α-Tocopherol Foliar Spray and Translocation Mediates Growth, Photosynthetic Pigments, Nutrient Uptake, and Oxidative Defense in Maize (Zea mays L.) under Drought Stress

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Analysis
2.2. Estimation of Different Growth Parameters
2.3. Estimation of Leaf Photosynthetic Pigments
2.4. Leaf Relative Water Content (LRWC)
2.5. Leaf Relative Membrane Permeability
2.6. Estimation of Leaf Malondialdehyde Content
2.7. Extraction of Antioxidant Enzymes and Total Soluble Proteins from Different Plant Parts
2.7.1. Estimation of Total Soluble Proteins in Different Plant Parts
2.7.2. Estimation of the Activities of Superoxide Dismutase, Peroxidase, and Catalase in Different Plant Parts
2.8. Determination of Non-Enzymatic Antioxidants in Different Plant Parts
2.9. Determination Mineral Nutrients
2.9.1. Estimation of K+, Ca2+, and Mg2+ in Different Plan Parts
2.9.2. Determination of N and P
2.10. Statistical Analysis
3. Results
3.1. Different Growth Attributes and Content of Leaf Photosynthetic Pigments of Water-Stressed Maize Plants Foliar-Applied Alpha Tocopherol
3.2. Leaf Relative Water Content, Leaf Relative Membrane Permeability, Total Soluble Proteins, and H2O2 Contents of Leaf Photosynthetic Pigments of Maize Plants Foliar-Applied with Alpha Tocopherol
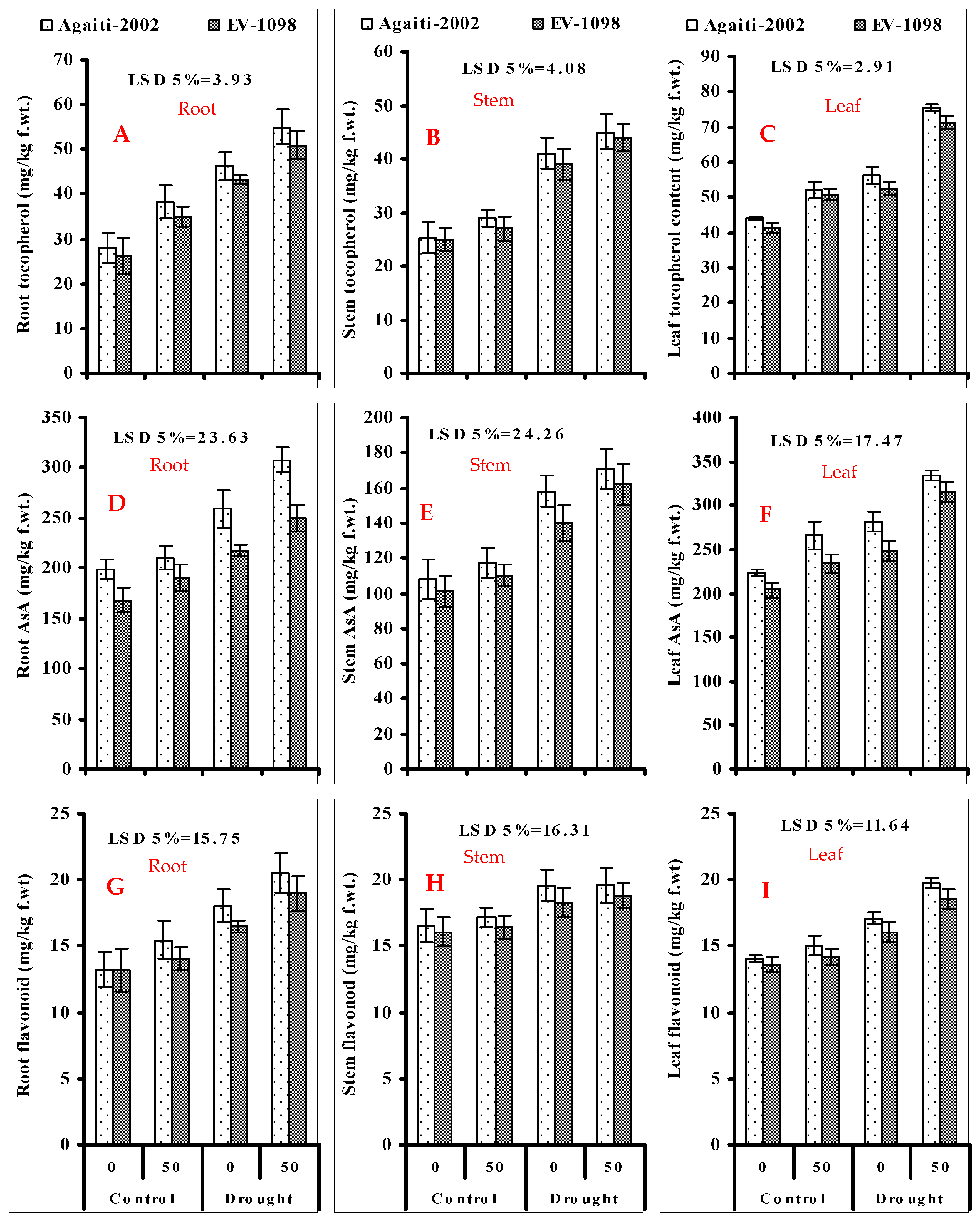
3.3. Root, Stem, and Leaf Total Tocopherol (Figure 1A–C); Ascorbic Acid (Figure 1D–F); and Total Flavonoid Contents (Figure 1G–I) of Maize Plants Foliar-Applied with α-Toc
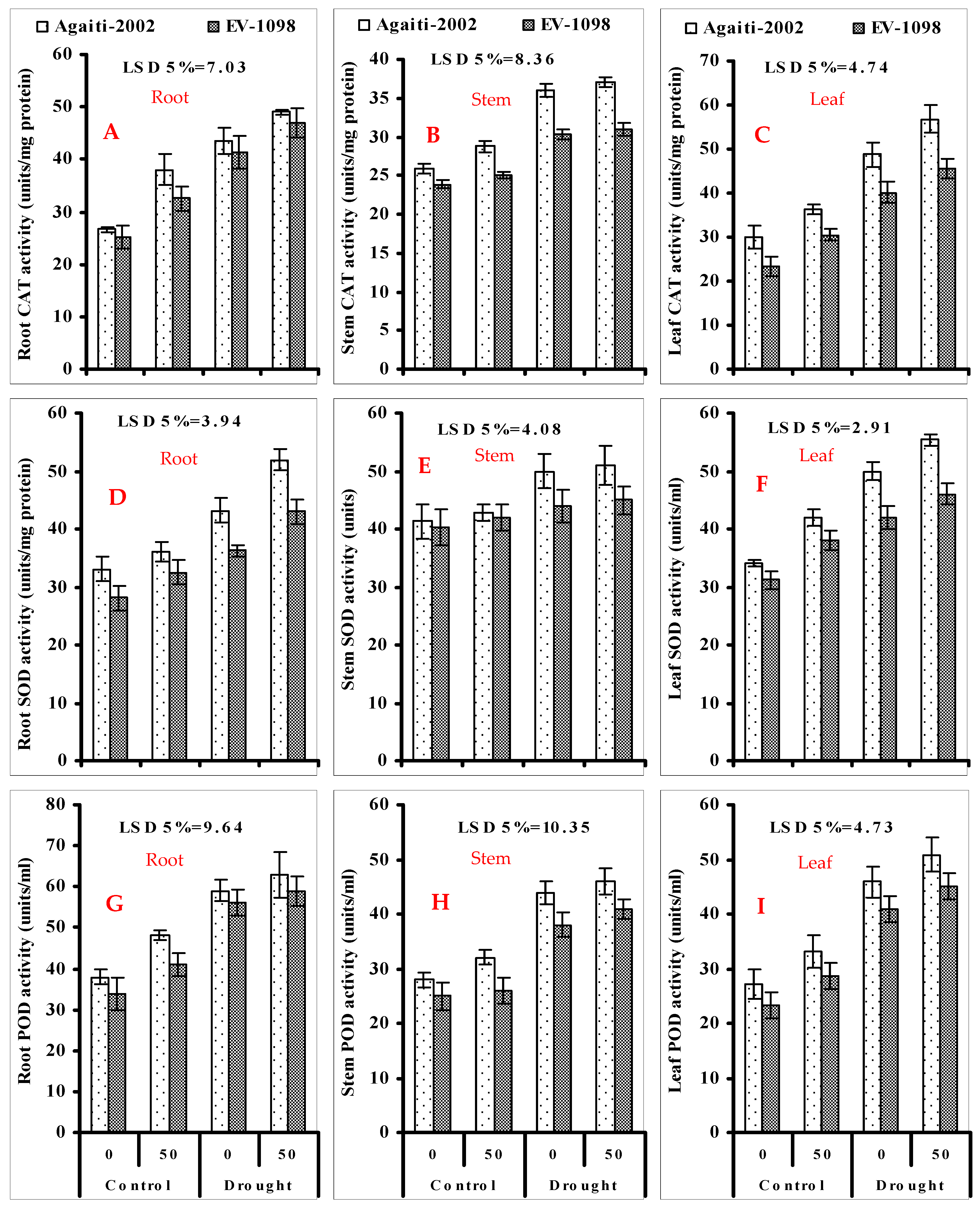
3.4. Activities of CAT (Figure 2A–C), SOD (Figure 2D–F) and POD (2G–I) in Root, Stem, and Leaf of Maize Plants Foliar-Applied with α-Toc
3.5. Contents of K, Ca, Mg, N, and P in Different Parts of Maize Plants Foliary-Applied with α-Toc When Grown under Different Water Regimes
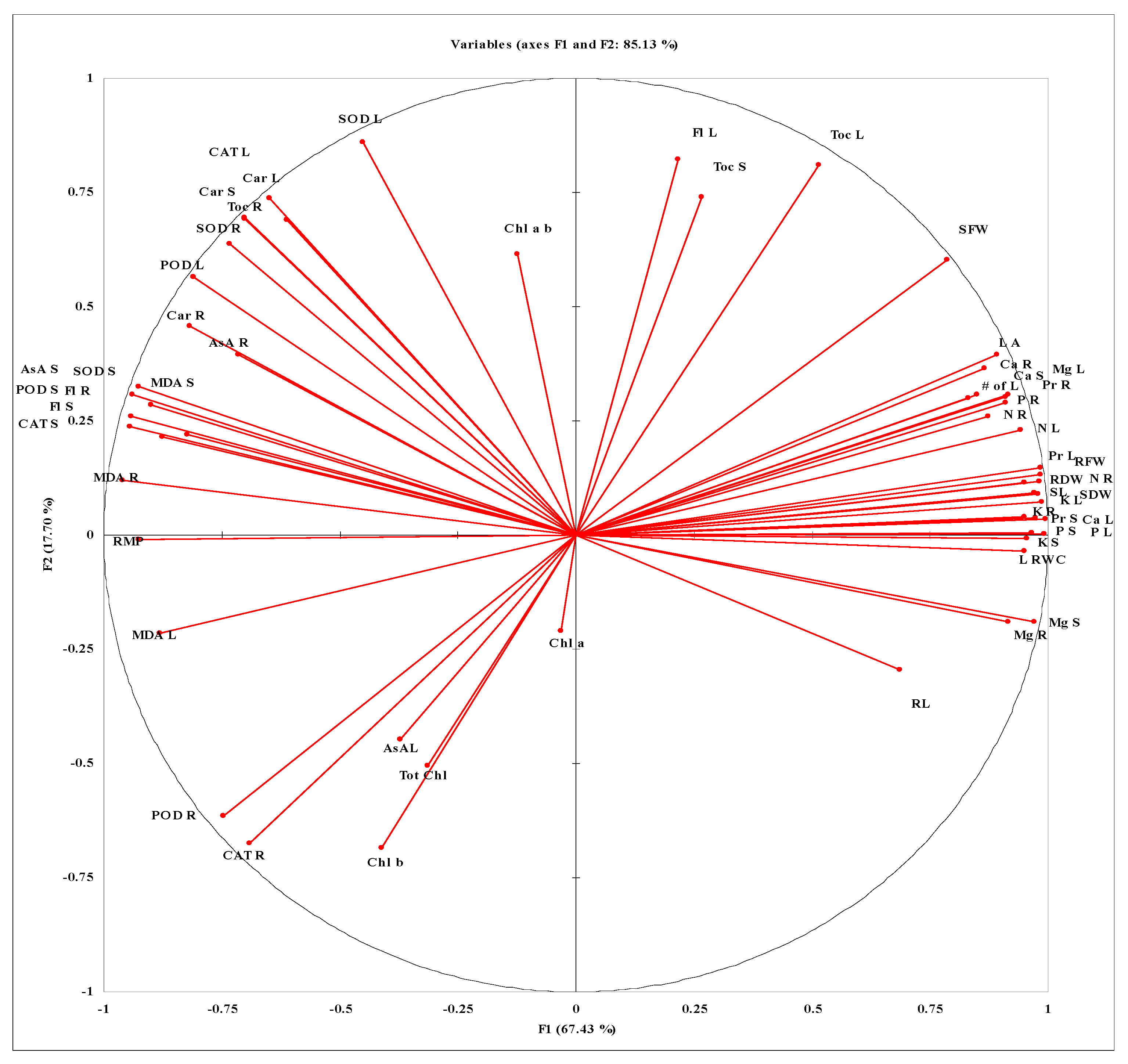
3.6. PCA Analysis and Spearman’s Correlation Coefficient (r2) Values Extracted from XLSTAT Software of All the Studied Attributes of Maize Plants Foliar-Applied with α-Toc
4. Discussion
4.1. Tocopherol Content in Different Plant Parts
4.2. Growth, Water Relations, and Photosynthetic Pigments
4.3. Lipid Peroxidation and Antioxidative Defence Mechanism
4.4. Uptake of Mineral Nutrients
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Toc L | leaf tocopherol |
| Toc R | root tocopherol |
| Toc S | stem tocopherol |
| SL | shoot length |
| RL | root length |
| NL | number of leaves |
| LA | leaf area |
| SFW | shoot fresh weight |
| SDW | shoot dry weight |
| RFW | root fresh weight |
| RDW | root dry weight |
| L RWC | leaf relative water content |
| Chl. a | chlorophyll a |
| Chl. b | chlorophyll b |
| Chl. a/b | chlorophyll a/b ratio |
| Tot Chl. | total chlorophyll |
| RMP | relative membrane permeability |
| MDA L | MDA leaf |
| MDA R | MDA root |
| MDA S | MDA stem |
| Protien L | protein leaf |
| Protein R | protein root |
| Protein S | protein stem |
| AsAL | ascorbic acid leaf |
| AsA R | ascorbic acid root |
| AsA S | ascorbic acid stem |
| Car L | carotenoids leaf |
| Car R | carotenoids root |
| Car S | carotenoids stem |
| Flav L | flavonoids leaf |
| Flav R | flavonoids root |
| Flav S | flavonoids stem |
| CAT L | catalase leaf |
| CAT R | catalase root |
| CAT S | catalase stem |
| POD L | peroxidase leaf |
| POD R | peroxidase root |
| POD S | peroxidase stem |
| SOD L | superoxide dismutase leaf |
| SOD R | superoxide dismutase root |
| SOD S | superoxide dismutase stem |
| K L | potassium leaf |
| K R | potassium root |
| K S | potassium stem |
| Ca L | calcium leaf |
| Ca R | calcium root |
| Ca S | calcium stem |
| Mg L | magnesium leaf |
| Mg R | magnesium root |
| Mg S | magnesium stem |
| N L | nitrogen leaf |
| N R | nitrogen root |
| N S | nitrogen stem |
| P L | phosphorus leaf |
| P R | phosphorus root |
| P S | phosphorus stem |
References
- Gobin, A. Impact of Heat and Drought Stress on Arable Crop Production in Belgium. Nat. Hazards Earth Syst. Sci. 2012, 12, 1911–1922. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. Rhizobacteria and Abiotic Stress Management. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Springer: Singapore, 2019; pp. 65–80. [Google Scholar]
- Cairns, J.E.; Sanchez, C.; Vargas, M.; Ordoñez, R.; Araus, J.L. Dissecting Maize Productivity: Ideotypes Associated with Grain Yield Under Drought Stress and Well-Watered Conditions. J. Integr. Plant Biol. 2012, 54, 1007–1020. [Google Scholar] [CrossRef] [PubMed]
- Adnan, S.; Ullah, K.; Gao, S.; Khosa, A.H.; Wang, Z. Shifting of Agro Climatic Zones, their Drought Vulnerability, and Precipitation and Temperature Trends in Pakistan. Int. J. Climatol. 2017, 37, 529–543. [Google Scholar] [CrossRef]
