The Diurnal Rhythm of Brassica napus L. Influences Contents of Sulfur-Containing Defense Compounds and Occurrence of Vascular Occlusions during an Infection with Verticillium longisporum

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
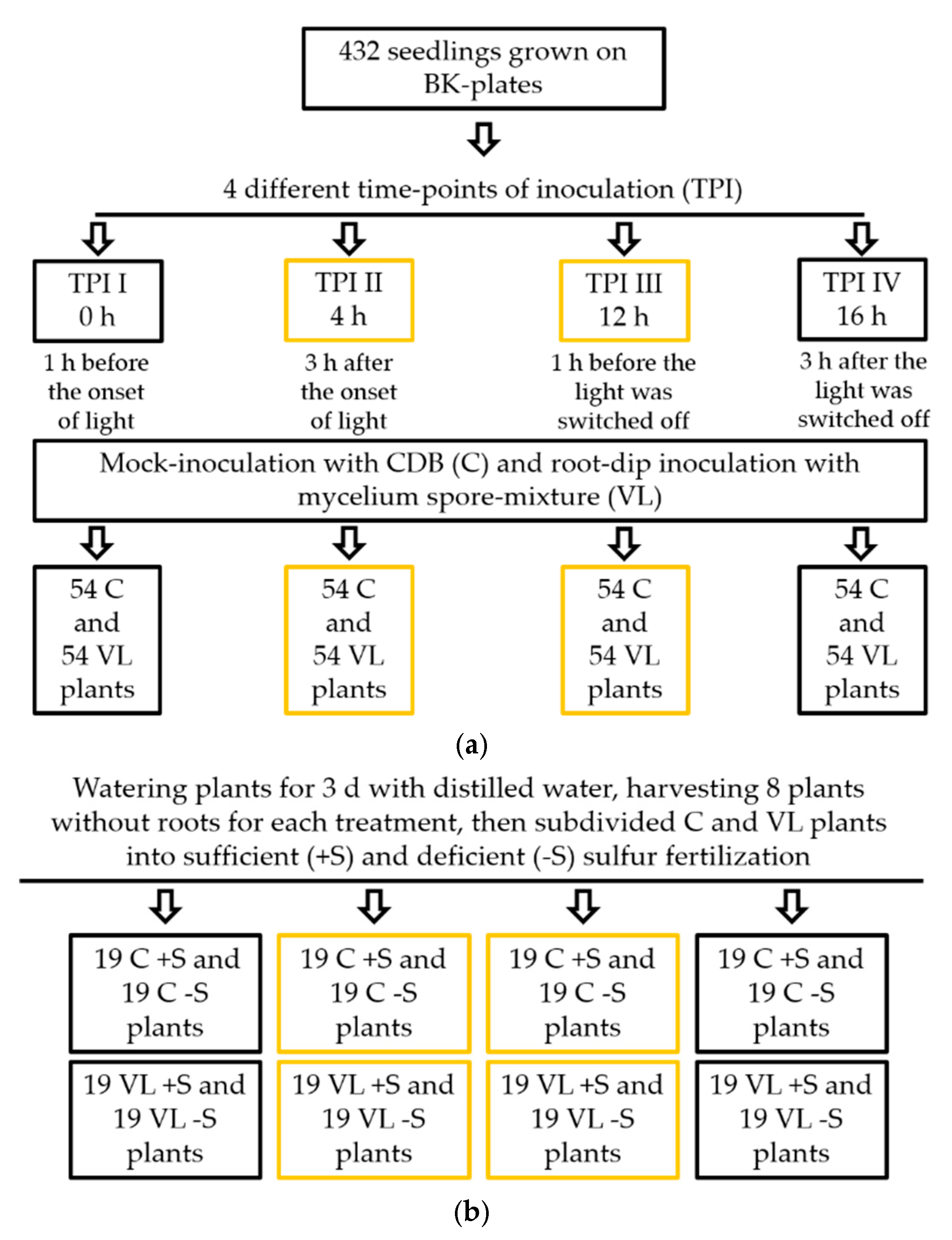
2.1. Plant Material and Growth Conditions
2.2. Verticillium longisporum Material, Growth and Cultivation
2.3. Inoculation Procedure of Brassica napus with Verticillium longisporum
2.4. DNA Extraction
2.5. Real-Time Quantitative PCR
2.6. Elemental Analysis and Analysis of Soluble Thiol Compounds
2.7. Analysis of Glucosinolates
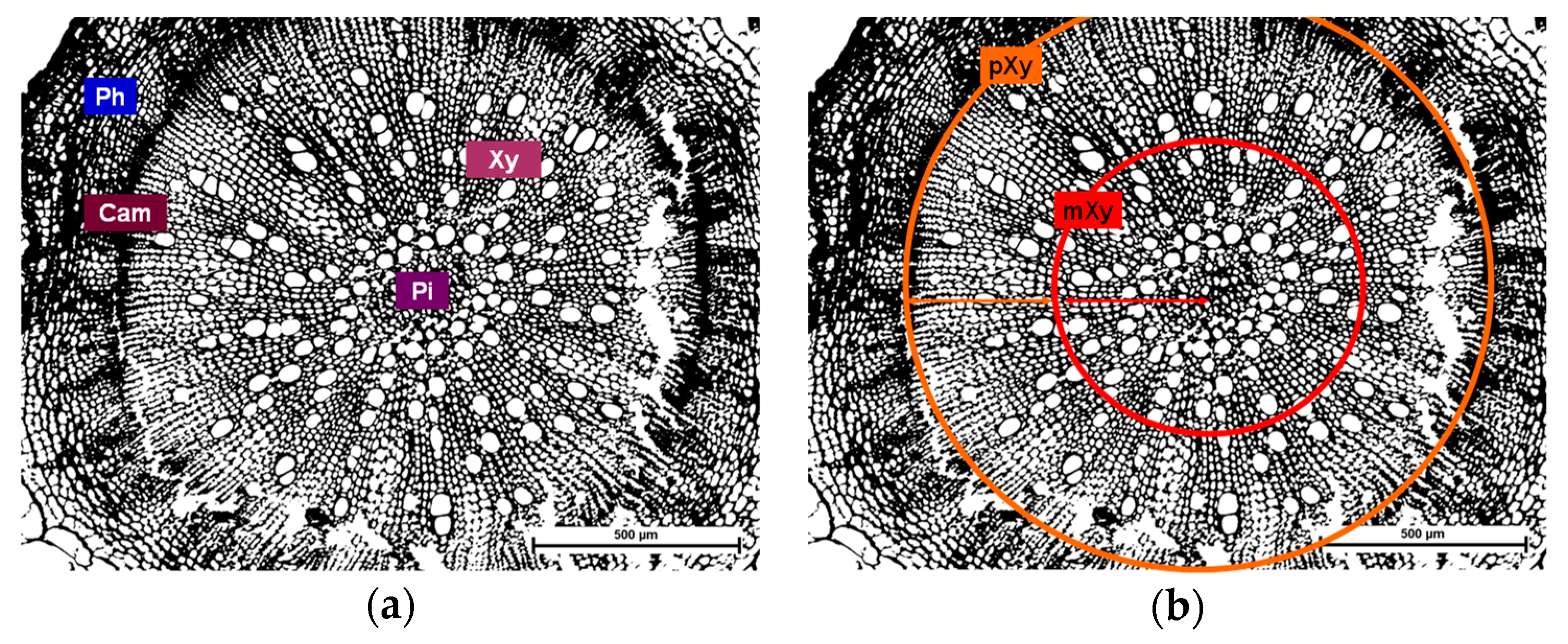
2.8. Histological Analysis
2.9. Statistical Analysis
3. Results
3.1. Detection and Verification of the Infection with V. longisporum in B. napus
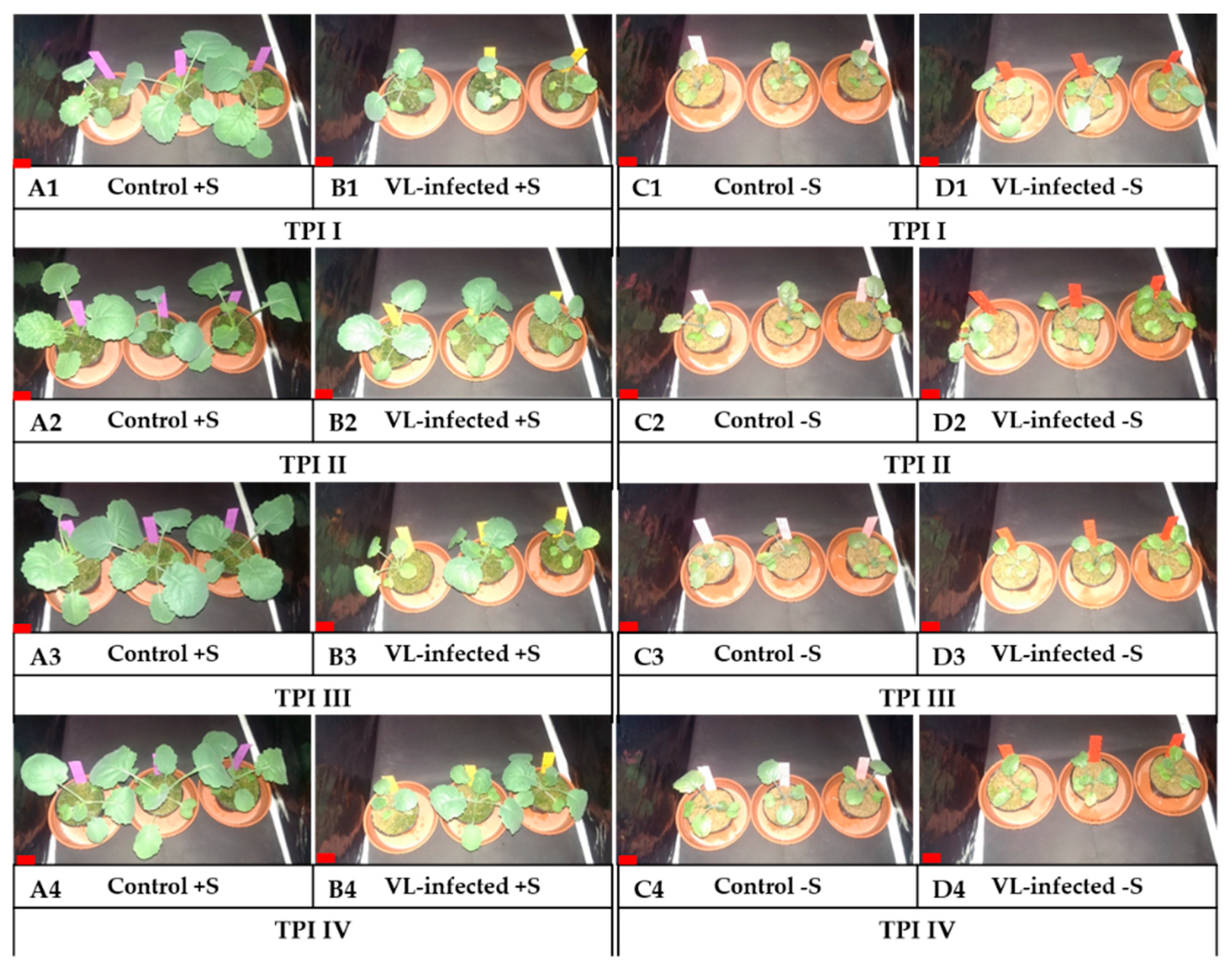
3.2. Stunting of B. napus Plants Infected with the V. longisporum Strain VL43
3.3. Levels of Indolic, Aliphatic and Benzylic GSLs in Mock-Treated and Plants Infected with V. longisporum at Different TPIs and Under Different Sulfur Supplies
3.4. Levels of Cysteine and Glutathione
3.5. Levels of Sulfur, Calcium, Potassium and Iron
3.6. The Influence of Different TPIs and Different Sulfur Supply on the Occurrence of Occlusions in the Xylem of B. napus Infected with V. longisporumSstrain VL43
4. Discussion
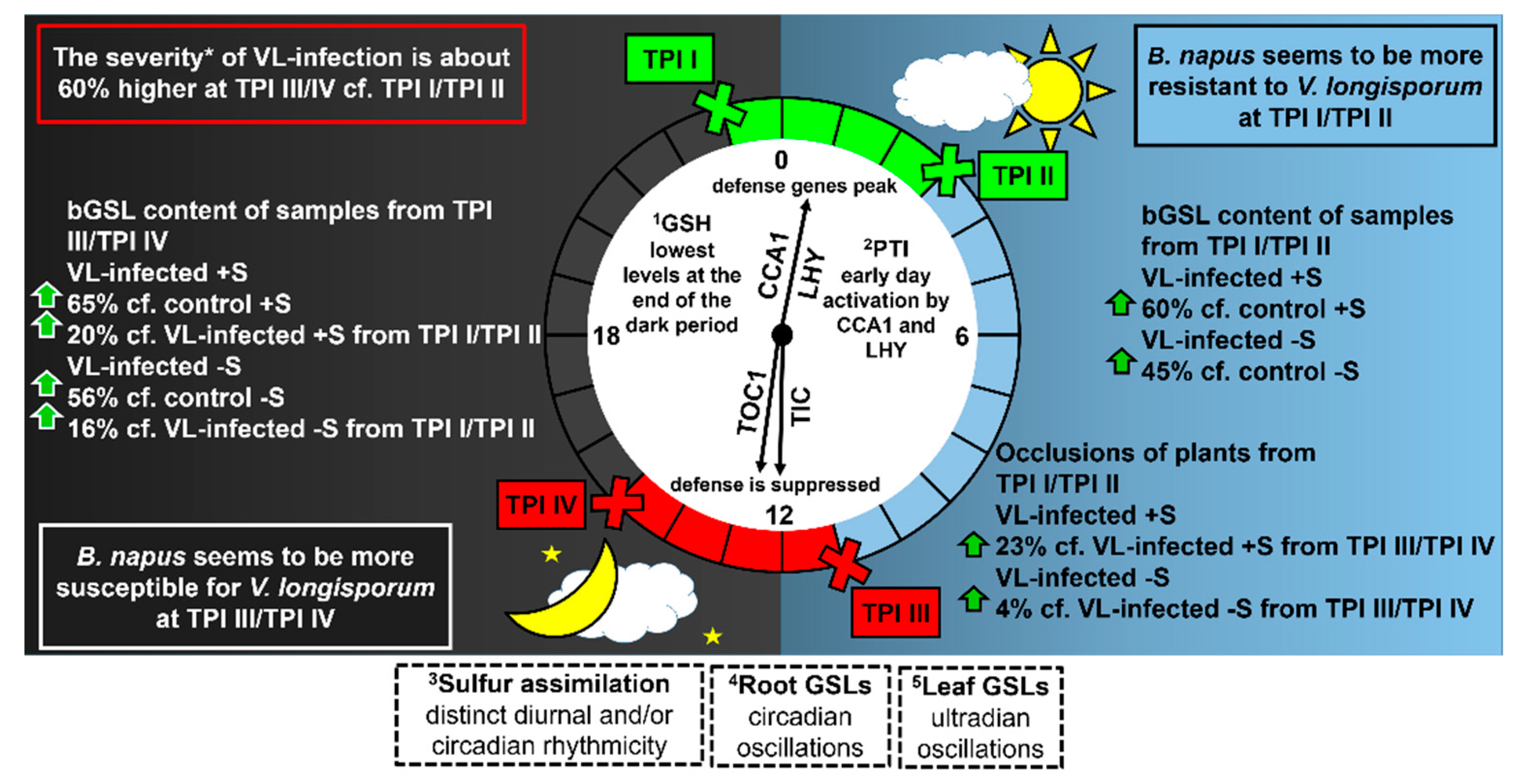
4.1. Higher Sulfur-Containing Compound Levels in Infected Plants–Battle (f)or Survival?
4.2. Occlusions: The Physiochemical Barrier of B. napus against the spread of V. longisporum
4.3. Importance of Diurnal Rhythm in the Defense against V. longisporum
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2005, 144, 31–43. [Google Scholar] [CrossRef]
- Zeilinger, S.; Gupta, V.K.; Dahms, T.E.S.; Silva, R.N.; Singh, H.B.; Upadhyay, R.S.; Gomes, E.V.; Tsui, C.K.-M.; Nayak, S.C. Friends or foes? Emerging insights from fungal interactions with plants. FEMS Microbiol. Rev. 2015, 40, 182–207. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Meena, V.S.; Maurya, B.R.; Meena, S.K.; Meena, R.K.; Kumar, A.; Verma, J.P.; Singh, N.P. Can Bacillus Species Enhance Nutrient Availability in Agricultural Soils? In Bacilli and Agrobiotechnology; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2016; pp. 367–395. [Google Scholar]
- DePotter, J.R.L.; Deketelaere, S.; Inderbitzin, P.; Von Tiedemann, A.; Höfte, M.; Subbarao, K.V.; Wood, T.A.; Thomma, B.P. Verticillium longisporum, the invisible threat to oilseed rape and other brassicaceous plant hosts. Mol. Plant Pathol. 2016, 17, 1004–1016. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Braus-Stromeyer, S.A.; Timpner, C.; Tran, V.T.; Lohaus, G.; Reusche, M.; Knüfer, J.; Teichmann, T.; Von Tiedemann, A.; Braus, G.H. Silencing of Vlaro2 for chorismate synthase revealed that the phytopathogen Verticillium longisporum induces the cross-pathway control in the xylem. Appl. Microbiol. Biotechnol. 2009, 85, 1961–1976. [Google Scholar] [CrossRef] [PubMed]
- Dunker, S.; Keunecke, H.; Steinbach, P.; von Tiedemann, A. Impact of Verticillium longisporum on yield and morphology of winter oilseed rape (Brassica napus) in relation to systemic spread in the plant. J. Phytopathol. 2008, 156, 698–707. [Google Scholar] [CrossRef]
- Heale, J.B. Diversification and speciation in Verticillium-an overview. In Adv. Verticillium Res. Disease Manag.; APS Press: St Paul, MN, USA, 2000; pp. 1–14. [Google Scholar]
- Heale, J.B.; Karapapa, V.K. The Verticillium threat to canada’s major oilseed crop: Canola. Can. J. Plant Pathol. 1999, 21, 1–7. [Google Scholar] [CrossRef]
