Optimisation of Weed Control in Organic Processing Spinach (Spinacia oleracea L.): Impacts of Cultivar, Seeding Rate, Plant Spacing and Integrated Weed Management Strategy



Abstract
1. Introduction
2. Materials and Methods
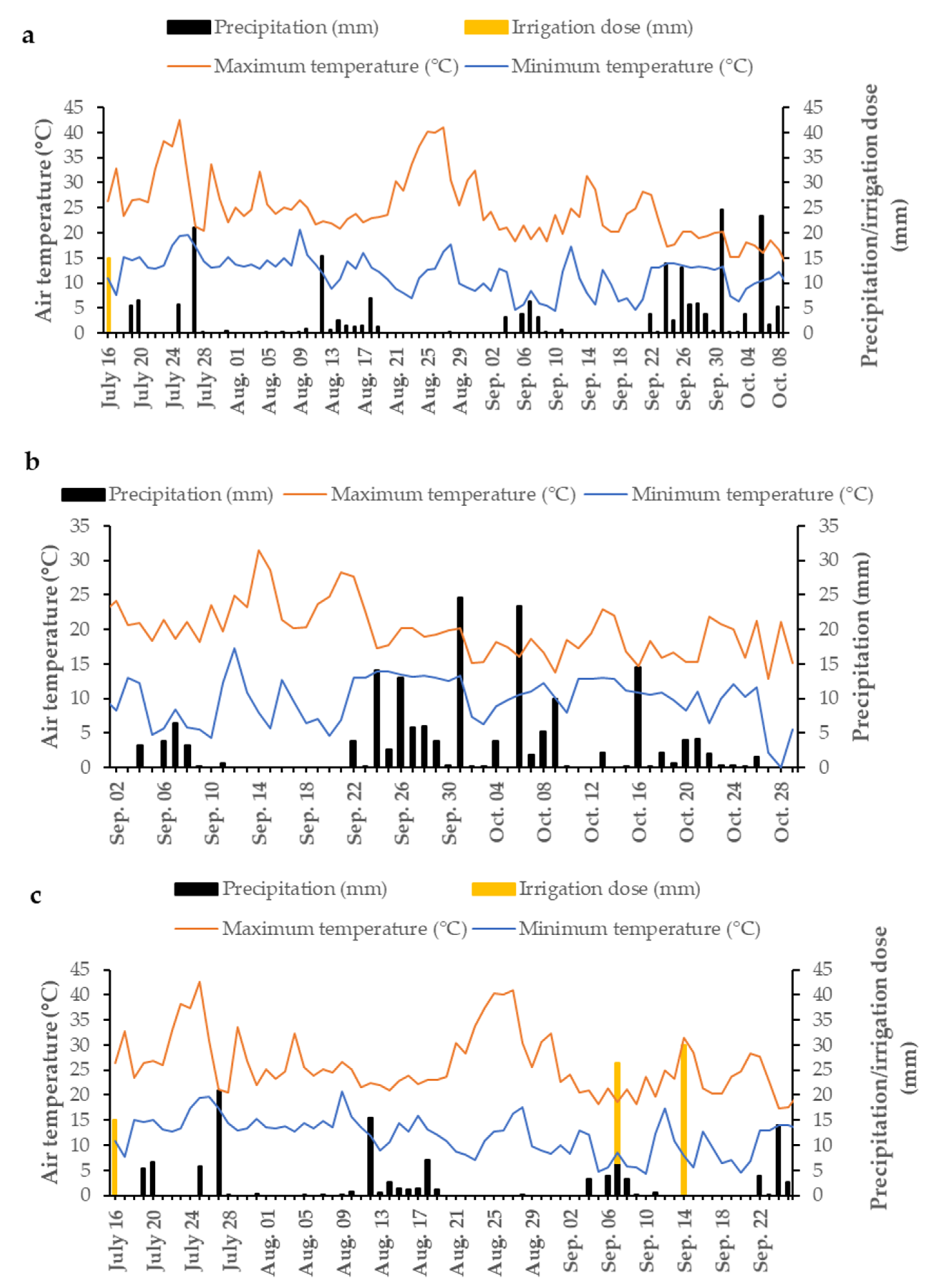
2.1. Field Experiments
2.2. Weeding Equipment
2.3. Measurements
2.4. Statistical Analysis