- Kazmi, D.H.; Li, J.; Rasul, G.; Tong, J.; Ali, G.; Cheema, S.B.; Liu, L.; Gemmer, M.; Fisherr, T. Statistical Downscaling and Future Scenario Generation of Temperatures for Pakistan Region. Theor. Appl. Climatol. 2015, 120, 341–350. [Google Scholar] [CrossRef]
- Hina, S.; Saleem, F. Historical Analysis (1981–2017) of Drought Severity and Magnitude over a Predominantly Arid Region of Pakistan. Clim. Res. 2019, 78, 189–204. [Google Scholar] [CrossRef]
- Um, M.; Kim, Y.; Park, D.; Jung, K.; Wang, Z.; Kim, M.M.; Shin, H. Impacts of Potential Evapotranspiration on Drought Phenomena in Different Regions and Climate Zones. Sci. Total Environ. 2020, 703, 135590. [Google Scholar] [CrossRef]
- Ali, Q.; Anwar, F.; Ashraf, M.; Saari, N.; Perveen, R. Ameliorating Effects of Exogenously Applied Proline on Seed Composition, Seed Oil Quality and Oil Antioxidant Activity of Maize (Zea mays L.) Under Drought Stress. Int. J. Mol. Sci. 2013, 14, 818–835. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Babar, M.A. Comparative physiological and metabolic analysis reveals a complex mechanism involved in drought tolerance in chickpea (Cicer arietinum L.) induced by PGPR and PGRs. Sci. Rep. 2019, 9, 1–19. [Google Scholar] [CrossRef]
- Rana, V.; Singh, D.; Dhiman, R.; Chaudhary, H.K. Evaluation of Drought Tolerance Among Elite Indian Bread Wheat Cultivars. Cereal. Res. Commun. 2014, 42, 91–101. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding Plant Responses to Drought From Genes to the Whole Plant. Physiol. Mol. Biol. Plants 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Rathinasabapathi, B.; Babar, M.A. UPLC-HRMS-based untargeted metabolic profiling reveals changes in chickpea (Cicer arietinum) metabolome following long-term drought stress. Plant Cell Environ. 2019, 42, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Demirevska, K.; Simova-Stoilova, L.; Fedina, I.; Georgieva, K.; Kunert, K. Response of Oryza cystatin L. Transformed Tobacco Plants to Drought, Heat and Light Stress. J. Agron. Crop Sci. 2010, 196, 90–99. [Google Scholar] [CrossRef]
- Ali, Q.; Javed, M.T.; Noman, A.; Haider, M.Z.; Waseem, M.; Iqbal, N.; Perveen, R. Assessment of Drought Tolerance in Mung Bean Cultivars/Lines as Depicted by the Activities of Germination Enzymes, Seedling’s Antioxidative Potential and Nutrient Acquisition. Arch. Agron. Soil. Sci. 2018, 64, 84–102. [Google Scholar] [CrossRef]
- Chen, J.; Junying, D. Effect of Drought on Photosynthesis and Grain Yield of Corn Hybrids With Different Drought Tolerance. Acta-Agron Sin. 1996, 22, 757–762. [Google Scholar]
- Khan, N.; Bano, A.; Curá, J.A. Role of Beneficial Microorganisms and Salicylic Acid in Improving Rainfed Agriculture and Future Food Safety. Microorganisms 2020, 8, 1018. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic Carbon Assimilation and Associated Metabolism in Relation to Water Deficits in Higher Plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef]
- Rapacz, M.; Kościelniak, J.; Jurczyk, B.; Adamska, A.; Wójcik, M. Different Patterns of Physiological and Molecular Response to Drought in Seedlings of Malt- and Feed-Type Barleys (Hordeum vulgare). J. Agron. Crop Sci. 2010, 196, 9–19. [Google Scholar] [CrossRef]
- Marschner, P. Mineral Nutrition of Higher Plants; Academic Press: London, UK, 2011. [Google Scholar]
- Mukhtar, I.; Shahid, M.A.; Khan, M.W.; Balal, R.M.; Iqbal, M.M.; Naz, T.; Zubair, M.; Ali, H.H. Improving salinity tolerance in chili by exogenous application of calcium and sulphur. Soil Environ. 2016, 1, 35. [Google Scholar]
- Heidari, M.; Karami, V. Effects of Different Mycorrhiza Species on Grain Yield, Nutrient Uptake and Oil Content of Sunflower Under Water Stress. J. Saudi Soc. Agric. Sci. 2014, 13, 9–13. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y.; Ahmad, S.M. Improving Agricultural Water Use Efficiency by Nutrient Management in Crop Plants. Acta Agric. Scand. B Soil Plant Sci. 2011, 61, 291–304. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Ihsan, M.Z. Crop Production Under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and Drought Stresses in Crop Plants: Implications, Cross Talk, and Potential Management Opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Rajala, A.; Niskanen, M.; Isolahti, M.; Peltonen-Sainio, P. Seed Quality Effects on Seedling Emergence, Plant Stand Establishment and Grain Yield in Two-Row Barley. Agric. Food Sci. 2011, 20, 228–234. [Google Scholar] [CrossRef]
- Singla, S.; Grover, K.; Angadi, S.V.; Schutte, B.; VanLeeuwen, D. Guar Stand Establishment, Physiology, and Yield Responses to Planting Date in Southern New Mexico. Agron. J. 2016, 8, 2289–2300. [Google Scholar] [CrossRef]
- Srivalli, B.; Sharma, G.; Khanna-Chopra, R. Antioxidative Defense System in an Upland Rice Cultivar Subjected to Increasing Intensity of Water Stress Followed by Recovery. Physiol. Plant. 2003, 119, 503–512. [Google Scholar] [CrossRef]
