Developing Heterotic Groups for Successful Hybrid Breeding in Perennial Ryegrass

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Genotyping-by-Sequencing
2.3. Principal Component Analysis, Hierarchical Clustering, and Permutations
2.4. Yield Trials
3. Results
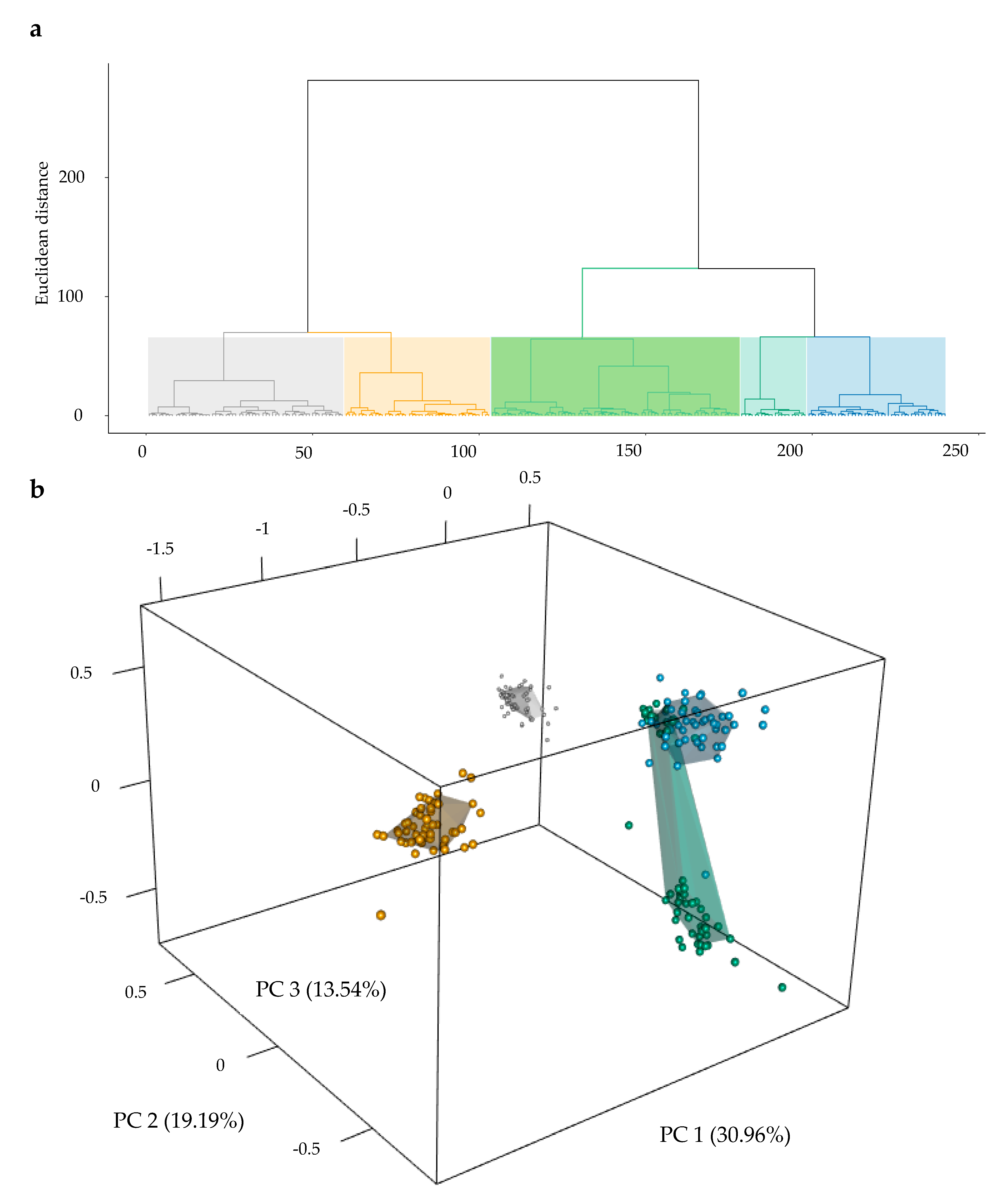
3.1. Genetic Structure of Selected Parental Populations from a CMS-Based Hybrid Breeding Program
3.2. Sample Numbers Required to Represent the Allelic Composition of Breeding Populations
3.3. Performance of Tetraploid Perennial Ryegrass Hybrids and Synthetic Varieties
4. Discussion
4.1. Improved Yield Performance through Hybrid Breeding
4.2. Identification of Heterotic Groups is Key for Successful and Efficient Hybrid Breeding in Perennial Ryegrass
4.3. Evidence for Sub-Populations within Heterotic Groups as Revealed by Genotyping-by-Sequencing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilkins, P.W.; Humphreys, M.O. Progress in breeding perennial forage grasses for temperate agriculture. J. Agric. Sci. 2003, 140, 129–150. [Google Scholar] [CrossRef]
- Bothe, A.; Westermeier, P.; Wosnitza, A.; Willner, E.; Schum, A.; Dehmer, K.J.; Hartmann, S. Drought tolerance in perennial ryegrass (Lolium perenne L.) as assessed by two contrasting phenotyping systems. J. Agron. Crop. Sci 2018, 204, 375–389. [Google Scholar] [CrossRef]
- Loka, D.; Harper, J.; Humphreys, M.; Gasior, D.; Wootton-Beard, P.; Gwynn-Jones, D.; Scullion, J.; Doonan, J.; Kingston-Smith, A.; Dodd, R.; et al. Impacts of abiotic stresses on the physiology and metabolism of cool-season grasses: A review. Food Energy Secur. 2019, 8, e00152. [Google Scholar] [CrossRef]
- Cyriac, D.; Hofmann, R.W.; Stewart, A.; Sathish, P.; Winefield, C.S.; Moot, D.J. Intraspecific differences in long-term drought tolerance in perennial ryegrass. PLoS ONE 2018, 13, e0194977. [Google Scholar] [CrossRef] [PubMed]
- Do Canto, J.; Studer, B.; Lubberstedt, T. Overcoming self-incompatibility in grasses: A pathway to hybrid breeding. Theor. Appl. Genet. 2016, 129, 1815–1829. [Google Scholar] [CrossRef]
- Hayward, M.D.; Abdullah, I.B. Selection and stability of synthetic varieties of Lolium perenne. Theor. Appl. Genet. 1985, 70, 48–51. [Google Scholar] [CrossRef]
- Casler, M.D.; Brummer, E.C. Theoretical Expected Genetic Gains for Among-and-Within-Family Selection Methods in Perennial Forage Crops. Crop. Sci. 2008, 48, 890–902. [Google Scholar] [CrossRef]