- Eynck, C.; Koopmann, B.; Karlovsky, P.; Von Tiedemann, A. Internal Resistance in Winter Oilseed Rape Inhibits Systemic Spread of the Vascular Pathogen Verticillium longisporum. Phytopathology 2009, 99, 802–811. [Google Scholar] [CrossRef]
- Booth, E.J.; Walker, K.C. The effect of site and foliar sulfur on oilseed rape: Comparison of sulfur responsive and non-responsive seasons. Phyton 1992, 32, 9–13. [Google Scholar]
- Pedersen, C.A.; Knudsen, L.; Schnug, E. Sulphur Fertilisation. In Nutrients in Ecosystems; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1998; pp. 115–134. [Google Scholar]
- Richards, I.R. Sulphur as a crop nutrient in the United Kingdom. Sulphur Agric. 1990, 14, 8–9. [Google Scholar]
- Schnug, E. Sulphur nutritional status of European crops and consequences for agriculture. Sulphur Agric. 1991, 15, 7–12. [Google Scholar]
- De Kok, L.J.; Westerman, S.; Stuiver, C.E.E.; Stulen, I. Atmospheric H2S as plant sulfur source: Interaction with pedospheric sulfur nutrition–a case study with Brassica oleracea L. In Sulfur Nutrition and Sulfur Assimilation in Higher Plants: Molecular, Biochemical and Physiological Aspects’; Brunold, C., Rennenberg, H., De Kok, L.J., Stulen, I., Davidian, J.C., Eds.; Haupt Verlag: Bern, Switzerland, 2000; pp. 41–55. [Google Scholar]
- Rausch, T.; Wachter, A. Sulfur metabolism: A versatile platform for launching defence operations. Trends Plant Sci. 2005, 10, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Kruse, C.; Jost, R.; Lipschis, M.; Kopp, B.; Hartmann, M.; Hell, R. Sulfur-Enhanced Defence: Effects of Sulfur Metabolism, Nitrogen Supply, and Pathogen Lifestyle. Plant Biol. 2007, 9, 608–619. [Google Scholar] [CrossRef] [PubMed]
- Hornbacher, J.; Rumlow, A.; Pallmann, P.; Turcios, A.E.; Riemenschneider, A.; Papenbrock, J. The Levels of Sulfur-containing Metabolites in Brassica napus are Not Influenced by the Circadian Clock but Diurnally. J. Plant Biol. 2019, 62, 359–373. [Google Scholar] [CrossRef]
- Kerwin, R.E.; Jiménez-Gómez, J.M.; Fulop, D.; Harmer, S.L.; Maloof, J.N.; Kliebenstein, D.J. Network Quantitative Trait Loci Mapping of Circadian Clock Outputs Identifies Metabolic Pathway-to-Clock Linkages in Arabidopsis. Plant Cell 2011, 23, 471–485. [Google Scholar] [CrossRef]
- Andersson, D.; Chakrabarty, R.; Bejai, S.; Zhang, J.; Rask, L.; Meijer, J. Myrosinases from root and leaves of Arabidopsis thaliana have different catalytic properties. Phytochemistry 2009, 70, 1345–1354. [Google Scholar] [CrossRef]
- Mithen, R.; Campos, H. Genetic variation of aliphatic glucosinolates in Arabidopsis thaliana and prospects for map based gene cloning. In Proceedings of the 9th International Symposium on Insect-Plant Relationships, Gwatt, Switzerland, 24–30 June 1995; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1996; pp. 202–205. [Google Scholar]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Hopkins, R.; Van Dam, N.M.; Van Loon, J.J. Role of Glucosinolates in Insect-Plant Relationships and Multitrophic Interactions. Annu. Rev. EÈntomol. 2009, 54, 57–83. [Google Scholar] [CrossRef]
- Mithen, R. Glucosinolates—Biochemistry, genetics and biological activity. Plant Growth Regul. 2001, 34, 91–103. [Google Scholar] [CrossRef]
- Bednarek, P. Sulfur-containing secondary metabolites from Arabidopsis thaliana and other Brassicaceae with function in plant immunity. ChemBioChem 2012, 13, 1846–1859. [Google Scholar] [CrossRef]
- Wittstock, U.; Halkier, B.A. Glucosinolate research in the Arabidopsis era. Trends Plant Sci. 2002, 7, 263–270. [Google Scholar] [CrossRef]
- Bones, A.M.; Rossiter, J.T. The myrosinase-glucosinolate system, its organisation and biochemistry. Physiol. Plant. 1996, 97, 194–208. [Google Scholar] [CrossRef]
