management: Case study in Prague (Czech Republic). Biodivers. Conserv. 2008, 17, 1439–1454. [Google Scholar] [CrossRef]
- Elias, D.; Hölzel, N.; Tischew, S. Positive effects of goat pasturing on the threatened spring geophyte Gagea bohemica in formerly abandoned dry grasslands. Flora 2018, 249, 53–59. [Google Scholar] [CrossRef]
- Elias, D.; Tischew, S. Goat pasturing—A biological solution to counteract shrub encroachment on abandoned dry grasslands in Central Europe? Agric. Ecosyst. Environ. 2016, 234, 98–106. [Google Scholar] [CrossRef]
- Schwabe, A.; Süss, K.; Storm, C. What are the long-term effects of livestock grazing in steppic sandy grassland with high conservation value? Results from a 12-year field study. Tuexenia 2013, 33, 189–212. [Google Scholar]
- Godó, L.; Valkó, O.; Tóthmérész, B.; Török, P.; Kelemen, A.; Deák, B. Scale-dependent effects of grazing on the species richness of alkaline and sand grasslands. Tuexenia 2017, 37, 229–246. [Google Scholar] [CrossRef]
- Pittarello, M.; Gorlier, A.; Lombardi, G.; Lonati, M. Plant species selection by sheep in semi-natural dry grasslands extensively grazed in the south-western Italian Alps. Rangel. J. 2017, 39, 123–131. [Google Scholar] [CrossRef]
- Rupprecht, D.; Gilhaus, K.; Hölzel, N. Effects of year-round grazing on the vegetation of nutrient-poor grass- and heathlands—Evidence from a large-scale survey. Agric. Ecosyst. Environ. 2016, 234, 16–22. [Google Scholar] [CrossRef]
- Ruprecht, E.; Enyedi, M.Z.; Eckstein, R.L.; Donath, T.W. Restorative removal of plant litter and vegetation 40 years after abandonment enhances re-emergence of steppe grassland vegetation. Biol. Conserv. 2010, 143, 449–456. [Google Scholar] [CrossRef]
- Ruprecht, E.; Enyedi, M.Z.; Szabó, A.; Fenesi, A. Biomass removal by clipping and raking vs burning for the restoration of abandoned Stipa-dominated European steppe-like grassland. Appl. Veg. Sci. 2016, 19, 78–88. [Google Scholar] [CrossRef]
- Schwabe, A.; Kratochwil, A. Festucetalia valesiacae communities and xerothermic vegetation complexes in the Central Alps related to environmental factors. Phytocoenologia 2004, 34, 329–446. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Die Inneralpine Trockenvegetation; Fischer: Stuttgart, Germany, 1961. [Google Scholar]
- Dengler, J.; Widmer, S.; Staubli, E.; Babbi, M.; Gehler, J.; Hepenstrick, D.; Bergamini, A.; Billeter, R.; Boch, S.; Rohrer, S.; et al. Dry grasslands of the central valleys of the Alps from a European perspective: The example of Ausserberg (Valais, Switzerland). Hacquetia 2019, 18, 155–177. [Google Scholar] [CrossRef]
- Royer, J.M. Synthèse eurosibérienne, phytosociologique et phytogéographique de la classe des Festuco-Brometea. Diss. Bot. 1991, 178, 1–296. [Google Scholar]
- Deák, B.; Valkó, O.; Török, P.; Végvári, Z.; Hartel, T.; Schmotzer, A.; Kapocsi, I.; Tóthmérész, B. Grassland fires in Hungary‒Experiences of nature conservationists on the effects of fire on biodiversity. Appl. Ecol. Environ. Res. 2014, 12, 267–283. [Google Scholar] [CrossRef]
- Stavi, I. Wildfires in Grasslands and Shrublands: A Review of Impacts on Vegetation, Soil, Hydrology, and Geomorphology. Water 2019, 11, 1042. [Google Scholar] [CrossRef]
- Valkó, O.; Török, P.; Deák, B.; Tóthmérész, B. Review: Prospects and limitations of prescribed burning as a management tool in European grasslands. Basic Appl. Ecol. 2014, 15, 26–33. [Google Scholar] [CrossRef]
- Biancotti, A.; Bellardone, G.; Bovo, S.; Cagnazzi, B.; Giacomelli, L.; Marchisio, C. Distribuzione Regionale di Piogge e Temperature; Regione Piemonte: Torino, Italy, 1998. [Google Scholar]
- Biondi, E.; Blasi, C.; Burrascano, S.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Galdenzi, D.; Gigante, D.; Lasen, C.; Spampinato, G.; et al. Manuale Italiano di Interpretazione Degli Habitat Della Direttiva 92/43/CEE; Società Botanica Italiana, Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Firenze, Italy, 2009. [Google Scholar]
- Conti, F.; Manzi, A.; Pedrotti, F. Liste Rosse Regionali Delle Piante d’Italia; WWF Italia, Società Botanica Italiana, Università degli Studi di Camerino: Camerino (MC), Firenze, Italy, 1997. [Google Scholar]
- Pignatti, S.; La Rosa, M.; Guarino, R. Flora D’italia, 2nd ed.; Edagricole di New Business Media: Bologna, Italy, 2017. [Google Scholar]
- Ravetto Enri, S.; Gorlier, A.; Nota, G.; Pittarello, M.; Lombardi, G.; Lonati, M. Distance from Night Penning Areas as an Effective Proxy to Estimate Site Use Intensity by Grazing Sheep in the Alps. Agronomy 2019, 9, 333. [Google Scholar] [CrossRef]
- Daget, P.; Poissonet, J. Une méthode d’analyse phytologique des prairies. Ann. Agron. 1971, 22, 5–41. [Google Scholar]
- Landolt, E.; Bäumler, B.; Erhardt, A.; Hegg, O.; Klötzli, F.; Lämmler, W.; Nobis, M.; Rudmann-Maurer, K.; Schweingruber, F.; Theurillat, J.-P. Flora Indicativa—Ecological Indicator Values and Biological Attributes of the Flora of Switzerland and the Alps; Haupt Verlag: Bern, Switzerland, 2010. [Google Scholar]
- Tasser, E.; Tappeiner, U. New model to predict rooting in diverse plant community compositions. Ecol. Model. 2005, 185, 195–211. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Aeschimann, D.; Lauber, K.; Moser, D.M.; Theurillat, J.-P. Flora Alpina; Zanichelli: Bologna, Italy, 2004. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Cribari-Neto, F.; Zeileis, A. Beta Regression in R. J. Stat. Softw. 2010, 34. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018.