3. Results
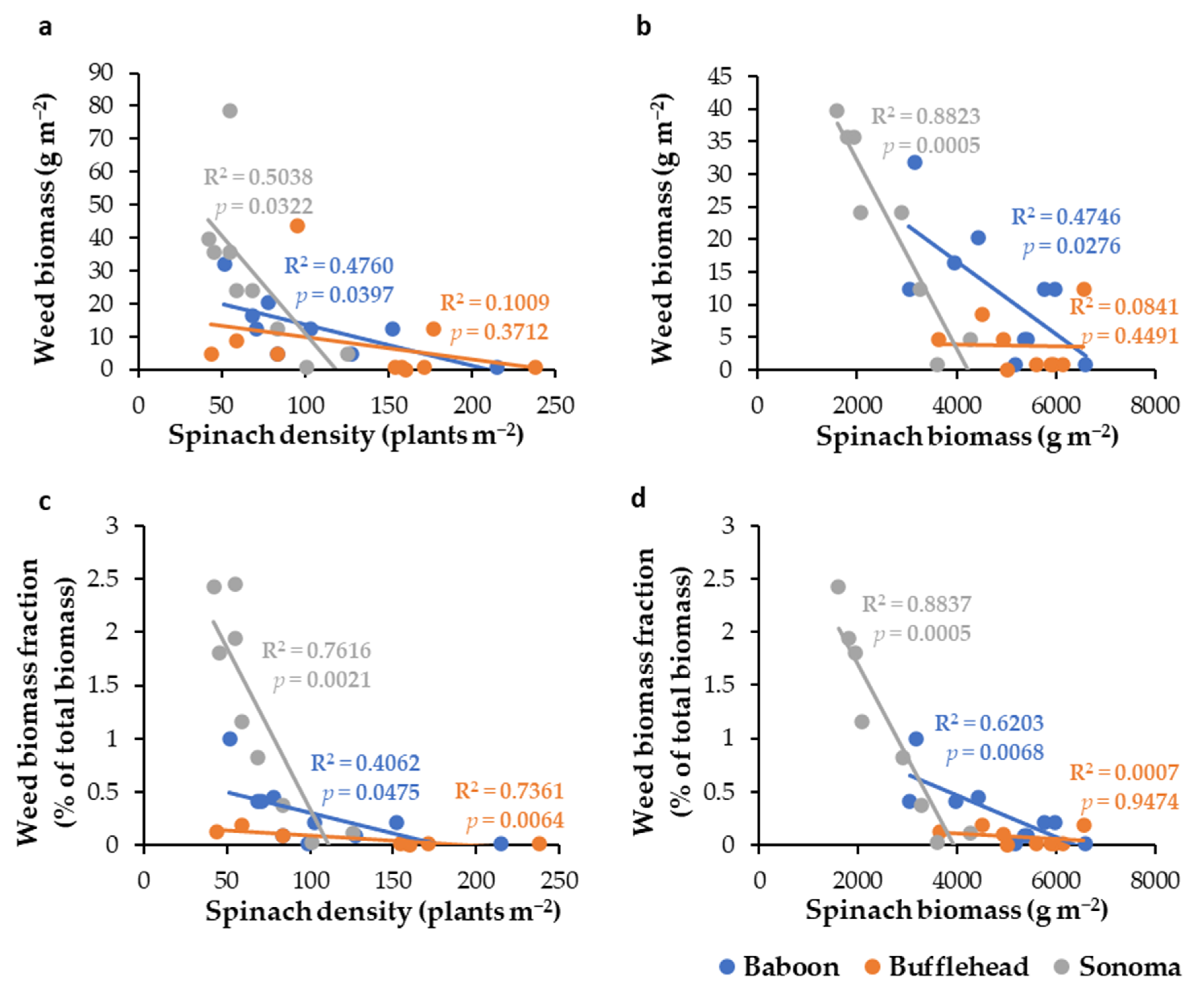
3.1. Impact of Spinach Cultivar and Seeding Rate (Experiment 1)
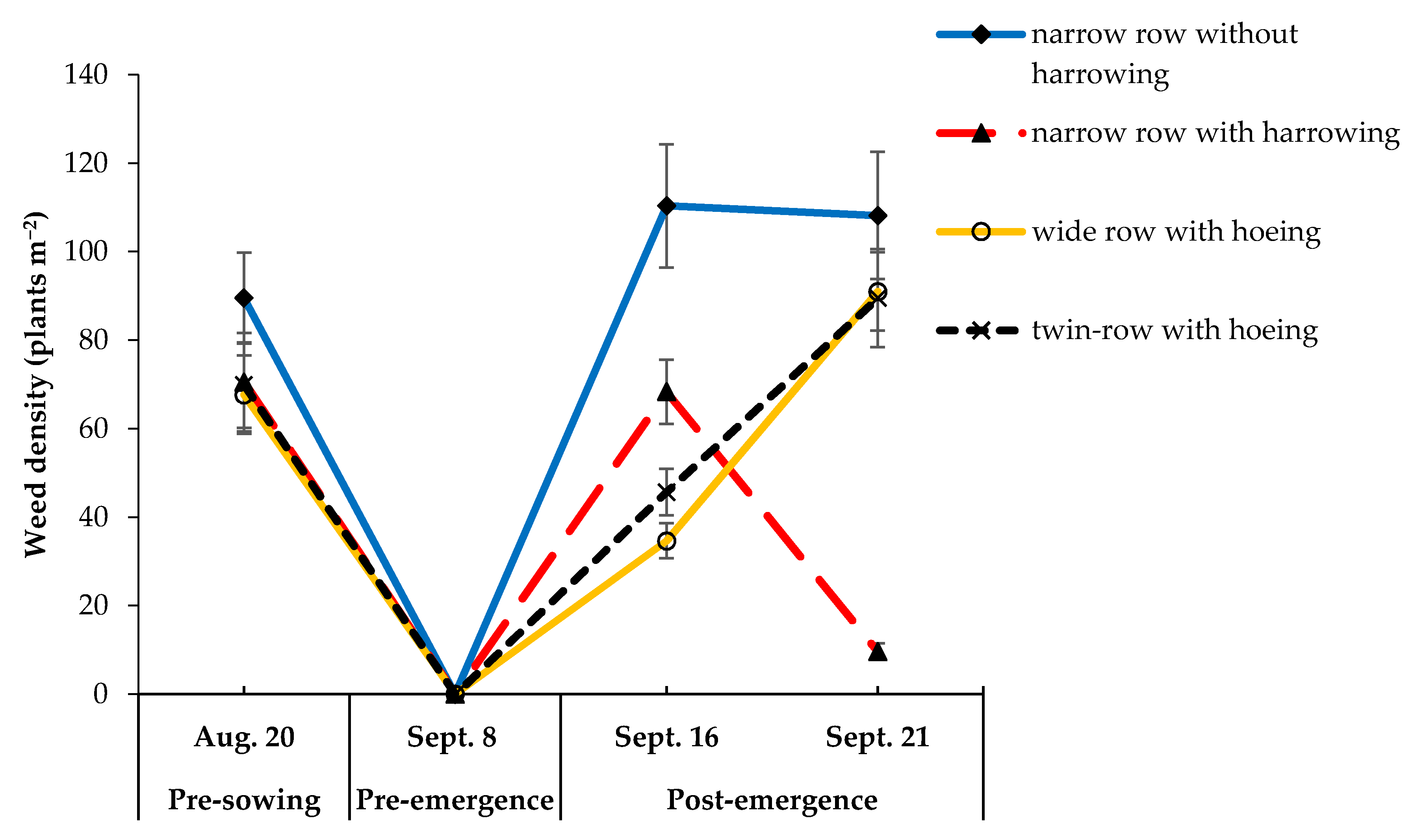
3.2. Impact of Plant Spacing Management Systems (Experiment 2)
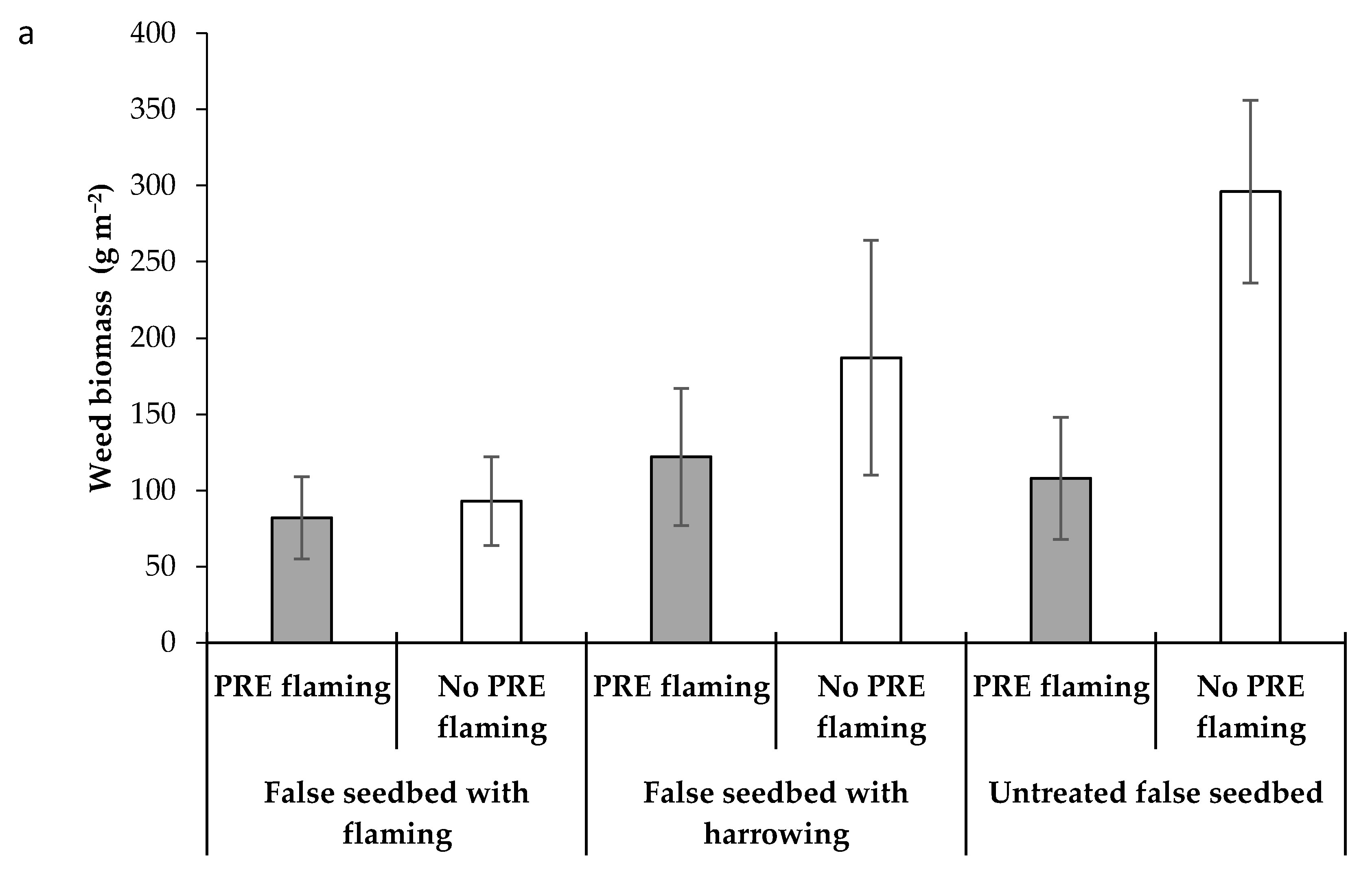
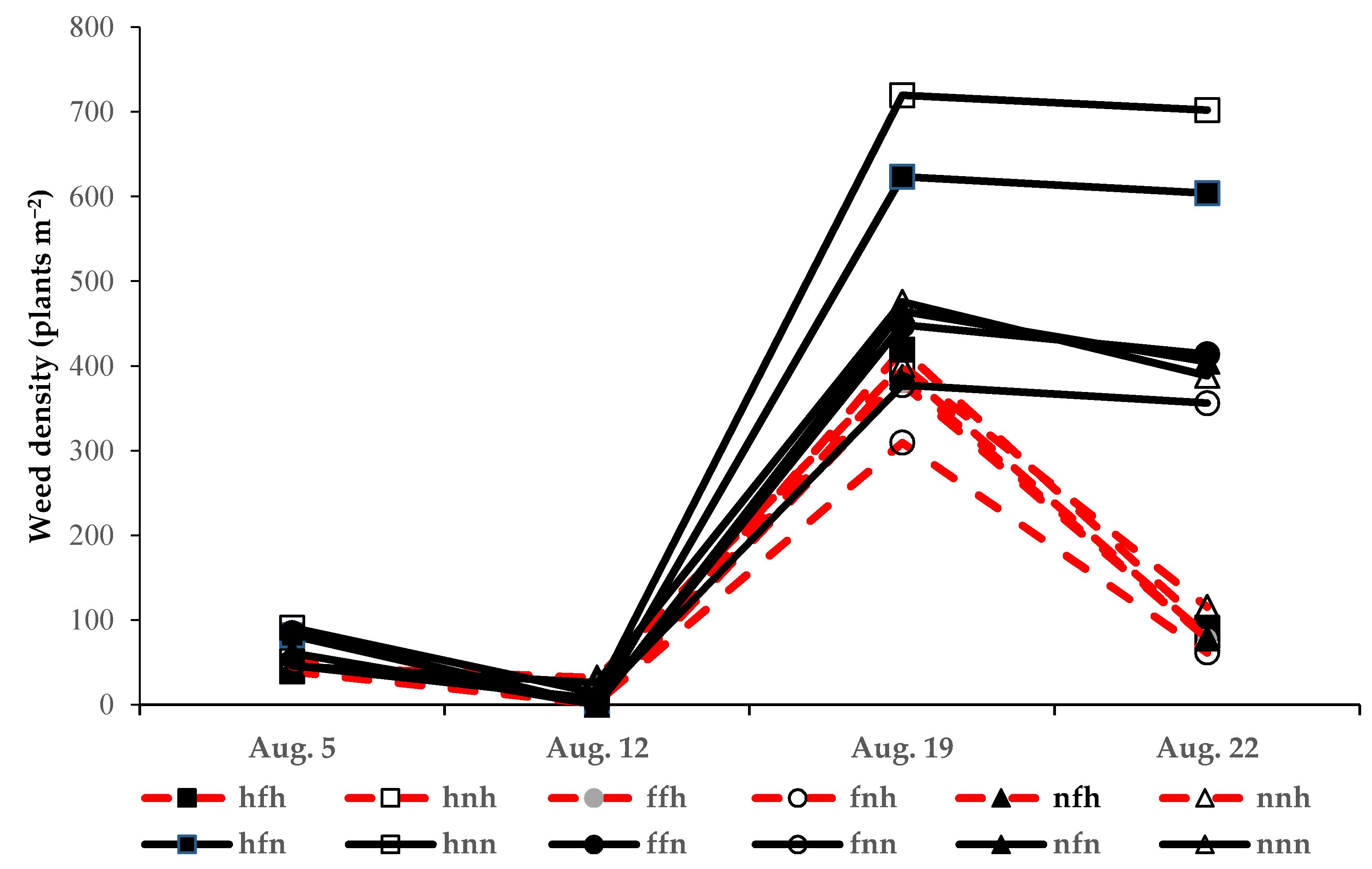
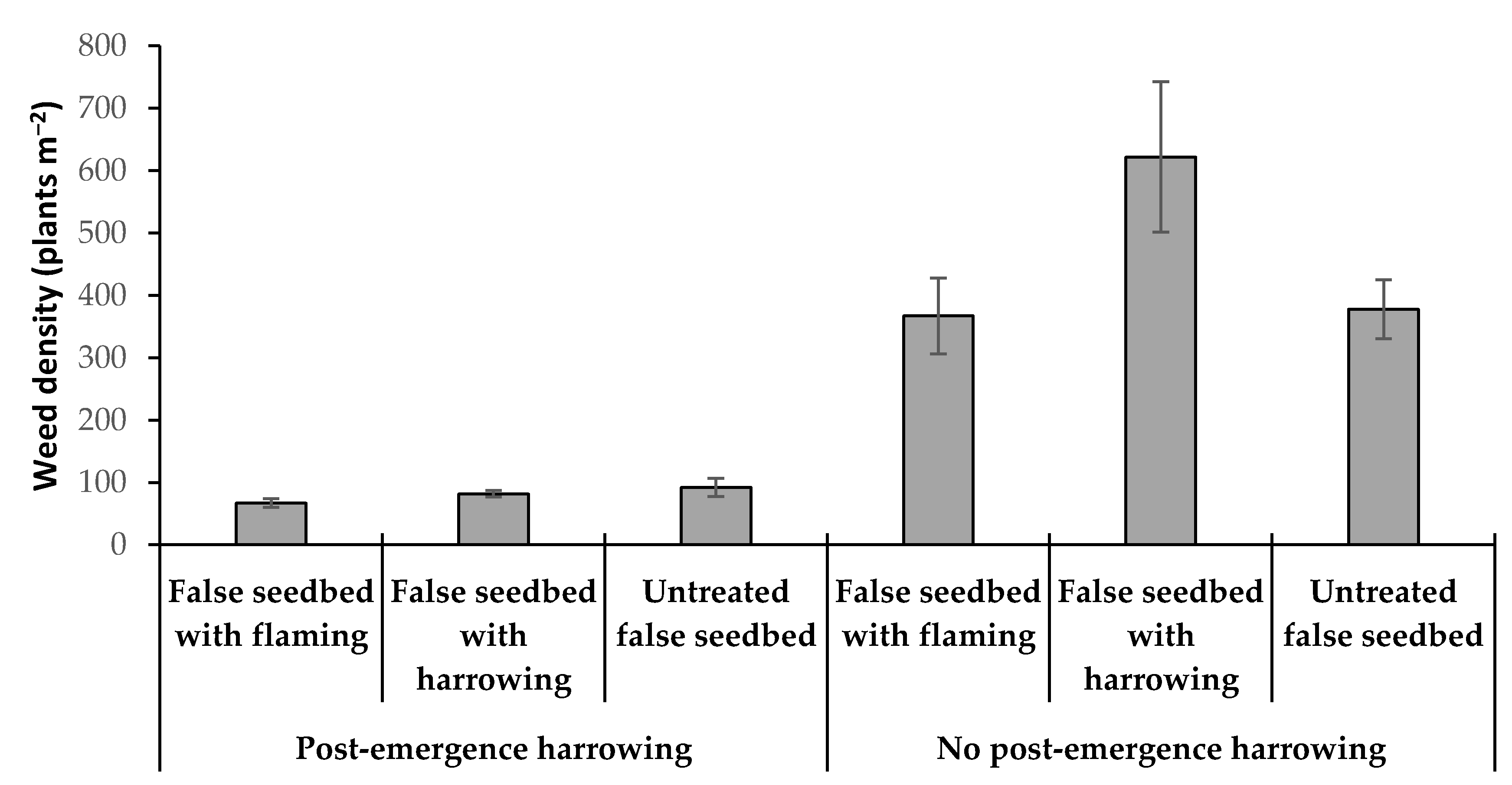