- Schnug, E.; Booth, E.; Haneklaus, S.; Walker, K.C. Sulphur supply and stress resistance in oilseed rape. In Proceedings of the 9th International Rapeseed Congress, Cambridge, UK, 4–7 July 1995; Volume 1995, pp. 229–231. [Google Scholar]
- Davidson, R.M.; Goss, R.L. Effects of P, S, N, lime, chlordane, and fungicides on ophiobolus patch disease of turf. Plant Disease Rep. 1972, 56, 565–567. [Google Scholar]
- Roden, L.C.; Ingle, R.A. Lights, Rhythms, Infection: The Role of Light and the Circadian Clock in Determining the Outcome of Plant–Pathogen Interactions. Plant Cell 2009, 21, 2546–2552. [Google Scholar] [CrossRef]
- Chandra-Shekara, A.C.; Gupte, M.; Navarre, D.; Raina, S.; Raina, R.; Klessig, D.; Kachroo, P. Light-dependent hypersensitive response and resistance signaling against Turnip Crinkle Virus in Arabidopsis. Plant J. 2006, 45, 320–334. [Google Scholar] [CrossRef]
- Griebel, T.; Zeier, J. Light Regulation and Daytime Dependency of Inducible Plant Defenses in Arabidopsis: Phytochrome Signaling Controls Systemic Acquired Resistance Rather Than Local Defense. Plant Physiol. 2008, 147, 790–801. [Google Scholar] [CrossRef]
- Lu, H.; McClung, C.R.; Zhang, C. Tick Tock: Circadian Regulation of Plant Innate Immunity. Annu. Rev. Phytopathol. 2017, 55, 287–311. [Google Scholar] [CrossRef]
- Oberpichler, I.; Rosen, R.; Rasouly, A.; Vugman, M.; Ron, E.Z.; Lamparter, T. Light affects motility and infectivity of Agrobacterium tumefaciens. Environ. Microbiol. 2008, 10, 2020–2029. [Google Scholar] [CrossRef]
- Wang, W.; Barnaby, J.Y.; Tada, Y.; Li, H.; Tör, M.; Caldelari, D.; Lee, D.-U.; Fu, X.-D.; Dong, X. Timing of plant immune responses by a central circadian regulator. Nature 2011, 470, 110–114. [Google Scholar] [CrossRef]
- Bhardwaj, V.; Meier, S.; Petersen, L.N.; Ingle, R.A.; Roden, L.C. Defence Responses of Arabidopsis thaliana to Infection by Pseudomonas syringae Are Regulated by the Circadian Clock. PLoS ONE 2011, 6, e26968. [Google Scholar] [CrossRef]
- Ingle, R.A.; Stoker, C.; Stone, W.; Adams, N.; Smith, R.; Grant, M.; Carré, I.A.; Roden, L.C.; Denby, K. Jasmonate signalling drives time-of-day differences in susceptibility of Arabidopsis to the fungal pathogen Botrytis cinerea. Plant J. 2015, 84, 937–948. [Google Scholar] [CrossRef]
- Haydon, M.J.; Roman, A.; Arshad, W. Nutrient homeostasis within the plant circadian network. Front. Plant Sci. 2015, 6, 299. [Google Scholar] [CrossRef] [PubMed]
- Chiasson, D.; Loughlin, P.C.; Mazurkiewicz, D.; Mohammadi-Dehcheshmeh, M.; Fedorova, E.E.; Okamoto, M.; McLean, E.; Glass, A.D.M.; Smith, S.E.; Bisseling, T.; et al. Soybean SAT1 (Symbiotic Ammonium Transporter 1) encodes a bHLH transcription factor involved in nodule growth and NH4+ transport. Proc. Natl. Acad. Sci. USA 2014, 111, 4814–4819. [Google Scholar] [CrossRef] [PubMed]
- Michael, T.P.; McClung, C.R. Enhancer Trapping Reveals Widespread Circadian Clock Transcriptional Control in Arabidopsis1[w]. Plant Physiol. 2003, 132, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Weese, A.; Pallmann, P.; Papenbrock, J.; Riemenschneider, A. Brassica napus L. cultivars show a broad variability in their morphology, physiology and metabolite levels in response to sulfur limitations and to pathogen attack. Front. Plant Sci. 2015, 6, 9. [Google Scholar] [CrossRef]
- Blake-Kalff, M.M.; Harrison, K.R.; Hawkesford, M.J.; Zhao, F.J.; McGrath, S.P. Distribution of Sulfur within Oilseed Rape Leaves in Response to Sulfur Deficiency during Vegetative Growth. Plant Physiol. 1998, 118, 1337–1344. [Google Scholar] [CrossRef]