- Ashraf, M. Inducing Drought Tolerance in Plants: Recent Advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef]
- Zhang, L.X.; Li, S.X.; Zhang, H.; Liang, Z.S. Nitrogen Rates and Water Stress Effects on Production, Lipid Peroxidation and Antioxidative Enzyme Activities in Two Maize (Zea mays L.) genotypes. J. Agron. Crop Sci. 2007, 193, 387–397. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Lee, D.J.; Cheema, S.A.; Aziz, T. Comparative Time Course Action of the Foliar Applied Glycinebetaine, Salicylic Acid, Nitrous Oxide, Brassinosteroids and Spermine in Improving Drought Resistance of Rice. J. Agron. Crop Sci. 2010, 196, 336–345. [Google Scholar] [CrossRef]
- Posmyk, M.M.; Kontek, R.; Janas, K.M. Antioxidant Enzymes Activity and Phenolic Compounds Content in Red Cabbage Seedlings Exposed to Copper Stress. Ecotoxicol. Environ. Saf. 2009, 72, 596–602. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Wahid, A.; Cheema, Z.A.; Cheema, M.A.; Khaliq, A. Physiological Role of Exogenously Applied Glycinebetaine to Improve Drought Tolerance in Fine Grain Aromatic Rice (Oryza sativa L.). J. Agron. Crop. Sci. 2008, 194, 325–333. [Google Scholar] [CrossRef]
- Khan, S.; Hasan, M.; Bari, A.; Khan, F. Climate Classification of Pakistan. In Proceedings of the Balwois Conference, Ohrid, Macedonia, 25–29 May 2010; pp. 1–47. [Google Scholar]
- Ali, Q.; Ashraf, M. Induction of Drought Tolerance in Maize (Zea mays L.) Due to Exogenous Application of Trehalose: Growth, Photosynthesis, Water Relations and Oxidative Defence Mechanism. J. Agron. Crop Sci. 2011, 197, 258–271. [Google Scholar] [CrossRef]
- Tayebi-Meigooni, A.; Awang, Y.; Biggs, A.R.; Mohamad, R.; Madani, B.; Ghasemzadeh, A. Mitigation of Salt-Induced Oxidative Damage in Chinese Kale (Brassica alboglabra L.) Using Ascorbic Acid. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2014, 64, 13–23. [Google Scholar]
- Noman, A.; Ali, Q.; Maqsood, J.; Iqbal, N.; Javed, M.T.; Rasool, N.; Naseem, J. Deciphering Physio-Biochemical, Yield, and Nutritional Quality Attributes of Water-Stressed Radish (Raphanus sativus L.) Plants Grown From Zn-Lys Primed Seeds. Chemosphere 2018, 195, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bano, A.; Zandi, P. Effects of exogenously applied plant growth regulators in combination with PGPR on the physiology and root growth of chickpea (Cicer arietinum) and their role in drought tolerance. J. Plant Interact. 2018, 1, 239–247. [Google Scholar] [CrossRef]
- Khan, N.; Zandi, P.; Ali, S.; Mehmood, A.; Adnan Shahid, M.; Yang, J. Impact of salicylic acid and PGPR on the drought tolerance and phytoremediation potential of Helianthus annus. Front. Microbiol. 2018, 9, 2507. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Ali, S.; Babar, M.A. Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stresses. Plant Growth Regul. 2020, 90, 189–203. [Google Scholar] [CrossRef]
- Jamil, S.; Ali, Q.; Iqbal, M.; Javed, M.T.; Iftikhar, W.; Shahzad, F.; Perveen, R. Modulations in Plant Water Relations and Tissue-Specific Osmoregulation by Foliar-Applied Ascorbic Acid and the Induction of Salt Tolerance in Maize Plants. Braz. J. Bot. 2015, 38, 527–538. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant Cellular and Molecular Responses to High Salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef]
- Fritsche, S.; Wang, X.; Jung, C. Recent Advances in Our Understanding of Tocopherol Biosynthesis in Plants: An Overview of Key Genes, Functions, and Breeding of Vitamin E Improved Crops. Antioxidants 2017, 6, 99. [Google Scholar] [CrossRef]
- Sakr, M.T.; El-Metwally, M.A. Alleviation of the Harmful Effects of Soil Salt Stress on Growth, Yield and Endogenous Antioxidant Content of Wheat Plant by Application of Antioxidants. Pak. J. Biol. Sci. 2009, 12, 624–630. [Google Scholar] [CrossRef]
- Sadak, S.M.; Dawood, S. Role of Ascorbic Acid and α Tocopherol in Alleviating Salinity Stress on Flax Plant (Linum usitatissimum L.). J. Stress Physiol. Biochem. 2014, 10, 93–111. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Saleem, A.; Saleem, U.; Subhani, G.M. Correlation and Path Coefficient Analysis in Maize (Zea mays L.). J. Agric. Sci. 2007, 45, 177–183. [Google Scholar]
- Aslam, M.; Zamir, M.S.I.; Anjum, S.A.; Khan, I.; Tanveer, M. An Investigation into Morphological and Physiological Approaches to Screen Maize (Zea mays L.) Hybrids for Drought Tolerance. Cereal Res. Commun. 2015, 43, 41–51. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I.; et al. Drought Induced Changes in Growth, Osmolyte Accumulation and Antioxidant Metabolism of Three Maize Hybrids. Front. Plant Sci. 2017, 8, 69. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper Enzyme in Isolated Chloroplast Polyphenol Oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar]