3.3. Impact of False Seedbed, Pre-Emergence and Post-Emergence Treatments (Experiment 3)
4. Discussion
4.1. Impact of Spinach Cultivar and Seeding Rate
4.2. Impact of Plant Spacing Management
4.3. Impact of False Seedbed, Pre-Emergence and Post-Emergence Tactics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Peruzzi, A.; Ginanni, M.; Raffaelli, M.; Borelli, M. Physical weed control in organic spinach production. In Proceedings of the 6th EWRS Symposium on Physical and Cultural Weed Control, Lillehamer, Norway, 8–10 March 2004; pp. 15–23. [Google Scholar]
- Spruijt, J.; van der Voort, M. Kwalitatieve Informatie Akkerbouw en Vollegrondsgroenteteelt, 29th ed.; Praktijkonderzoek Plant & Omgeving: Lelystad, The Nederlands, 2015. (In Dutch) [Google Scholar]
- Lichtenhahn, M.; Koller, M.; Dierauer, H.; Baumann, D. Biogemüsebau: Unkrautregulierung—Termingerecht und Schlagkräftig, 2nd ed.; Forschungsinstitut für Biologischen Landbau (FiBL): Frankfurt am Main, Germany, 2002; Available online: https://shop.fibl.org/chde/1027-gemuese-unkraut.html (accessed on 17 November 2020).