- Zeise, K.; Von Tiedemann, A. Morphological and Physiological Differentiation among Vegetative Compatibility Groups of Verticillium dahliae in Relation to V. longisporum. J. Phytopathol. 2001, 149, 469–475. [Google Scholar] [CrossRef]
- Rumlow, A.; Keunen, E.; Klein, J.; Pallmann, P.; Riemenschneider, A.; Cuypers, A.; Papenbrock, J. Quantitative Expression Analysis in Brassica napus by Northern Blot Analysis and Reverse Transcription-Quantitative PCR in a Complex Experimental Setting. PLoS ONE 2016, 11, e0163679. [Google Scholar] [CrossRef]
- Eynck, C.; Koopmann, B.; Grunewaldt-Stoecker, G.; Karlovsky, P.; Von Tiedemann, A. Differential interactions of Verticillium longisporum and V. dahliae with Brassica napus detected with molecular and histological techniques. Eur. J. Plant Pathol. 2007, 118, 259–274. [Google Scholar] [CrossRef]
- Riemenschneider, A.; Wegele, R.; Schmidt, A.; Papenbrock, J. Isolation and characterization of a D-cysteine desulfhydrase protein from Arabidopsis thaliana. FEBS J. 2005, 272, 1291–1304. [Google Scholar] [CrossRef]
- Feder, N.E.D.; O’Brien, T.P. Plant microtechnique: Some principles and new methods. Am. J. Bot. 1968, 55, 123–142. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Tinevez, J.Y. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676. [Google Scholar] [CrossRef] [PubMed]
- McCullagh, P.; Nelder, J. Generalized Linear Models, 2rd ed.; Chapman & Hall: London, UK, 1989. [Google Scholar]
- Lenth, R. Emmeans Package: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.3. 5.1. 2019. Available online: http://packages.renjin.org/package/org.renjin.cran/emmeans (accessed on 18 August 2020).
- Kamble, A.; Koopmann, B.; Von Tiedemann, A. Induced resistance to Verticillium longisporum in Brassica napus by β-aminobutyric acid. Plant Pathol. 2012, 62, 552–561. [Google Scholar] [CrossRef]
- Dunker, S.; Keunecke, H.; von Tiedemann, A. Verticillium longisporum in winter oilseed rape-Impact on plant development and yield. IOBC WPRS Bull. 2006, 29, 361. [Google Scholar]
- Lopisso, D.T.; Knüfer, J.; Koopmann, B.; Von Tiedemann, A. The Vascular Pathogen Verticillium longisporum Does Not Affect Water Relations and Plant Responses to Drought Stress of Its Host, Brassica napus. Phytopathology 2017, 107, 444–454. [Google Scholar] [CrossRef]
- Iven, T.; König, S.; Singh, S.; Braus-Stromeyer, S.A.; Bischoff, M.; Tietze, L.F.; Braus, G.H.; Lipka, V.; Feussner, I.; Dröge-Laser, W. Transcriptional Activation and Production of Tryptophan-Derived Secondary Metabolites in Arabidopsis Roots Contributes to the Defense against the Fungal Vascular Pathogen Verticillium longisporum. Mol. Plant 2012, 5, 1389–1402. [Google Scholar] [CrossRef]
- Rygulla, W.; Friedt, W.; Seyis, F.; Lühs, W.; Eynck, C.; Von Tiedemann, A.; Snowdon, R.J. Combination of resistance to Verticillium longisporum from zero erucic acid Brassica oleracea and oilseed Brassica rapa genotypes in resynthesized rapeseed (Brassica napus) lines. Plant Breed. 2007, 126, 596–602. [Google Scholar] [CrossRef]
- Witzel, K.; Hanschen, F.S.; Schreiner, M.; Krumbein, A.; Ruppel, S.; Grosch, R. Verticillium Suppression Is Associated with the Glucosinolate Composition of Arabidopsis thaliana Leaves. PLoS ONE 2013, 8, e71877. [Google Scholar] [CrossRef]
- Timpner, C.; Braus-Stromeyer, S.A.; Tran, V.T.; Braus, G.H. The Cpc1 Regulator of the Cross-Pathway Control of Amino Acid Biosynthesis Is Required for Pathogenicity of the Vascular Pathogen Verticillium longisporum. Mol. Plant Microbe Interact. 2013, 26, 1312–1324. [Google Scholar] [CrossRef]