- Kirk, J.T.O.; Allen, R.L. Dependence of Chloroplast Pigment Synthesis on Protein Synthesis: Effect of Actidione. Biochem. Biophys. Res. Commun. 1965, 21, 523–530. [Google Scholar] [CrossRef]
- Yang, G.; Rhodes, D.; Joly, R.J. Effects of High Temperature on Membrane Stability and Chlorophyll Fluorescence in Glycinebetaine-Deficient and Glycinebetaine-Containing Maize Lines. Physiol. Mol. Biol. Plants 1996, 23, 437–443. [Google Scholar] [CrossRef]
- Cakmak, I.; Horst, W.J. Effect of Aluminium on Lipid Peroxidation, Superoxide Dismutase, Catalase, and Peroxidase Activities in Root Tips of Soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Bradford, M.M.A. Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Occurrence in Higher Plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Maehly, A.C. Assay of Catalase and Peroxidase. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of Water Stress-Induced Changes in the Levels of Endogenous Ascorbic Acid and Hydrogen Peroxide in Vigna Seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Karadeniz, F.; Burdurlu, H.S.; Koca, N.; Soyer, Y. Antioxidant Activity of Selected Fruits and Vegetables Grown in Turkey. Turk. J. Agric. For. 2005, 29, 297–303. [Google Scholar]
- Backer, H.; Frank, O.; De Angelis, B.; Feingold, S. Plasma Tocopherol in Man at Various Times After Ingesting Free or Acetylated Tocopherol. Nutr. Rep. Int. 1980, 21, 531–536. [Google Scholar]
- Bremner, J.M.; Keeney, D.R. Steam Distillation Methods for Determination of Ammonium, Nitrate and Nitrite. Anal. Chim. Acta 1965, 32, 485–495. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, R.; Nayyar, H. α-Tocopherol Application Modulates the Response of Wheat (Triticum aestivum L.) Seedlings to Elevated Temperatures by Mitigation of Stress Injury and Enhancement of Antioxidants. J. Plant Growth Regul. 2012, 32, 307–314. [Google Scholar] [CrossRef]
- Ali, Q.; Ali, S.; Iqbal, N.; Javed, M.T.; Rizwan, M.; Khaliq, R.; Wijaya, L. Alpha-Tocopherol Fertigation Confers Growth Physio-Biochemical and Qualitative Yield Enhancement in Field Grown Water Deficit Wheat (Triticum aestivum L.). Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Taize, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2015. [Google Scholar]
- Foryer, C.; Noctor, G. Oxygen Processing in Photosynthesis: Regulation and Signaling. New Phytol. 2000, 146, 359–388. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. The Function of Tocopherols and Tocotrienols in Plants. Crit. Rev. Plant Sci. 2002, 21, 31–57. [Google Scholar] [CrossRef]
- Trebst, A.; Depka, B.; Holländer-Czytko, H. A Specific Role for Tocopherol and of Chemical Singlet Oxygen Quenchers in the Maintenance of Photosystem II Structure and Function in Chlamydomonas reinhardtii. FEBS Lett. 2002, 516, 156–160. [Google Scholar] [CrossRef]
- El-Quesni, F.E.M.; Abd El-Aziz, N.G.; Kandil, M.M. Some Studies on the Effect of Ascorbic Acid and a-Tocopherol on the Growth and Some Chemical Composition of Hibiscus rosa sinensis L. at Nubaria. Ozean J. Appl. Sci. 2009, 2, 159–167. [Google Scholar]
- Sadak, M.S.; Rady, M.M.; Badr, N.M.; Gaballah, M.S. Increasing Sun Flower Salt Tolerance Using Nicotinamide and a-Tocopherol. Int. J. Acad. Res. 2010, 2, 263–270. [Google Scholar]
- Javed, M.T.; Greger, M. Cadmium Triggers Elodea canadensis to Change the Surrounding Water pH and Thereby Cd uptake. Int. J. Phytoremediation 2010, 13, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Orabi, S.A.; Abdelhamid, M.T. Protective Role of α-Tocopherol on Two Vicia faba Cultivars Against Seawater-Induced Lipid Peroxidation by Enhancing Capacity of Anti-Oxidative System. J. Saudi Soc. Agric. Sci. 2016, 15, 145–154. [Google Scholar] [CrossRef]
- Shigeoka, S.; Ishikawa, T.; Tamoi, M.; Miyagawa, Y.; Takeda, T.; Yabuta, Y.; Yoshimura, K. Regulation and Function of Ascorbate Peroxidase Isoenzymes. J. Exp. Bot. 2002, 53, 1305–1319. [Google Scholar] [CrossRef]
- Ali, Q.; Haider, M.Z.; Iftikhar, W.; Jamil, S.; Javed, M.T.; Noman, A.; Perveen, R. Drought Tolerance Potential of Vigna mungo L. Lines as Deciphered by Modulated Growth, Antioxidant Defense, and Nutrient Acquisition Patterns. Braz. J. Bot. 2016, 39, 801–812. [Google Scholar] [CrossRef]
- Maeda, H.; Sakuragi, Y.; Bryant, D.A.; DellaPenna, D. Tocopherols Protect Synechocystis sp. Strain PCC 6803 From Lipid Peroxidation. Plant Physiol. 2005, 138, 1422–1435. [Google Scholar] [CrossRef]
- Sattler, S.E.; Cahoon, E.B.; Coughlan, S.J.; DellaPenna, D. Characterization of Tocopherol Cyclases From Higher Plants and Cyanobacteria. Evolutionary Implications for Tocopherol Synthesis and Function. Plant Physiol. 2003, 132, 2184–2195. [Google Scholar] [CrossRef]
- Wang, X.; Quinn, P.J. Preferential Interaction of α-Tocopherol With Phosphatidylcholines in Mixed Aqueous Dispersions of Phosphatidylcholine and Phosphatidylethanolamine. Eur. J. Biochem. 2000, 267, 6362–6368. [Google Scholar] [CrossRef]