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bolker, B.M. Glmm TMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Lenth, R. Emmeans: Estimated Marginal Means, aka Least-Squares Means, R Package Version 1.3.0; 2018. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 10 September 2019).
- Sternberg, M.; Gutman, M.; Perevolotsky, A.; Ungar, E.D.; Kigel, J. Vegetation response to grazing management in a Mediterranean herbaceous community: A functional group approach. J. Appl. Ecol. 2000, 37, 224–237. [Google Scholar] [CrossRef]
- Török, P.; Valkó, O.; Deák, B.; Kelemen, A.; Tóthmérész, B. Traditional Cattle Grazing in a Mosaic Alkali Landscape: Effects on Grassland Biodiversity along a Moisture Gradient. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Dostálek, J.; Frantík, T. Response of dry grassland vegetation to fluctuations in weather conditions: A 9-year case study in Prague (Czech Republic). Biologia 2011, 66, 837–847. [Google Scholar] [CrossRef][Green Version]
- Valkó, O.; Deák, B.; Magura, T.; Török, P.; Kelemen, A.; Tóth, K.; Horváth, R.; Nagy, D.D.; Debnár, Z.; Zsigrai, G.; et al. Supporting biodiversity by prescribed burning in grasslands—A multi-taxa approach. Sci. Total Environ. 2016, 572, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Vidaller, C.; Dutoit, T.; Ramone, H.; Bischoff, A. Fire increases the reproduction of the dominant grass Brachypodium retusum and Mediterranean steppe diversity in a combined burning and grazing experiment. Appl. Veg. Sci. 2019, 22, 127–137. [Google Scholar] [CrossRef]
- Lonati, M.; Vacchiano, G.; Berretti, R.; Motta, R. Effect of stand-replacing fires on Mediterranean plant species in their marginal alpine range. Alp. Bot. 2013, 123, 123–133. [Google Scholar] [CrossRef][Green Version]
- Ascoli, D.; Lonati, M.; Marzano, R.; Bovio, G.; Cavallero, A.; Lombardi, G. Prescribed burning and browsing to control tree encroachment in southern European heathlands. For. Ecol. Manag. 2013, 289, 69–77. [Google Scholar] [CrossRef]
- Silva, V.; Catry, F.X.; Fernandes, P.M.; Rego, F.C.; Bugalho, M.N. Trade-offs between fire hazard reduction and conservation in a Natura 2000 shrub–grassland mosaic. Appl. Veg. Sci. 2020, 23, 39–52. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stipo-Poion carniolicae | Xerobromion erecti | Total | |
|---|---|---|---|
| NG | 7 | 5 | 12 |
| GR | 17 | 8 | 25 |
| PA | 10 | 15 | 25 |
| Total | 34 | 28 | 62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nota, G.; Ravetto Enri, S.; Pittarello, M.; Gorlier, A.; Lombardi, G.; Lonati, M. Sheep Grazing and Wildfire: Disturbance Effects on Dry Grassland Vegetation in the Western Italian Alps. Agronomy 2021, 11, 6. https://doi.org/10.3390/agronomy11010006
Nota G, Ravetto Enri S, Pittarello M, Gorlier A, Lombardi G, Lonati M. Sheep Grazing and Wildfire: Disturbance Effects on Dry Grassland Vegetation in the Western Italian Alps. Agronomy. 2021; 11(1):6. https://doi.org/10.3390/agronomy11010006
Chicago/Turabian StyleNota, Ginevra, Simone Ravetto Enri, Marco Pittarello, Alessandra Gorlier, Giampiero Lombardi, and Michele Lonati. 2021. "Sheep Grazing and Wildfire: Disturbance Effects on Dry Grassland Vegetation in the Western Italian Alps" Agronomy 11, no. 1: 6. https://doi.org/10.3390/agronomy11010006
APA StyleNota, G., Ravetto Enri, S., Pittarello, M., Gorlier, A., Lombardi, G., & Lonati, M. (2021). Sheep Grazing and Wildfire: Disturbance Effects on Dry Grassland Vegetation in the Western Italian Alps. Agronomy, 11(1), 6. https://doi.org/10.3390/agronomy11010006