- Bloem, E.; Riemenschneider, A.; Volker, J.; Papenbrock, J.; Schmidt, A.; Salac, I.; Haneklaus, S.; Schnug, E. Sulphur supply and infection with Pyrenopeziza brassicae influence L-cysteine desulphydrase activity in Brassica napus L. J. Exp. Bot. 2004, 55, 2305–2312. [Google Scholar] [CrossRef]
- Bloem, E.; Haneklaus, S.; Kleinwächter, M.; Paulsen, J.; Schnug, E.; Selmar, D. Stress-induced changes of bioactive compounds in Tropaeolum majus L. Ind. Crop. Prod. 2014, 60, 349–359. [Google Scholar] [CrossRef]
- Künstler, A.; Kátay, G.; Gullner, G.; Király, L. Artificial elevation of glutathione contents in salicylic acid-deficient tobacco (Nicotiana tabacum cv. Xanthi NahG) reduces susceptibility to the powdery mildew pathogen Euoidium longipes. Plant Biol. 2019, 22, 70–80. [Google Scholar] [CrossRef]
- Geu-Flores, F.; Møldrup, M.E.; Böttcher, C.; Olsen, C.E.; Scheel, D.; Halkier, B.A. Cytosolic γ-Glutamyl Peptidases Process Glutathione Conjugates in the Biosynthesis of Glucosinolates and Camalexin in Arabidopsis. Plant Cell 2011, 23, 2456–2469. [Google Scholar] [CrossRef]
- Talboys, P.W. Chemical control of Verticillium wilts. Phytopathol. Mediterr. 1984, 23, 163–175. [Google Scholar]
- Beckman, C.H. Phenolic-storing cells: Keys to programmed cell death and periderm formation in wilt disease resistance and in general defence responses in plants? Physiol. Mol. Plant Pathol. 2000, 57, 101–110. [Google Scholar] [CrossRef]
- Fradin, E.F.; Thomma, B.P. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Covington, M.F.; Harmer, S.L. The Circadian Clock Regulates Auxin Signaling and Responses in Arabidopsis. PLoS Biol. 2007, 5, e222. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.-Y.; Zhou, M.; Yoo, H.; Pruneda-Paz, J.L.; Spivey, N.W.; Kay, S.A.; Dong, X. Spatial and temporal regulation of biosynthesis of the plant immune signal salicylic acid. Proc. Natl. Acad. Sci. USA 2015, 112, 9166–9173. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Song, T.; Zhang, X.; Yuan, H.; Su, L.; Li, W.; Xu, J.; Liu, S.; Chen, L.; Chen, T.; et al. Unconventionally secreted effectors of two filamentous pathogens target plant salicylate biosynthesis. Nat. Commun. 2014, 5, 4686. [Google Scholar] [CrossRef]
- Spoel, S.H.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef]
- Zheng, X.; Koopmann, B.; Von Tiedemann, A. Role of Salicylic Acid and Components of the Phenylpropanoid Pathway in Basal and Cultivar-Related Resistance of Oilseed Rape (Brassica napus) to Verticillium longisporum. Plants 2019, 8, 491. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Heidrich, K.; Sanchez-Villarreal, A.; Parker, J.E.; Davis, S.J. TIME FOR COFFEE Represses Accumulation of the MYC2 Transcription Factor to Provide Time-of-Day Regulation of Jasmonate Signaling in Arabidopsis. Plant Cell 2012, 24, 2470–2482. [Google Scholar] [CrossRef] [PubMed]
- Oliverio, K.A.; Crepy, M.; Martin-Tryon, E.L.; Milich, R.; Harmer, S.L.; Putterill, J.; Yanovsky, M.J.; Casal, J. GIGANTEA Regulates Phytochrome A-Mediated Photomorphogenesis Independently of Its Role in the Circadian Clock. Plant Physiol. 2007, 144, 495–502. [Google Scholar] [CrossRef][Green Version]
- Kopriva, S.; Muheim, R.; Koprivova, A.; Trachsel, N.; Catalano, C.; Suter, M.; Brunold, C. Light regulation of assimilatory sulphate reduction in Arabidopsis thaliana. Plant J. 1999, 20, 37–44. [Google Scholar] [CrossRef]
- Pilgrim, M.L.; Caspar, T.; Quail, P.H.; McClung, C.R. Circadian and light-regulated expression of nitrate reductase in Arabidopsis. Plant Mol. Boil. 1993, 23, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Zeeman, S.C.; Smith, S.M.; Smith, A.M. The diurnal metabolism of leaf starch. Biochem. J. 2006, 401, 13–28. [Google Scholar] [CrossRef]