- Maeda, H.; DellaPenna, D. Tocopherol Functions in Photosynthetic Organisms. Curr. Opin. Plant Biol. 2007, 10, 260–265. [Google Scholar] [CrossRef]
- Fukuzawa, K.; Matsuura, K.; Tokumura, A.; Suzuki, A.; Terao, J. Kinetics and Dynamics of Singlet Oxygen Scavenging by α-Tocopherol in Phospholipid Model Membranes. Free Radic. Biol. Med. 1997, 22, 923–930. [Google Scholar] [CrossRef]
- Kobayashi, N.; DellaPenna, D. Tocopherol Metabolism, Oxidation and Recycling Under High Light Stress in Arabidopsis. Plant J. 2008, 55, 607–618. [Google Scholar] [CrossRef]
- Fahrenholtz, S.R.; Doleiden, F.H.; Trozzolo, A.M.; Lamola, A.A. On the Quenching of Singlet Oxygen by α-Tocopherol. J. Photochem. Photobiol. Biol. 1974, 20, 505–509. [Google Scholar] [CrossRef]
- Semida, W.M.; Taha, R.S.; Abdelhamid, M.T.; Rady, M.M. Foliar-Applied α-Tocopherol Enhances Salt-Tolerance in Vicia faba L. Plants Grown Under Saline Conditions. S. Afr. J. Bot. 2014, 95, 24–31. [Google Scholar] [CrossRef]
- Khattab, H. Role of Glutathione and Polyadenylic Acid on the Oxidative Defense Systems of Two Different Cultivars of Canola Seedlings Grown Under Saline Conditions. Aust. J. Basic Appl. Sci. 2007, 1, 323–334. [Google Scholar]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid Oxidation in Plants: From Biochemical Properties to Physiological Functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef]
- Shao, H.B.; Chu, L.Y.; Shao, M.A.; Jaleel, C.A.; Hong-mei, M. Higher Plant Antioxidants and Redox Signaling Under Environmental Stresses. C. R. Biol. 2008, 331, 433–441. [Google Scholar] [CrossRef]
- Hu, Y.; Schmidhalter, U. Drought and Salinity: A Comparison of Their Effects on Mineral Nutrition of Plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| α-Toc | Cultivars | SL (cm) | RL (cm) | NOL/plant | PLA (cm2) | SFW (g) | SDW (g) | RFW (g) | RDW (g) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 0 mmol | Agaiti-2002 | 12.23 ± 1.10 a | 7.77 ± 0.48 a | 3.75 ± 0.20 b | 69.10 ± 5.73 bc | 2.92 ± 0.215 b | 0.37 ± 0.04 a | 1.18 ± 0.15 a | 0.13 ± 0.019 b |
| EV-1098 | 12.03 ± 0.67 a | 6.70 ± 0.50 ab | 3.85 ± 0.20 b | 62.40 ± 4.04 cd | 2.76 ± 0.224 bc | 0.32 ± 0.03 abc | 1.18 ± 0.11 a | 0.13 ± 0.022 b | ||
| 50 mmol | Agaiti-2002 | 13.33 ± 0.82 a | 6.20 ± 0.35 bc | 4.25 ± 0.30 a | 79.98 ± 5.08 a | 3.47 ± 0.402 a | 0.40 ± 0.03 a | 1.39 ± 0.14 a | 0.16 ± 0.014 a | |
| EV-1098 | 12.85 ± 1.11 a | 7.50 ± 0.79 a | 4.50 ± 0.35 a | 73.60 ± 0.63 ab | 3.74 ± 0.140 a | 0.38 ± 0.03 a | 1.35 ± 0.15 a | 0.17 ± 0.024 a | ||
| Drought | 0 mmol | Agaiti-2002 | 8.27 ± 1.11 d | 4.67 ± 0.49 cd | 3.25 ± 0.13 c | 45.50 ± 1.06 f | 2.28 ± 0.145 c | 0.23 ± 0.03 bd | 0.74 ± 0.13 c | 0.09 ± 0.021 d |
| EV-1098 | 8.45 ± 0.66 d | 4.00 ± 0.39 d | 3.00 ± 0.35 c | 47.50 ± 1.13 ef | 2.37 ± 0.135 c | 0.24 ± 0.01 cd | 0.78 ± 0.12 c | 0.10 ± 0.011 cd | ||
| 50 mmol | Agaiti-2002 | 10.30 ± 1.51 b | 5.15 ± 0.59 e | 3.67 ± 0.20 b | 55.00 ± 1.74 de | 2.56 ± 0.136 b | 0.27 ± 0.01 cd | 0.93 ± 0.16 b | 0.11 ± 0.007 bcd | |
| EV-1098 | 9.77 ± 1.16 c | 4.73 ± 0.37 cd | 3.75 ± 0.25 b | 54.50 ± 1.07 def | 2.72 ± 0.139 b | 0.28 ± 0.03 bcd | 0.99 ± 0.12 b | 0.12 ± 0.018 bc | ||
| LSD 5% | 1.311 | 1.11 | 0.414 | 4.26 | 0.50 | 0.081 | 0.15 | 0.025 | ||
| SOV | d.f | SL | RL | NOL | LA | SFW | SDW | RFW | RDW |
| WS | 1 | 66.7 *** | 34.08 ** | 2.37 ** | 2440 *** | 2.85 *** | 0.045 ** | 1.06 *** | 0.011 *** |
| Toc | 1 | 8.13 ** | 0.02 ns | 2.01 ** | 41.5 *** | 1.67 ** | 0.003* | 0.23 ns | 0.004 *** |
| CV | 1 | 0.53 ns | 0.42 ns | 0.01 ns | 37.5 * | 0.04 ns | 0.007 ns | 0.002 ns | 5.04 × 10−4 ns |
| WS * Toc | 1 | 0.54 ns | 0.88 ns | 0.02 ns | 8.16 ns | 0.34 ns | 0.004 ns | 6 × 10−4 ns | 3.37 × 10−4 ns |
| WS * CV | 1 | 0.16 ns | 0.66 ns | 0.17 ns | 121 *** | 0.001 ns | 0.014 ns | 0.010 ns | 1.04 × 10−4 ns |
| Toc * CV | 1 | 0.67 ns | 1.92 ns | 0.09 ns | 0.17 ns | 0.08 ns | 0.002 ns | 2.7 × 10−4 ns | 1.04 × 10−4 ns |
| WS * Toc * CV | 1 | 0.10 ns | 2.66 ns | 0.04 ns | 0.17 ns | 0.06 ns | 0.002 ns | 0.002 ns | 1.04 × 10−4 ns |
| Error | 16 | 0.78 | 3.31 | 0.17 | 7.08 | 0.13 | 0.003 | 0.065 | 2.17 × 10−4 |
| SOV | d.f | Protein L | Protein R | Potein S | LRWC | LRMP | Chl. a | Chl. b | Chl a/b |
| WS | 1 | 46728 *** | 6834 *** | 2281 *** | 3314 *** | 517 *** | 0.15 *** | 0.043 *** | 1.270 *** |
| Toc | 1 | 11051 *** | 3978 *** | 253 ** | 172 ** | 50.0* | 0.10 *** | 0.015 *** | 0.130 ns |
| CV | 1 | 651.04 ** | 1395 ** | 13.5 ns | 118 * | 0.18 ns | 0.01 ns | 2.8 × 10−4 ns | 0.004 ns |
| WS * Toc | 1 | 35.04 ns | 234 *** | 181.5 ** | 157 *** | 76.5 * | 2.4 × 10−6 ns | 0.004 ** | 0.290 * |
| WS * CV | 1 | 1717 *** | 513 *** | 37.5 ns | 22.6ns | 0.80 ns | 4.8 × 10−4 ns | 2.2 × 10−5 ns | 0.003 ns |