- Tei, F.; Stagnari, F.; Granier, A. Preliminary results on physical weed control in processing spinach. In Proceedings of the 5th EWRS Workshop on Physical and Cultural Weed Control, Pisa, Italy, 11–13 March 2002; pp. 164–171. [Google Scholar]
- Peruzzi, A.; Ginanni, M.; Raffaelli, M.; Fontanelli, M. Physical weed control in organic spinach in the Serchio Valley, Italy. In Proceedings of the 13th EWRS Symposium on Physical and Cultural Weed Control, Bari, Italy, 19–23 June 2005. [Google Scholar]
- Boyd, N.S.; Brennan, E.B.; Fennimore, S.A. Stale seedbed techniques for organic vegetable production. Weed Tech. 2006, 20, 1052–1057. [Google Scholar] [CrossRef]
- Fischer, R.A.; Miles, R.E. The role of spatial pattern in the competition between crop plants and weeds. A theoretical analysis. Math. Biosci. 1973, 18, 335–350. [Google Scholar] [CrossRef]
- Schnieders, B.J. A Quantitative Analysis of Inter-Specific Competition in Crops with a Row Structure. Ph.D. Thesis, Agricultural University Wageningen, Wageningen, The Netherlands, 1999. [Google Scholar]
- Bàrberi, P.; Bigongiali, F.; Antichi, D.; Carlesi, S.; Fontanelli, M.; Frasconi, C.; Lulli, L. Innovative crop and weed management strategies for organic spinach: Crop yield and weed suppression. In Cultivating the Future Based on Science, Proceedings of the 2nd Conference of the International Society of Organic Agriculture Research ISOFAR, Modena, Italy, 18–20 June 2008; Research Institute of Organic Agriculture FiBL: Frick, Switzerland, 2008; Available online: https://orgprints.org/12314/ (accessed on 17 November 2020).
- Fontanelli, M.; Martelloni, L.; Raffaelli, M.; Frasconi, C.; Ginanni, M.; Peruzzi, A. Weed management in autumn fresh market spinach: A nonchemical alternative. HortTechnology 2015, 25, 177–184. [Google Scholar] [CrossRef]
- Thompson, K.; Bakker, J.P.; Bekker, R.M. The Soil Seed Banks of North West Europe: Methodology, Density and Longevity; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- De Cauwer, B.; De Cuypere, T.; De Ryck, S.; Delanote, L.; Dewaele, K.; Willekens, K.; Reheul, D. Reduction in field emergence and seedbank density of Galinsoga quadriradiata and other weeds after contrasting false seedbed strategies in organic vegetable fields. Weed Res. 2019, 59, 265–278. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 7 March 2020).