- Aguirre, A.A.; Studer, B.; Frei, U.; Lubberstedt, T. Prospects for Hybrid breeding in Bioenergy Grasses. Bioenergy Res. 2012, 5, 10–19. [Google Scholar] [CrossRef]
- Carew, R.; Florkowski, W.J.; Meng, T. Intellectual property rights and plant variety protection of horticultural crops: Evidence from Canada. Can. J. Plant. Sci. 2017, 97, 737–754. [Google Scholar] [CrossRef]
- Halpert, M.-T.; Chappell, M.J. Prima facie reasons to question enclosed intellectual property regimes and favor open-source regimes for germplasm. F1000Research 2017, 6, 284. [Google Scholar] [CrossRef]
- Blackmore, T.; Thorogood, D.; Skøt, L.; McMahon, R.; Powell, W.; Hegarty, M. Germplasm dynamics: The role of ecotypic diversity in shaping the patterns of genetic variation in Lolium perenne. Sci. Rep. 2016, 6, 22603. [Google Scholar] [PubMed]
- Pembleton, L.W.; Shinozuka, H.; Wang, J.; Spangenberg, G.C.; Forster, J.W.; Cogan, N.O.I. Design of an F1 hybrid breeding strategy for ryegrasses based on selection of self-incompatibility locus-specific alleles. Front. Plant Sci. 2015, 6, 764. [Google Scholar] [PubMed]
- Chen, E.; Huang, X.; Tian, Z.; Wing, R.A.; Han, B. The Genomics of Oryza Species Provides Insights into Rice Domestication and Heterosis. Annu. Rev. Plant Biol. 2019, 70, 639–665. [Google Scholar]
- Kempe, K.; Gils, M. Pollination control technologies for hybrid breeding. Mol. Breed. 2011, 27, 417–437. [Google Scholar]
- Islam, M.S.; Studer, B.; Møller, I.M.; Asp, T. Genetics and biology of cytoplasmic male sterility and its applications in forage and turf grass breeding. Plant Breed. 2014, 133, 299–312. [Google Scholar]
- Bolaric, S.; Barth, S.; Melchinger, A.E.; Posselt, U.K. Molecular characterization of genetic diversity in European germplasm of perennial ryegrass. Euphytica 2005, 146, 39–44. [Google Scholar]
- Zhao, Y.; Li, Z.; Liu, G.; Jiang, Y.; Maurer, H.P.; Würschum, T.; Mock, H.-P.; Matros, A.; Ebmeyer, E.; Schachschneider, R.; et al. Genome-based establishment of a high-yielding heterotic pattern for hybrid wheat breeding. Proc. Natl. Acad. Sci. USA 2015, 112, 15624–15629. [Google Scholar]
- Posselt, U.K. Hybrid production in Lolium perenne based on incompatibility. Euphytica 1993, 71, 29–33. [Google Scholar]
- Duvick, D.N. Biotechnology in the 1930s: The development of hybrid maize. Nat. Rev. Genet. 2001, 2, 69–74. [Google Scholar]
- Becker, H.C.; Löptien, H.; Röbbelen, G. Breeding: An overview. In Developments in Plant Genetics and Breeding; Gómez-Campo, C., Ed.; Elsevier: Amsterdam, The Netherlands, 1999; Volume 4, pp. 413–460. [Google Scholar]