| Toc * CV | 1 | 1785 *** | 18.4 ns | 37.5 ns | 1.25 ns | 16.1 ns | 0.002 ns | 5.8 × 10−5 ns | 0.010 ns |
| WS * Toc * CV | 1 | 3.37 ns | 408 *** | 1.5 ns | 6.56 ns | 0.19 ns | 1.14 × 10−54 ns | 0.002 * | 0.199 ns |
| Error | 16 | 62.83 | 18 | 19.5 | 17.06 | 10.8 | 0.002 | 3.1 × 10−4 | 0.051 |
| SOV | d.f | Total Chl. | POD R | POD S | POD L | CAT R | CAT S | CAT L | SOD S |
| WS | 1 | 0.351 *** | 2223 *** | 260 *** | 1932 *** | 1220 *** | 329.8 *** | 2011 *** | 270 *** |
| Toc | 1 | 0.190 *** | 198 *** | 37.6 *** | 146 *** | 359 *** | 12.01 ns | 268 *** | 4.74 ns |
| CV | 1 | 0.008 ns | 135 ** | 149 * | 157 ** | 45.8 ** | 126.1 ** | 414 *** | 71.4 * |
| WS * Toc | 1 | 0.004 ns | 30.4 ns | 1.7 × 10−4 ns | 0.89 ns | 18.6 ns | 1.054 ns | 0.219 * | 1.1 ns |
| WS * CV | 1 | 7 × 10−4 ns | 9.4 ns | 1.50 ns | 1.21 ns | 3.2 ns | 12.07 ns | 19.65 * | 27.4 ns |
| Toc * CV | 1 | 0.001 ns | 9.3 ns | 1.53 ns | 1.21 ns | 3.2 ns | 1.054 ns | 0.22 ns | 0.39 ns |
| WS * Toc * CV | 1 | 0.003 ns | 3.4 ns | 5.93 ns | 0.08 ns | 3.5 ns | 1.032 ns | 2.85 ns | 0.018 ns |
| Error | 16 | 0.002 | 9.5 | 7.10 | 12.1 | 6.9 | 8.286 | 8.91 | 8.485 |
| SOV | d.f | SOD L | SOD R | MDA S | MDA L | MDA R | TOC S | TOC L | TOC R |
| WS | 1 | 1229 *** | 7222 *** | 3174 *** | 1890 *** | 3313 *** | 1493 *** | 1666 *** | 1741 *** |
| Toc | 1 | 86.98 * | 176 ** | 73.5 ** | 828 *** | 793 *** | 83.7 ** | 1148 *** | 489 *** |
| CV | 1 | 81.40 * | 218 ** | 4.0 * | 84.3 *** | 384 *** | 9.0 ns | 60.11 * | 55.2 * |
| WS * Toc | 1 | 107 ** | 24.7 ns | 1.5 ns | 108 *** | 726 *** | 3.24 ns | 170 ** | 1.30 ns |
| WS * CV | 1 | 0.06 ns | 16.1 ns | 6.0 ns | 18.3 ns | 337 *** | 0.30 ns | 2.66 ns | 1.71 ns |
| Toc * CV | 1 | 62.3 * | 0.05 ns | 1.5 ns | 30.3 * | 253 *** | 0.42 ns | 0.16 ns | 1.71 ns |
| WS * Toc * CV | 1 | 28.8 ns | 0.80 ns | 1.5 ns | 18.3ns | 216 *** | 3.24 ns | 0.66 ns | 0.01 ns |
| Error | 16 | 10.9 | 15.65 | 4.9 | 5.12 | 8.62 | 5.52 | 12.0 | 11.3 |
| SOV | d.f | AsA S | AsA L | AsA R | Car S | Car L | Car R | Flav S | Flav L |
| WS | 1 | 14259 *** | 23814 *** | 26498 *** | 5953 *** | 10688 *** | 5730 *** | 45.37 ** | 84.38 *** |
| Toc | 1 | 1053.*** | 14113 *** | 4858 *** | 1722 *** | 4776 *** | 273 ns | 0.37 ns | 18.38 * |
| CV | 1 | 630 *** | 4056 *** | 8720 *** | 67.3 ns | 412.7 * | 894 * | 9.37 ns | 9.375 ns |
| WS * Toc | 1 | 108 * | 937 *** | 881 *** | 146 ns | 265 | 8.77 ns | 0.38 ns | 3.35 ns |
| WS * CV | 1 | 63.3 ns | 0.01 ns | 930 *** | 2.53 ns | 39.9 | 6.20 ns | 0.37 ns | 0.37 ns |
| Toc * CV | 1 | 30.4 ns | 1.5 ns | 12.7 ns | 0.26 ns | 27.9 | 8.19 ns | 0.38 ns | 0.38 ns |
| WS * Toc * CV | 1 | 30.3 ns | 253 * | 304 ** | 0.25 ns | 9.9 | 3.04 ns | 0.37 ns | 0.37 ns |
| Error | 16 | 8.62 | 31.75 | 29.7 | 59.4 | 64.9 | 160.6 | 4.5 | 3.5 |
| SOV | d.f | Flav R | K S | K L | K R | Ca S | Ca L | Ca R | Mg S |
| WS | 1 | 135.4 *** | 261 ** | 403 *** | 396 *** | 56.8 *** | 172 *** | 32.08 *** | 0.014 ns |
| Toc | 1 | 30.3 * | 53.2 ns | 68.2 * | 45.7 ns | 1.67 ns | 15.6 *** | 14.7 ** | 0.144 ns |
| CV | 1 | 9.37 ns | 2.83 ns | 1.82 ns | 0.55 ns | 2.66 * | 4.61 * | 0.26 ns | 0.768 ns |
| WS * Toc | 1 | 3.375 ns | 0.15 ns | 2.95 ns | 0.40 ns | 6.36 ** | 0.90 ns | 0.011 ns | 1.264 * |
| WS * CV | 1 | 3.375 ns | 7.67 ns | 0.12 ns | 0.22 ns | 2.11 ns | 1.14 ns | 0.206 ns | 9 × 10−4 ns |
| Toc * CV | 1 | 0.375 ns | 8.89 ns | 11.5 ns | 2.01 ns | 0.77 ns | 0.98 ns | 0.061 ns | 1.92 ** |
| WS * Toc * CV | 1 | 0.375 ns | 0.81 ns | 0.26 ns | 1.02 ns | 0.85 ns | 0.14 ns | 0.160 ns | 0.069 ns |
| Error | 16 | 4.125 | 20.1 ns | 11.9 | 14.2 | 0.57 | 0.79 | 1.035 | 0.221 |
| SOV | d.f | Mg L | Mg R | P S | P L | P R | N S | N L | N R |
| WS | 1 | 0.29 ns | 3.93 *** | 10.5 *** | 11.7 *** | 5.15 * | 260 ** | 321.84 *** | 368.8 *** |
| Toc | 1 | 0.28 ns | 0.02 ns | 2.45 * | 1.15 * | 1.56 ns | 52.3 ns | 45.49 ns | 109.4 * |
| CV | 1 | 2.86 *** | 0.01 ns | 0.40 ns | 0.02 ns | 0.36 ns | 1.53 ns | 26.44 ns | 36.03 ns |
| WS * Toc | 1 | 0.056 ns | 0.20 ns | 0.34 ns | 0.295 ns | 0.43 ns | 0.58 ns | 3.37 ns | 1.89 ns |
| WS * CV | 1 | 2.95 *** | 0.16 ns | 0.32 ns | 0.039 ns | 0.001 ns | 1.89 ns | 3.26 ns | 3.07 ns |
| Toc * CV | 1 | 0.43 ns | 0.04 ns | 0.30 ns | 0.005 ns | 0.36 ns | 1.73 ns | 2.72 ns | 3.50 ns |
| WS * Toc * CV | 1 | 1.23 ** | 0.12 ns | 1.11 ns | 0.324 ns | 0.35 ns | 1.81 ns | 1.52 ns | 2.14 ns |
| Error | 16 | 0.11 | 0.13 | 0.51 | 0.203 | 0.69 | 21.4ns | 11.65 | 19.3 |
| Stress | α-Toc | Cultivars | Chl. a (mg/g FW) | Chl. b (mg/g FW) | Chl a/b | Total Chl. (mg/g FW) | Leaf Car (µg/g FW) | Root Car (µg/g FW) | Stem Car (µg/g FW) |
|---|---|---|---|---|---|---|---|---|---|
| Control | 0 mmol | Agaiti-2002 | 1.52 ± 0.11 ab | 0.36 ± 0.01 bc | 4.53 ± 0.16 a | 1.88 ± 0.12 ab | 102.16 ± 1.78 e | 99.26 ± 9.60 de | 82.70 ± 5.97 e |