- Harmer, S.L. The Circadian System in Higher Plants. Annu. Rev. Plant Boil. 2009, 60, 357–377. [Google Scholar] [CrossRef]
- De Moraes, C.M.; Mescher, M.C.; Tumlinson, J.H. Caterpillar-induced nocturnal plant volatiles repel conspecific females. Nature 2001, 410, 577–580. [Google Scholar] [CrossRef]
- Kim, S.-G.; Yon, F.; Gaquerel, E.; Gulati, J.; Baldwin, I.T. Tissue Specific Diurnal Rhythms of Metabolites and Their Regulation during Herbivore Attack in a Native Tobacco, Nicotiana attenuata. PLoS ONE 2011, 6, e26214. [Google Scholar] [CrossRef]
- Karapetyan, S.; Dong, X. Redox and the circadian clock in plant immunity: A balancing act. Free. Radic. Boil. Med. 2017, 119, 56–61. [Google Scholar] [CrossRef]
- Zechmann, B. Diurnal changes of subcellular glutathione content in Arabidopsis thaliana. Biol. Plant. 2017, 61, 791–796. [Google Scholar] [CrossRef]
- Zhang, C.; Xie, Q.; Anderson, R.G.; Ng, G.; Seitz, N.C.; Peterson, T.; McClung, C.R.; McDowell, J.M.; Kong, D.; Kwak, J.M.; et al. Crosstalk between the Circadian Clock and Innate Immunity in Arabidopsis. PLoS Pathog. 2013, 9, e1003370. [Google Scholar] [CrossRef] [PubMed]
- Rosa, E.A.; Rodrigues, P.M.F. The effect of light and temperature on glucosinolate concentration in the leaves and roots of cabbage seedlings. J. Sci. Food Agric. 1998, 78, 208–212. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fertilization | Harvesting | Sample Size of Each Group | Method |
|---|---|---|---|
| Distilled water as required | 3 dpi | Eight plants without root | DNA-extraction qPCR |
| Distilled water as required and Corresponding BK medium (+S/−S) 1: 150 mL each pot weekly | 7 dpi | Eight plants without root | DNA-extraction qPCR Elemental analysis via ICP-OES GSL analysis via HPLC Cysteine and glutathione analysis via HPLC |
| 14 dpi | Eight plants without root Histology: hypocotyl of one plant | DNA-extraction qPCR Elemental analysis via ICP-OES GSL analysis via HPLC Cysteine and glutathione analysis via HPLC Histology: Toluidine blue staining | |
| 21 dpi | Only histology: hypocotyl of 1 plant | Histology: Toluidine blue staining |
| 3 dpi/ng VL DNA g−1 FW | VL without +S/−S Fertilization | VL without +S/−S Fertilization |
| TPI I | 6.17 | 7.05 |
| TPI II | 13.85 | 10.54 |
| TPI III | 17.22 | 19.54 |
| TPI IV | 27.12 | 27.20 |
| 7 dpi/ng VL DNA g−1 FW | VL +S | VL −S |
| TPI I | 13.54 | 12.37 |
| TPI II | 4.03 | 7.68 |
| TPI III | 8.01 | 12.50 |
| TPI IV | 8.15 | 8.27 |
| 14 dpi/ng VL DNA g−1 FW | VL +S | VL −S |
| TPI I | 0.29 | 4.79 |
| TPI II | 1.06 | 5.87 |
| TPI III | 0.51 | 1.82 |
| TPI IV | 0.33 | 5.45 |
| Glucobrassicin [nmol g−1 DW] | |||||||||
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C +S | VL +S | C −S | VL −S |
| TPI I | 195.99 | 304.14 | 136.91 | 328.44 | TPI I | 92.52 ± 4.30 | 158.54 ± 7.70 | 0.41 | 10.76 |
| TPI II | 231.11 | 348.42 | 162.56 | 230.67 | TPI II | 109.19 | 123.30 | 0.06 ± 0.02 | 2.75 ± 0.78 |
| TPI III | 262.87 | 356.19 | 202.76 | 297.80 | TPI III | 97.09 ± 9.92 | 165.79 | 0.32 | 11.91 |
| TPI IV | 351.40 | 456.19 | 213.60 | 267.74 | TPI IV | 106.38 | 139.87 ± 3.86 | 1.11 | 12.31 ± 0.84 |
| Neoglucobrassicin [nmol g−1 DW] | |||||||||
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C +S | VL +S | C −S | VL −S |
| TPI I | 28.83 | 29.87 | 20.62 | 39.53 | TPI I | 20.71 ± 3.58 | 17.77 ± 1.30 | 2.93 | 5.83 |
| TPI II | 37.80 | 40.12 | 28.44 | 47.18 | TPI II | 22.74 | 20.01 | 2.60 ± 0.23 | 6.20 ± 1.45 |
| TPI III | 37.14 | 28.95 | 28.37 | 21.53 | TPI III | 19.05 ± 2.78 | 19.58 | 4.05 | 6.99 |
| TPI IV | 41.08 | 43.18 | 33.60 | 31.21 | TPI IV | 23.32 | 23.62 ± 1.64 | 5.18 | 8.27 ± 0.82 |