- Miedaner, T.; Korzun, V.; Bauer, E. Chapter 15—Genomics-Based Hybrid Rye Breeding. In Applications of Genetic and Genomic Research in Cereals; Miedaner, T., Korzun, V., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 329–348. ISBN 9780081021637. [Google Scholar]
- Schlegel, R. Hybrid breeding boosted molecular genetics in rye. Russ. J. Genet. Appl. Res. 2016, 6, 569–583. [Google Scholar]
- Guan, X.; Yuyama, N.; Stewart, A.; Ding, C.; Xu, N.; Kiyoshi, T.; Cai, H. Genetic Diversity and Structure of Lolium Species Surveyed on Nuclear Simple Sequence Repeat and Cytoplasmic Markers. Front. Plant Sci. 2017, 8, 584. [Google Scholar] [CrossRef] [PubMed]
- Miedaner, T.; Glass, C.; Dreyer, F.; Wortmann, P.W.H.; Geiger, H.H. Mapping of genes for male-fertility restoration in ‘Pampa’ CMS winter rye (Secale cereale L.). Theor. Appl. Genet. 2000, 101, 1226–1233. [Google Scholar] [CrossRef]
- Laidig, F.; Piepho, H.-P.; Rentel, D.; Drobek, T.; Meyer, U.; Huesken, A. Breeding progress, variation, and correlation of grain and quality traits in winter rye hybrid and population varieties and national on-farm progress in Germany over 26 years. Theor. Appl. Genet. 2017, 130, 981–998. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Feuerstein, U.; Luesink, W.; Schulze, S.; Asp, T.; Studer, B.; Becker, H.C.; Dehmer, K.J. DArT, SNP, and SSR analyses of genetic diversity in Lolium perenne L. using bulk sampling. BMC Genet. 2018, 19, 10. [Google Scholar] [CrossRef]
- Byrne, S.L.; Nagy, I.; Pfeifer, M.; Armstead, I.; Swain, S.; Studer, B.; Mayer, K.; Campbell, J.D.; Czaban, A.; Hentrup, S.; et al. A synteny-based draft genome sequence of the forage grass Lolium perenne. Plant J. 2015, 84, 816–826. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef]
- Knaus, B.J.; Grünwald, N.J. vcfr: A package to manipulate and visualize variant call format data in R. Mol. Ecol. Resour. 2017, 17, 44–53. [Google Scholar] [CrossRef]
- Humphreys, M.O. The contribution of conventional plant breeding to forage crop improvement. In Proceedings of the 18th International Grassland Congress, Winnipeg and Saskatoon, SK, Canada, 8–17 June 1997. [Google Scholar]
- Conaghan, P.; Casler, M.D. A theoretical and practical analysis of the optimum breeding system for perennial ryegrass. Ir. J. Agric. Food Res. 2011, 50, 47–63. [Google Scholar]