| EV-1098 | 1.47 ± 0.18 bc | 0.38 ± 0.02 bc | 3.87 ± 0.25 d | 1.85 ± 0.17 b | 93.52 ± 4.41 f | 84.16 ± 12.26 f | 80.41 ± 6.24 e | ||
| 50 mmol | Agaiti-2002 | 1.63 ± 0.06 a | 0.47 ± 0.01 a | 4.40 ± 0.14 ab | 2.00 ± 0.16 ab | 120.27 ± 13.12 d | 105.34 ± 10.99 d | 105.00 ± 2.83 d | |
| EV-1098 | 1.60 ± 0.14 ab | 0.44 ± 0.01 ab | 3.64 ± 0.33 ed | 2.04 ± 0.18 a | 118.23 ± 4.89 d | 94.79 ± 6.55 e | 101.89 ± 4.40 d | ||
| Drought | 0 mmol | Agaiti-2002 | 1.34 ± 0.04 cd | 0.33 ± 0.02 c | 3.62 ± 0.09 e | 1.67 ± 0.05 b | 139.98 ± 10.40 b | 129.65 ± 9.31 ab | 120.00 ± 5.83 bc |
| EV-1098 | 1.31 ± 0.12 bc | 0.31 ± 0.02 c | 4.28 ± 0.05 bc | 1.62 ± 0.10 c | 127.25 ± 5.61 c | 118.75 ± 2.98 c | 116.00 ± 5.75 c | ||
| 50 mmol | Agaiti-2002 | 1.46 ± 0.12 bcd | 0.34 ± 0.02 c | 4.29 ± 0.16 bc | 1.80 ± 0.10 b | 176.08 ± 2.74 a | 134.73 ± 11.52 a | 132.00 ± 6.54 a | |
| EV-1098 | 1.46 ± 0.09 bcd | 0.35 ± 0.02 c | 4.17 ± 0.16 c | 1.81 ± 0.12 b | 163.33 ± 5.61 b | 124.26 ± 9.60b c | 128.00 ± 4.89 ab | ||
| LSD 5% | 0.15 | 0.12 | 0.64 | 0.52 | 8.74 | 11.81 | 8.15 | ||
| Stress | α-Toc | Cultivars | LRWC (%) | RMP (%) | Leaf TSP (mg/kg FW) | Root TSP (mg/kg FW) | Stem TSP (mg/kg FW) | Leaf MDA (ոmol/g FW) | Root MDA (ոmol/g FW) | Stem MDA (ոmol/g FW) |
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 0 mmol | Agaiti-2002 | 87.54 ± 1.71 a | 32.33 ± 3.92 e | 210.00 ± 29.89 f | 144.33 ± 7.50 f | 73.00 ± 15.15 c | 68.10 ± 1.19 bc | 66.18 ± 6.40 c | 62.02 ± 4.48 cd |
| EV-1098 | 81.73 ± 5.00 b | 33.96 ± 2.33 de | 233.33 ± 23.34 e | 160.33 ± 4.58 e | 72.00 ± 9.36 c | 62.35 ± 2.94 c | 66.10 ± 8.17 c | 60.31 ± 5.18 cd | ||
| 50 mmol | Agaiti-2002 | 88.36 ± 1.62 a | 34.83 ± 2.76 d | 267.33 ± 24.22 d | 186.67 ± 2.57 cd | 83.67 ± 9.01 b | 60.18 ± 8.75 c | 65.23 ± 7.33 c | 60.35 ± 2.12 c | |
| EV-1098 | 81.38 ± 5.36 b | 33.55 ± 3.19 de | 257.00 ± 18.26 e | 182.00 ± 3.45 d | 86.67 ± 14.81 b | 55.49 ± 3.26 bc | 66.53 ± 4.37 c | 56.42 ± 3.29 c | ||
| Drought | 0 mmol | Agaiti-2002 | 58.23 ± 3.52 d | 45.32 ± 0.50 b | 312.33 ± 31.67 c | 183.67 ± 10.05 d | 101.67 ± 16.96 a | 90.65 ± 6.94 a | 99.83 ± 6.21 a | 85.59 ± 4.37 ab |
| EV-1098 | 51.34 ± 3.09 e | 47.88 ± 3.78 a | 304.67 ± 25.01 c | 201.33 ± 5.64 b | 94.33 ± 13.58 ab | 84.84 ± 3.74 a | 102.84 ± 1.98 a | 84.04 ± 4.32 b | ||
| 50 mmol | Agaiti-2002 | 67.87 ± 0.62 c | 40.29 ± 2.08 c | 376.67 ± 22.30 a | 196.00 ± 9.98 bc | 99.00 ± 9.93 a | 70.72 ± 1.83 b | 64.3 ± 7.68 b | 81.92 ± 4.91 ab | |
| EV-1098 | 66.17 ± 2.02 c | 39.08 ± 2.44 c | 330.67 ± 33.06 b | 227.67 ± 5.81 a | 98.00 ± 10.98 a | 72.22 ± 3.74 b | 92.50 ± 6.40 b | 80.60 ± 3.67 a | ||
| LSD 5% | 4.06 | 3.49 | 24.44 | 12.64 | 10.26 | 5.82 | 7.88 | 6.12 | ||
| Stress | α-Toc | Cultivars | K+ Leaf (mg g−1 DW) | K+ Root (mg g−1 DW) | K+ stem (mg g−1 DW) | Ca2+ leaf (mg g−1 DW) | Ca2+ Root (mg g−1 DW) | Ca2+ stem (mg g−1 DW) | Mg2+ leaf (mg g−1 DW) | Mg2+ root (mg g−1 DW) | Mg2+ stem (mg g−1 DW) |
| Control | 0 mmol | Agaiti-2002 | 39.30 ± 3.18 a | 41.35 ± 2.99 ab | 33.09 ± 3.20 bc | 11.35 ± 0.20 d | 10.34 ± 0.75 c | 10.03 ± 0.61 b | 3.41 ± 0.06 b | 4.13 ± 0.30 a | 3.31 ± 0.32 ab |
| EV-1098 | 41.17 ± 1.47 ab | 40.21 ± 2.12 b | 31.99 ± 2.38 c | 12.92 ± 0.61 c | 10.05 ± 0.03 c | 9.02 ± 0.43 c | 2.73 ± 0.15 d | 4.02 ± 0.12 a | 3.14 ± 0.31 b | ||
| 50 mmol | Agaiti-2002 | 43.15 ± 2.19 a | 42.90 ± 1.41 a | 35.12 ± 1.67 ab | 14.36 ± 1.46 b | 10.99 ± 0.18 b | 11.71 ± 1.22 a | 4.01 ± 0.44 a | 4.29 ± 0.14 a | 3.51 ± 0.33 a | |
| EV-1098 | 43.67 ± 2.45 a | 42.93 ± 1.77 a | 36.00 ± 1.21 a | 15.33 ± 0.40 a | 12.23 ± 0.31 a | 10.40 ± 0.36 b | 3.27 ± 0.16 bc | 4.09 ± 0.22 a | 3.45 ± 0.07 a | ||
| Drought | 0 mmol | Agaiti-2002 | 30.33 ± 3.47 e | 35.67 ± 2.82 cd | 25.00 ± 1.85 ef | 6.48 ± 0.59 g | 7.33 ± 0.40 e | 5.74 ± 0.71 g | 2.03 ± 0.35 e | 2.64 ± 0.29 b | 2.38 ± 0.22 d |
| EV-1098 | 32.33 ± 1.76 de | 33.00 ± 2.79 e | 24.33 ± 1.76 f | 7.67 ± 0.40 f | 7.67 ± 0.81 f | 6.87 ± 0.72 f | 2.24 ± 0.19 e | 2.34 ± 0.29 b | 2.82 ± 0.26 c | ||
| 50 mmol | Agaiti-2002 | 36.00 ± 1.85 c | 37.64 ± 2.29 c | 27.33 ± 1.76 d | 8.45 ± 0.30 e | 9.15 ± 0.82 d | 8.33 ± 0.40 e | 2.54 ± 0.09 d | 2.66 ± 0.33 b | 2.49 ± 0.31 d | |
| EV-1098 | 34.81 ± 2.19 cd | 34.67 ± 2.45 de | 27.00 ± 3.89 de | 8.33 ± 0.40 e | 9.96 ± 0.66 c | 8.88 ± 0.13 d | 3.11 ± 0.19 c | 2.71 ± 0.25 b | 2.47 ± 0.20 d | ||
| LSD 5% | 2.99 | 2.20 | 2.26 | 0.50 | 0.47 | 0.40 | 0.29 | 0.41 | 0.22 | ||
| Stress | α-Toc | Cultivars | N Leaf (mg g−1 DW) | N Root (mg g−1 DW) | N Stem (mg g−1 DW) | P Leaf (mg g−1 DW) | P Root (mg g−1 DW) | P Stem (mg g−1 DW) | |||
| Control | 0 mmol | Agaiti-2002 | 38.63 ± 2.28 b | 41.35 ± 2.99 ab | 33.09 ± 3.20 b | 5.20 ± 0.33 b | 5.91 ± 0.43 b | 4.50 ± 0.46 c | |||
| EV-1098 | 41.17 ± 1.47 b | 40.21 ± 2.12 ab | 32.72 ± 2.93 b | 5.55 ± 0.25 b | 5.74 ± 0.27 b | 4.01 ± 0.28 de | |||||
| 50 mmol | Agaiti-2002 | 45.82 ± 2.77 a | 42.90 ± 1.41 a | 38.16 ± 1.90 a | 6.06 ± 0.34 a | 6.13 ± 0.20 ab | 5.02 ± 0.22 b | ||||