| 4-Hydroxyglucobrassicin [nmol g−1 DW] | |||||||||
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C +S | VL +S | C −S | VL −S |
| TPI I | 9.04 | 20.19 | 19.24 | 3.47 | TPI I | 14.94 ± 1.42 | 19.80 ± 5.84 | 0.40 | 0.19 |
| TPI II | 9.97 | 22.83 | 18.44 | 0.96 | TPI II | 15.00 | 6.74 | 0.29 ± 0.02 | 0.22 ± 0.03 |
| TPI III | 4.93 | 20.20 | 2.08 | 5.30 | TPI III | 14.76 ± 2.87 | 4.00 | 0.27 | 0.32 |
| TPI IV | 16.24 | 10.77 | 7.35 | 1.42 | TPI IV | 25.93 | 9.16 ± 4.34 | 0.29 | 0.32 ± 0.04 |
| Progoitrin [nmol g−1 DW] | |||||||||
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C +S | VL +S | C −S | VL −S |
| TPI I | 93.98 | 171.34 | 68.43 | 210.08 | TPI I | 58.07 ± 1.60 | 104.17 ± 9.41 | 0.75 | 0.97 |
| TPI II | 89.62 | 219.13 | 111.49 | 72.20 | TPI II | 95.27 | 97.67 | 1.43 ± 0.43 | 0.90 ± 0.25 |
| TPI III | 61.33 | 169.01 | 46.83 | 109.53 | TPI III | 105.96 ± 12.33 | 108.96 | 0.48 | 0.73 |
| TPI IV | 170.73 | 241.14 | 103.43 | 113.41 | TPI IV | 95.63 | 107.59 ± 8.89 | 0.89 | 1.04 ± 0.05 |
| Glucoalyssin [nmol g−1 DW] | |||||||||
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C +S | VL +S | C −S | VL −S |
| TPI I | 69.56 | 99.80 | 30.68 | 119.98 | TPI I | 54.31 ± 1.73 | 95.48 ± 11.29 | 4.74 | 0.65 |
| TPI II | 54.13 | 183.47 | 56.26 | 47.09 | TPI II | 84.88 | 70.56 | 6.51 ± 0.66 | 2.98 ± 0.33 |
| TPI III | 29.23 | 116.93 | 18.61 | 42.17 | TPI III | 91.62 ± 10.66 | 79.63 | 2.91 | 2.95 |
| TPI IV | 115.74 | 203.69 | 50.57 | 39.80 | TPI IV | 108.33 | 87.88 ± 11.79 | 3.72 | 3.28 ± 0.36 |
| Glucoraphanin [nmol g−1 DW] | |||||||||
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C +S | VL +S | C −S | VL −S |
| TPI I | 14.39 | 14.35 | 2.69 | 9.09 | TPI I | 9.93 ± 0.34 | 14.83 ± 1.61 | 2.15 | 3.91 |
| TPI II | 3.58 | 25.61 | 6.82 | 2.11 | TPI II | 13.25 | 8.23 | 2.91 ± 0.70 | 3.54 ± 0.19 |
| TPI III | 1.56 | 17.26 | 1.54 | 2.30 | TPI III | 11.81 ± 2.10 | 8.21 | 0.88 | 6.72 |
| TPI IV | 13.40 | 27.65 | 4.44 | 4.16 | TPI IV | 17.89 | 8.52 ± 2.38 | 2.00 | 7.54 ± 1.14 |
| Glucobrassicanapin [nmol g−1 DW] | |||||||||
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C +S | VL +S | C −S | VL −S |
| TPI I | 23.89 | 45.03 | 13.68 | 65.71 | TPI I | 27.14 ± 1.30 | 43.01 ± 3.90 | 0.50 | 0.50 |
| TPI II | 42.35 | 54.84 | 26.62 | 20.18 | TPI II | 45.36 | 61.28 | 0.30 ± 0.03 | 0.32 ± 0.27 |
| TPI III | 20.54 | 37.22 | 15.41 | 23.31 | TPI III | 56.59 ± 6.94 | 66.76 | 0.44 | 0.21 |
| TPI IV | 43.87 | 52.53 | 30.01 | 13.38 | TPI IV | 46.39 | 62.48 ± 3.26 | 0.60 | 0.33 ± 0.13 |
| Gluconapin [nmol g−1 DW] | |||||||||
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C +S | VL +S | C −S | VL −S |
| TPI I | 35.33 | 70.21 | 11.30 | 39.84 | TPI I | 22.13 ± 0.93 | 38.01 ± 2.25 | 1.47 | 0.77 |
| TPI II | 28.10 | 77.65 | 24.91 | 8.63 | TPI II | 39.35 | 28.64 | 1.60 ± 0.38 | 0.34 ± 0.09 |
| TPI III | 11.18 | 55.26 | 8.48 | 11.70 | TPI III | 55.56 ± 7.99 | 32.10 | 1.95 | 0.35 |
| TPI IV | 70.01 | 77.80 | 16.69 | 12.60 | TPI IV | 45.30 | 29.42 ± 2.94 | 2.17 | 0.37 ± 0.09 |
| Gluconasturtiin [nmol g−1 DW] | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C +S | VL +S | C −S | VL −S |
| TPI I | 98.71 | 215.12 | 92.05 | 171.25 | TPI I | 52.05 ± 1.43 | 125.92 ± 8.14 | 10.40 | 55.84 |
| TPI II | 69.16 | 190.75 | 107.36 | 183.82 | TPI II | 44.07 | 86.90 | 5.69 ± 0.09 | 35.56 ± 6.55 |
| TPI III | 55.98 | 240.52 | 84.85 | 191.47 | TPI III | 51.71 ± 4.10 | 142.95 | 6.92 | 50.17 |
| TPI IV | 123.95 | 263.75 | 101.10 | 230.38 | TPI IV | 59.60 | 109.09 ± 2.78 | 10.22 | 45.62 ± 1.56 |
| Cysteine [nmol g−1 FW] | |||||||||
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C −S | VL +S | C −S | VL −S |