- Gomez, K.; Gomez, A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1984; pp. 139–207. [Google Scholar]
- Drews, S.; Neufoff, D.; Köpke, U. Weed suppression ability of three winter wheat varieties at different row spacing under organic farming conditions. Weed Res. 2009, 49, 526–533. [Google Scholar] [CrossRef]
- Yoshida, Y.; Fujiwara, T.; Kumakura, H. Effects of plug transplanting on growth and bolting of spinach. Acta Hort. 2008, 771, 145–149. [Google Scholar] [CrossRef]
- McCollough, M.; Gallandt, E.; Darby, H.; Molloy, T. Band sowing with hoeing in organic grains: I. Comparisons with alternative weed management practices in spring barley. Weed Sci. 2019, 68, 285–293. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment 1 | Experiment 2 | Experiment 3 | |
|---|---|---|---|
| False seedbed period: | |||
| Seedbed preparation | July 18 | July 18 | |
| Harrowing pass | July 30 | August 7 * | |
| Flaming pass | August 12 | August 7 * | |
| Harrowing pass | August 20 | ||
| Pre-emergence period: | |||
| Sowing date | August 20 | September 3 (+ seedbed preparation) | August 7 |
| Spinach cultivar | Bufflehead, Baboon, Sonoma | Bufflehead | La Paz |
| Seeding rate (seeds m−2) | 300, 400 | 300 | 300 |
| Row spacing (cm) | 10.5 | 10.5 (narrow row), 21 (wide row), 10.5/21 (twin-row) | 10.5 |
| Weed control treatment | August 23 (harrowing) | September 8 (flaming) | August 12 (flaming) * |
| Post-emergence period: | |||
| Crop emergence | August 26 (Bufflehead), August 27 (Baboon), August 28 (Sonoma) | September 10 | August 13 |
| First harrowing pass | September 17 (10.5 cm inter-row zone) | August 20 * | |
| Hoeing pass | September 21 (21 cm inter-row zone) | ||
| Second harrowing pass | September 23 (10.5 cm inter-row zone) | August 23 * | |
| Harvest date | October 10 | October 29 | September 24–25 |
| ANOVA 1 | Spinach Cultivar 2 | ||||||
|---|---|---|---|---|---|---|---|
| Block | Seeding Rate | Cultivar | Seeding Rate × Cultivar | Baboon | Bufflehead | Sonoma | |
| Spinach parameters: | |||||||
| Spinach biomass (g m−2) | NS | NS | *** | NS | 4 893 ± 379.1 a | 5 445 ± 285.1 a | 2 751 ± 275.8 b |
| Marketable spinach biomass(g m−2) | NS | NS | *** | NS | 3 469 ± 302.3 a | 4 047 ± 235.6 a | 1 929 ± 201.7 b |
| Spinach waste (g m−2) | ** | NS | *** | NS | 1 424 ± 124.4 a | 1 398 ± 99.6 a | 0 823 ± 130.4 b |
| Spinach waste fraction(% of spinach biomass) | * | NS | NS | NS | 29.5 ± 2.25 a | 25.7 ± 1.48 a | 29.8 ± 4.11 a |
| Petiole fraction(% of marketable spinach) | NS | NS | *** | NS | 30.5 ± 1.54 a | 35.6 ± 1.52 a | 17.0 ± 0.98 b |
| Weed parameters: | |||||||
| Weed biomass (g m−2) | NS | NS | * | NS | 0 012 ± 3.1 b | 0 00 8 ± 4.2 b | 0 038 ± 11.6 a |
| Weed biomass fraction(% of total biomass) | NS | NS | *** | NS | 00.3 ± 0.09 b | 0 0.1 ± 0.06 b | 01.5 ± 0.40 a |
| ANOVA 1 | Plant Spacing Management Systems 2 | |||||
|---|---|---|---|---|---|---|
| Block | Plant Spacing Management | Narrow-Row without Harrowing | Narrow-Row with Harrowing | Wide-Row with Hoeing | Twin Row with Hoeing | |
| Spinach parameters: | ||||||
| Spinach biomass (g m−2) | NS | * | 2 902 ± 117 a | 2 368 ± 117 b | 2 807 ± 117 a | 2 623 ± 64 ab |
| Marketable spinach biomass (g m−2) | NS | ** | 1 609 ± 118 a | 1 225 ± 57 b | 1 705 ± 112 a | 1 595 ± 91 a |
| Spinach waste (g m−2) | NS | * | 1 293 ± 44 a | 1 143 ± 87 ab | 1 102 ± 34 b | 1 028 ± 58 b |
| Spinach waste fraction(% of spinach biomass) | NS | ** | 44.9 ± 2.35 a | 48.0 ± 2.02 a | 39.6 ± 1.93 b | 39.3 ± 2.44 b |
| Petiole fraction(% of marketable spinach biomass) | NS | ** c | 38.9 ± 1.17 a | 30.2 ± 1.56 c | 35.0 ± 0.85 b | 35.6 ± 0.97 ab |
| Weed parameters: | ||||||