| EV-1098 | 47.00 ± 2.52 a | 43.93 ± 2.72 a | 39.06 ± 2.73 a | 6.01 ± 0.23 a | 6.28 ± 0.31 a | 5.28 ± 0.49 a | |||||
| Drought | 0 mmol | Agaiti-2002 | 25.33 ± 3.46 d | 33.33 ± 2.45 c | 26.00 ± 2.27 c | 4.33 ± 0.50 cd | 4.62 ± 0.42 d | 3.84 ± 0.22 e | |||
| EV-1098 | 23.33 ± 2.64 d | 34.41 ± 3.00 e | 23.00 ± 3.20 d | 4.06 ± 0.27 d | 4.86 ± 0.50 cd | 3.58 ± 0.25 f | |||||
| 50 mmol | Agaiti-2002 | 30.00 ± 2.10 c | 36.61 ± 3.27 b | 31.00 ± 1.85 b | 4.29 ± 0.27 cd | 5.02 ± 0.47 c | 4.08 ± 0.23 d | ||||
| EV-1098 | 32.67 ± 0.81 c | 37.67 ± 1.45 b | 27.67 ± 2.02 c | 4.54 ± 0.22 c | 5.05 ± 0.53 c | 3.82 ± 0.11 e | |||||
| LSD 5% | 2.95 | 4.00 | 2.80 | 0.39 | 0.32 | 0.22 | |||||
| Toc L | Toc R | Toc S | |
|---|---|---|---|
| Toc L | 1.000 | ||
| Toc R | 0.204 ns | 1.00 | |
| Toc S | 0.847 *** | 0.302 ns | 1.00 |
| SL | 0.534 ** | −0.644 *** | 0.287ns |
| RL | 0.192 ns | −0.661 *** | 0.122 ns |
| NL | 0.797 *** | −0.353 ns | 0.557 ** |
| L A | 0.768 *** | −0.371 ns | 0.664 *** |
| SFW | 0.921 *** | −0.124 ns | 0.661 *** |
| SDW | 0.578 ** | 0.643 *** | 0.368 ns |
| RFW | 0.627 *** | −0.597 ** | 0.381 ns |
| RDW | 0.780 *** | −0.420* | 0.531 ns |
| L RWC | 0.391 ns | 0.721 *** | 0.164 ns |
| Chl. a | −0.088 ns | −0.059 ns | 0.266 ns |
| Chl. b | −0.787 *** | −0.181 ns | −0.423* |
| Chl. a/b | 0.553 ** | 0.538 ** | 0.740 *** |
| Tot Chl. | −0.624 *** | −0.133 ns | −0.239 ns |
| RMP | −0.416* | 0.664 *** | −0.084 ns |
| MDA L | −0.534 ** | −0.463 * | −0.202 ns |
| MDA R | −0.342 ns | −0.751 *** | −0.052 ns |
| MDA S | −0.240 ns | −0.845 *** | −0.037 ns |
| Protien L | 0.561 ** | 0.591 ** | 0.392 ns |
| Protein R | 0.695 *** | 0.324 ns | 0.314 ns |
| Protein S | 0.468 * | 0.666 *** | 0.159 ns |
| AsA L | −0.434 * | −0.115 ns | −0.149 ns |
| AsA R | −0.180 ns | 0.709 *** | −0.023 ns |
| AsA S | −0.196 ns | 0.797 *** | 0.145 ns |
| Car L | 0.191 ns | 0.925 *** | 0.339 ns |
| Car R | −0.078 ns | 0.870 *** | 0.190 ns |
| Car S | 0.218 ns | 0.974 *** | 0.411 ns |
| Flav L | 0.669 *** | 0.443* | 0.426* |
| Flav R | −0.348 ns | 0.679 *** | −0.269 ns |
| Flav S | −0.211 ns | 0.733 *** | 0.047 ns |
| CAT L | 0.275 ns | 0.966 *** | 0.434 * |
| CAT R | −0.884 *** | 0.041 ns | −0.757 *** |
| CAT S | −0.283 ns | 0.863 *** | −0.114 ns |
| POD L | 0.049 ns | 0.961 *** | 0.255 ns |
| POD R | −0.831 *** | 0.116 ns | −0.633 *** |
| POD S | −0.263 ns | 0.872 *** | −0.107 ns |
| SOD L | 0.438 * | 0.936 *** | 0.430 * |
| SOD R | 0.173 ns | 0.951 *** | 0.273 ns |
| SOD S | −0.220 ns | 0.859 *** | 0.047 ns |
| K L | 0.553 ** | 0.606 ** | 0.282 ns |
| K R | 0.555 ** | 0.675 *** | 0.333 ns |
| K S | 0.470 * | 0.673 *** | 0.319 ns |
| Ca L | 0.569 ** | 0.633 *** | 0.339 ns |
| Ca R | 0.674 *** | 0.461 * | 0.416 * |
| Ca S | 0.749 *** | 0.437 * | 0.469 * |
| Mg L | 0.656 *** | 0.439 * | 0.470 * |
| Mg R | 0.372 ns | 0.761 *** | 0.201 ns |
| Mg S | 0.335 ns | 0.833 *** | 0.151 ns |
| N L | 0.610 ** | 0.613 ** | 0.368 ns |
| N R | 0.669 *** | 0.494 * | 0.488 * |
| N R | 0.626 *** | 0.601 ** | 0.337 ns |
| P L | 0.539 ** | 0.683 *** | 0.383 ns |
| P R | 0.729 *** | 0.430* | 0.620 ** |
| P S | 0.536 ** | 0.688 *** | 0.281 ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, Q.; Tariq Javed, M.; Haider, M.Z.; Habib, N.; Rizwan, M.; Perveen, R.; Ali, S.; Nasser Alyemeni, M.; El-Serehy, H.A.; Al-Misned, F.A. α-Tocopherol Foliar Spray and Translocation Mediates Growth, Photosynthetic Pigments, Nutrient Uptake, and Oxidative Defense in Maize (Zea mays L.) under Drought Stress. Agronomy 2020, 10, 1235. https://doi.org/10.3390/agronomy10091235
Ali Q, Tariq Javed M, Haider MZ, Habib N, Rizwan M, Perveen R, Ali S, Nasser Alyemeni M, El-Serehy HA, Al-Misned FA. α-Tocopherol Foliar Spray and Translocation Mediates Growth, Photosynthetic Pigments, Nutrient Uptake, and Oxidative Defense in Maize (Zea mays L.) under Drought Stress. Agronomy. 2020; 10(9):1235. https://doi.org/10.3390/agronomy10091235
Chicago/Turabian StyleAli, Qasim, Muhammad Tariq Javed, Muhammad Zulqurnain Haider, Noman Habib, Muhammad Rizwan, Rashida Perveen, Shafaqat Ali, Mohammed Nasser Alyemeni, Hamed A. El-Serehy, and Fahad A. Al-Misned. 2020. "α-Tocopherol Foliar Spray and Translocation Mediates Growth, Photosynthetic Pigments, Nutrient Uptake, and Oxidative Defense in Maize (Zea mays L.) under Drought Stress" Agronomy 10, no. 9: 1235. https://doi.org/10.3390/agronomy10091235
APA StyleAli, Q., Tariq Javed, M., Haider, M. Z., Habib, N., Rizwan, M., Perveen, R., Ali, S., Nasser Alyemeni, M., El-Serehy, H. A., & Al-Misned, F. A. (2020). α-Tocopherol Foliar Spray and Translocation Mediates Growth, Photosynthetic Pigments, Nutrient Uptake, and Oxidative Defense in Maize (Zea mays L.) under Drought Stress. Agronomy, 10(9), 1235. https://doi.org/10.3390/agronomy10091235