| TPI I | 27.41 | 28.03 | 13.80 | 6.57 | TPI I | 17.73 ± 1.42 | 19.66 ± 1.58 | 2.95 ± 0.52 | 4.60 ± 0.30 |
| TPI II | 20.57 | 28.43 | 8.62 | 16.29 | TPI II | 22.07 ± 1.30 | 21.42 ± 1.33 | 1.93 ± 0.20 | 1.99 ± 0.60 |
| TPI III | 18.57 | 24.66 | 13.70 | 15.43 | TPI III | 23.35 ± 0.91 | 21.72 ± 2.05 | 1.98 ± 0.36 | 3.41 ± 0.17 |
| TPI IV | 28.51 | 30.26 | 11.85 | 12.00 | TPI IV | 20.20 ± 1.50 | 18.77 ± 1.97 | 1.79 ± 0.88 | 4.10 ± 0.26 |
| GSH [nmol g−1 FW] | |||||||||
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C +S | VL +S | C −S | VL −S |
| TPI I | 559.17 | 420.96 | 278.54 | 568.68 | TPI I | 671.79 ± 26.98 | 707.66 ± 22.27 | 66.21 ± 4.98 | 136.38 ± 13.05 |
| TPI II | 646.72 | 629.21 | 276.83 | 464.82 | TPI II | 690.45 ± 13.15 | 750.78 ± 20.13 | 68.52 ± 9.78 | 63.00 ± 3.03 |
| TPI III | 519.29 | 588.02 | 312.53 | 348.69 | TPI III | 630.34 ± 47.26 | 720.24 ± 37.57 | 65.16 ± 1.73 | 84.59 ± 2.99 |
| TPI IV | 522.86 | 544.11 | 253.71 | 379.52 | TPI IV | 705.57 ± 31.60 | 725.97 ± 12.47 | 58.99 ± 2.20 | 98.21 ± 6.30 |
| Sulfur [mg g−1 DW] | |||||||||
| 7 dpi | C +S | VL +S | C −S | VL −S | 14 dpi | C +S | VL +S | C −S | VL −S |
| TPI I | 8.63 ± 0.02 | 7.74 ± 0.02 | 1.33 ± 0.01 | 3.12 ± 0.02 | TPI I | 6.32 ± 0.04 | 5.29 ± 0.02 | 0.73 ± 0.00 | 1.02 ± 0.00 |
| TPI II | 10.11 ± 0.06 | 9.52 ± 0.06 | 2.09 ± 0.01 | 2.58 ± 0.02 | TPI II | 6.91 ± 0.00 | 6.51 ± 0.01 | 0.71 ± 0.01 | 1.19 ± 0.00 |
| TPI III | 10.72 ± 0.00 | 8.62 ± 0.02 | 2.13 ± 0.00 | 3.01 ± 0.00 | TPI III | 5.31 ± 0.02 | 6.71 ± 0.05 | 0.57 ± 0.00 | 0.61 ± 0.00 |
| TPI IV | 8.70 ± 0.01 | 7.95 ± 0.04 | 2.07 ± 0.01 | 2.15 ± 0.01 | TPI IV | 5.61 ± 0.02 | 6.00 ± 0.03 | 0.63 ± 0.00 | 0.72 ± 0.00 |
| Occurrence of Occlusions in the Mid Area of the Xylem [%] | |||||
| 14 dpi | VL +S | VL −S | 21 dpi | VL +S | VL −S |
| TPI I | 34.60 ± 0.52 | 7.75 ± 0.50 | TPI I | 38.17 ± 0.64 | 34.42 ± 0.38 |
| TPI II | 21.42 ± 0.32 | 9.33 ± 0.52 | TPI II | 33.89 ± 0.50 | 30.44 ± 0.61 |
| TPI III | 8.18 ± 0.17 | 3.56 ± 0.30 | TPI III | 29.60 ± 0.32 | 20.11 ± 0.39 |
| TPI IV | 3.29 ± 0.24 | 5.13 ± 0.67 | TPI IV | 27.99 ± 0.20 | 8.90 ± 0.30 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rupp, S.I.; Hornbacher, J.; Horst-Niessen, I.; Schaarschmidt, F.; Riemenschneider, A.; Papenbrock, J. The Diurnal Rhythm of Brassica napus L. Influences Contents of Sulfur-Containing Defense Compounds and Occurrence of Vascular Occlusions during an Infection with Verticillium longisporum. Agronomy 2020, 10, 1227. https://doi.org/10.3390/agronomy10091227
Rupp SI, Hornbacher J, Horst-Niessen I, Schaarschmidt F, Riemenschneider A, Papenbrock J. The Diurnal Rhythm of Brassica napus L. Influences Contents of Sulfur-Containing Defense Compounds and Occurrence of Vascular Occlusions during an Infection with Verticillium longisporum. Agronomy. 2020; 10(9):1227. https://doi.org/10.3390/agronomy10091227
Chicago/Turabian StyleRupp, Sofia Isabell, Johann Hornbacher, Ina Horst-Niessen, Frank Schaarschmidt, Anja Riemenschneider, and Jutta Papenbrock. 2020. "The Diurnal Rhythm of Brassica napus L. Influences Contents of Sulfur-Containing Defense Compounds and Occurrence of Vascular Occlusions during an Infection with Verticillium longisporum" Agronomy 10, no. 9: 1227. https://doi.org/10.3390/agronomy10091227
APA StyleRupp, S. I., Hornbacher, J., Horst-Niessen, I., Schaarschmidt, F., Riemenschneider, A., & Papenbrock, J. (2020). The Diurnal Rhythm of Brassica napus L. Influences Contents of Sulfur-Containing Defense Compounds and Occurrence of Vascular Occlusions during an Infection with Verticillium longisporum. Agronomy, 10(9), 1227. https://doi.org/10.3390/agronomy10091227