| Weed biomass (g m−2) | * | *** c | 0 73 ± 11 a | 0 12 ± 2 c | 0 36 ± 5 b | 0 45 ± 5 b |
| Weed biomass fraction(% of total biomass) | * | *** c | 2.5 ± 0.39 a | 0.5 ± 0.09 c | 1.3 ± 0.23 b | 1.7 ± 0.20 b |
| BLOCK | POST | FS | PRE | POST × FS | POST × PRE | FS × PRE | POST × FS × PRE | |
|---|---|---|---|---|---|---|---|---|
| Weed parameters: | ||||||||
| Weed biomass (g m−2) | NS | ** | * | *** | NS | NS | ** | NS |
| Weed biomass fraction | NS | ** | NS | ** | NS | NS | * | NS |
| (% of total biomass) | ||||||||
| Weed density (plants m−2): | ||||||||
| August 5 | NS | NS | NS | NS | NS | NS | NS | NS |
| August 22 | NS | * | * | NS | * | NS | NS | NS |
| Spinach parameters: | ||||||||
| Spinach biomass (g m−2) | NS | NS | NS | NS | NS | NS | NS | NS |
| Spinach density (plants m−2): | ||||||||
| August 16 | NS | NS | NS | NS | NS | NS | NS | NS |
| August 26 | NS | * | ** | NS | NS | * | NS | * |
| hfh | hnh | ffh | fnh | nfh | nnh | hfn | hnn | ffn | fnn | nfn | nnn | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Weed biomass (g m−2): | ||||||||||||
| mean | 13.2 | 37.3 | 9.3 | 12.4 | 15.5 | 178.8 | 230.1 | 337.4 | 153.9 | 172.6 | 199.8 | 413.6 |
| SE | 2.31 | 19.12 | 0.69 | 3.81 | 3.37 | 63.03 | 50.67 | 109.4 | 24.67 | 21.84 | 47.21 | 57.31 |
| Weed biomass fraction (% of total biomass): | ||||||||||||
| mean | 0.5 | 1.4 | 0.3 | 0.4 | 0.6 | 6.7 | 7.7 | 12.2 | 5.1 | 5.8 | 7.3 | 13.6 |
| SE | 0.07 | 0.77 | 0.03 | 0.12 | 0.13 | 2.76 | 2.26 | 5.16 | 0.95 | 0.63 | 1.79 | 2.59 |
| Spinach biomass (g m−2): | ||||||||||||
| mean | 2 625 | 2 772 | 2 943 | 2 921 | 2 856 | 2 633 | 2 965 | 2 617 | 2 928 | 2 799 | 2 564 | 2 735 |
| SE | 338.7 | 249.0 | 231.8 | 378.3 | 262.0 | 258.8 | 234.5 | 286.9 | 188.2 | 306.1 | 148.9 | 251.9 |
| Spinach density (plants m−2): | ||||||||||||
| August 16; | ||||||||||||
| mean | 309 | 318 | 288 | 292 | 245 | 417 | 326 | 280 | 272 | 278 | 334 | 307 |
| SE | 36 | 33.2 | 31.5 | 9.4 | 7.4 | 44.6 | 40.7 | 20.2 | 26.2 | 29.8 | 30.5 | 7.5 |
| August 26; | ||||||||||||
| mean | 197 | 213 | 221 | 202 | 165 | 300 | 285 | 287 | 262 | 229 | 295 | 247 |
| SE | 22.2 | 25.8 | 23.3 | 13.9 | 5.3 | 27.4 | 25.4 | 17.6 | 25.5 | 4.0 | 23.0 | 7.9 |
| Significance † | ** | ** | * | ** | ** | NS | NS | NS | NS | NS | * | ** |
| Cohen’s d effect size for a paired t-test | −1.5 | −1.41 | −0.96 | −3.03 | −4.99 | −1.26 | −0.49 | 0.15 | −0.15 | −0.92 | −0.58 | −3.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Cauwer, B.; Delanote, L.; Devos, M.; De Ryck, S.; Reheul, D. Optimisation of Weed Control in Organic Processing Spinach (Spinacia oleracea L.): Impacts of Cultivar, Seeding Rate, Plant Spacing and Integrated Weed Management Strategy. Agronomy 2021, 11, 53. https://doi.org/10.3390/agronomy11010053
De Cauwer B, Delanote L, Devos M, De Ryck S, Reheul D. Optimisation of Weed Control in Organic Processing Spinach (Spinacia oleracea L.): Impacts of Cultivar, Seeding Rate, Plant Spacing and Integrated Weed Management Strategy. Agronomy. 2021; 11(1):53. https://doi.org/10.3390/agronomy11010053
Chicago/Turabian StyleDe Cauwer, Benny, Lieven Delanote, Michaël Devos, Sander De Ryck, and Dirk Reheul. 2021. "Optimisation of Weed Control in Organic Processing Spinach (Spinacia oleracea L.): Impacts of Cultivar, Seeding Rate, Plant Spacing and Integrated Weed Management Strategy" Agronomy 11, no. 1: 53. https://doi.org/10.3390/agronomy11010053
APA StyleDe Cauwer, B., Delanote, L., Devos, M., De Ryck, S., & Reheul, D. (2021). Optimisation of Weed Control in Organic Processing Spinach (Spinacia oleracea L.): Impacts of Cultivar, Seeding Rate, Plant Spacing and Integrated Weed Management Strategy. Agronomy, 11(1), 53. https://doi.org/10.3390/agronomy11010053