- Posselt, U.K. Identification of Heterotic Patterns in Perennial Ryegrass. In Sustainable Use of Genetic Diversity in Forage and Turf Breeding; Springer: Dordrecht, The Netherlands, 2010; pp. 569–572. [Google Scholar]
- Yingheng, W.; Qiuhua, C.; Hongguang, X.; Fangxi, W.; Ling, L.; Wei, H.; Liping, C.; Hua’an, X.; Jianfu, Z. Determination of Heterotic Groups and Heterosis Analysis of Yield Performance in indica Rice. Rice Sci. 2018, 25, 261–269. [Google Scholar] [CrossRef]
- Dey, S.S.; Sharma, S.R.; Bhatia, R.; Parkash, C.; Barwal, R.N. Superior CMS (Ogura) lines with better combining ability improve yield and maturity in cauliflower (Brassica oleracea var. botrytis). Euphytica 2011, 182, 187. [Google Scholar] [CrossRef]
- Byrne, S.; Czaban, A.; Studer, B.; Panitz, F.; Bendixen, C.; Asp, T. Genome wide allele frequency fingerprints (GWAFFs) of populations via genotyping by sequencing. PLoS ONE 2013, 8, e57438. [Google Scholar] [CrossRef]
- Hale, M.L.; Burg, T.M.; Steeves, T.E. Sampling for microsatellite-based population genetic studies: 25 to 30 individuals per population is enough to accurately estimate allele frequencies. PLoS ONE 2012, 7, e45170. [Google Scholar] [CrossRef]
- Pembleton, L.W.; Drayton, M.C.; Bain, M.; Baillie, R.C.; Inch, C.; Spangenberg, G.C.; Wang, J.; Forster, J.W.; Cogan, N.O.I. Targeted genotyping-by-sequencing permits cost-effective identification and discrimination of pasture grass species and cultivars. Theor. Appl. Genet. 2016, 129, 991–1005. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vogt, M.; Yates, S.; Sykes, T.; Luesink, W.; Koch, M.; Studer, B. Developing Heterotic Groups for Successful Hybrid Breeding in Perennial Ryegrass. Agronomy 2020, 10, 1410. https://doi.org/10.3390/agronomy10091410
Vogt M, Yates S, Sykes T, Luesink W, Koch M, Studer B. Developing Heterotic Groups for Successful Hybrid Breeding in Perennial Ryegrass. Agronomy. 2020; 10(9):1410. https://doi.org/10.3390/agronomy10091410
Chicago/Turabian StyleVogt, Maximilian, Steven Yates, Timothy Sykes, Wilbert Luesink, Michael Koch, and Bruno Studer. 2020. "Developing Heterotic Groups for Successful Hybrid Breeding in Perennial Ryegrass" Agronomy 10, no. 9: 1410. https://doi.org/10.3390/agronomy10091410
APA StyleVogt, M., Yates, S., Sykes, T., Luesink, W., Koch, M., & Studer, B. (2020). Developing Heterotic Groups for Successful Hybrid Breeding in Perennial Ryegrass. Agronomy, 10(9), 1410. https://doi.org/10.3390/agronomy10091410