Supporting Beneficial Insects for Agricultural Sustainability: The Role of Livestock-Integrated Organic and Cover Cropping to Enhance Ground Beetle (Carabidae) Communities



Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description and Cropping History
2.2. Experimental Design
2.3. Ground Beetle Community Sampling
2.4. Data Analysis
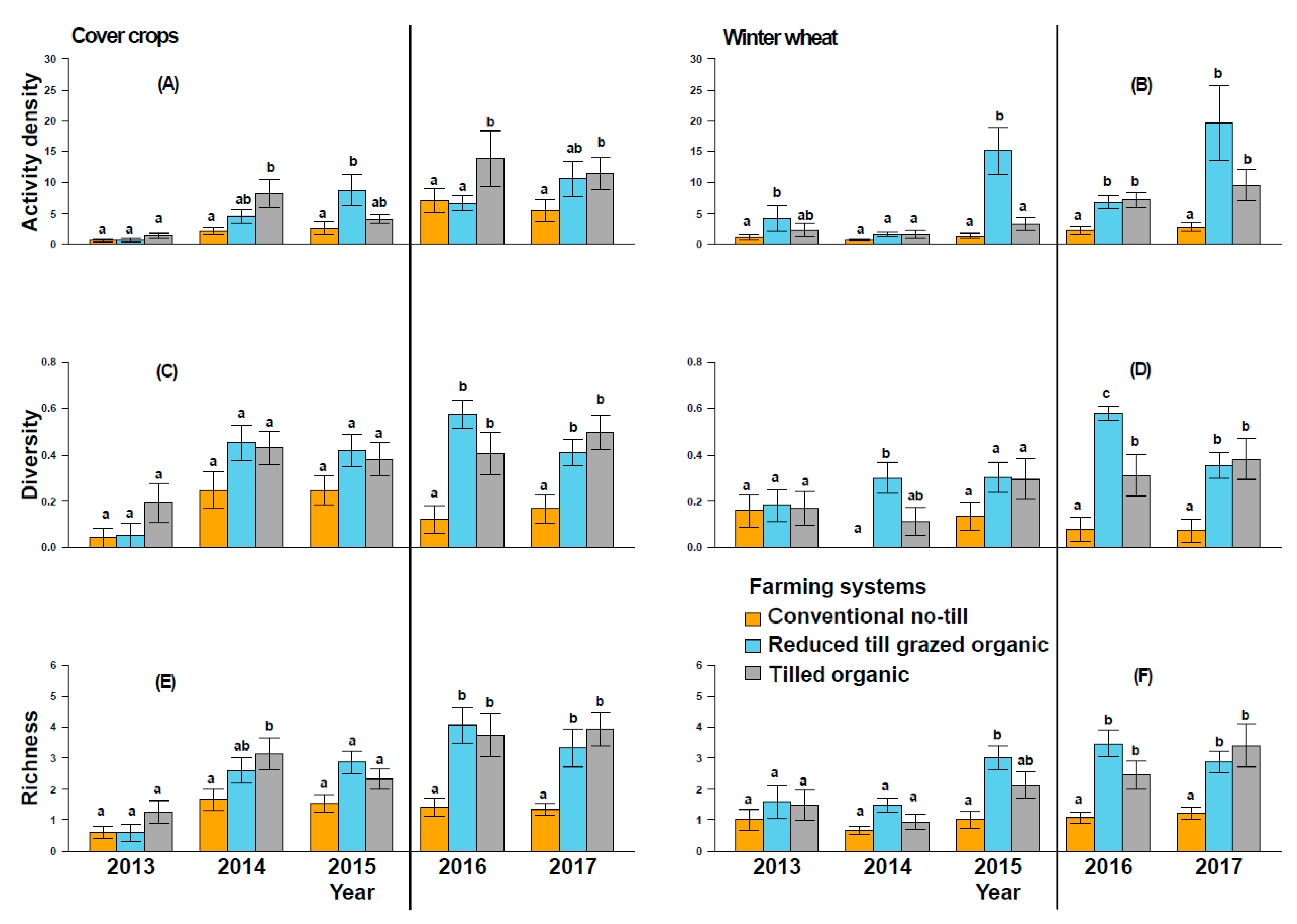
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hole, D.G.; Perkins, A.J.; Wilson, J.D.; Alexander, I.H.; Grice, P.V.; Evans, A.D. Does organic farming benefit biodiversity? Biol. Conserv. 2005, 122, 113–130. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, S.; Adhikari, A.; Weaver, D.K.; Bekkerman, A.; Menalled, F.D. Impacts of agricultural management systems on biodiversity and ecosystem services in highly simplified dryland landscapes. Sustainability 2019, 11, 3223. [Google Scholar] [CrossRef]
- Kremen, C.; Iles, A.; Bacon, C. Diversified farming systems: An agroecological, systems-based alternative to modern industrial agriculutre. Ecol. Soc. 2012, 17, 44. [Google Scholar] [CrossRef]
- Tonitto, C.; David, M.B.; Drinkwater, L.E. Replacing bare fallows with cover crops in fertilizer-intensive cropping systems: A meta-analysis of crop yield and N dynamics. Agric. Ecosyst. Environ. 2006, 112, 58–72. [Google Scholar] [CrossRef]
- Sheaffer, C.C.; Seguin, P. Forage legumes for sustainable cropping systems. J. Crop Prod. 2003, 8, 187–216. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C.; Bailey, B.A. Role of cover crops in improving soil and row crop productivity. Commun. Soil Sci. Plant Anal. 2005, 36, 2733–2757. [Google Scholar] [CrossRef]
- Gardner, J.C.; Faulkner, D.B. Use of cover crops with integrated crop-livestock production systems. In Cover Crops for Clean Water; Hargrove, W.L., Ed.; Soil and Water Conservation Society: Ankeny, IA, USA, 1991; pp. 185–191. [Google Scholar]
- Snapp, S.S.; Swinton, S.M.; Labarta, R.; Mutch, D.; Black, J.R.; Leep, R.; Nyiraneza, J.; O’Neil, K. Evaluating cover crops for benefits, costs and performance within cropping system niches. Agron. J. 2005, 97, 322–332. [Google Scholar]
- Miller, P.R.; Lighthiser, E.J.; Jones, C.A.; Holmes, J.A.; Rick, T.L.; Wraith, J.M. Pea green manure management affects organic winter wheat yield and quality in semiarid Montana. Can. J. Plant Sci. 2011, 91, 497–508. [Google Scholar] [CrossRef]
- Thiessen Martens, J.; Entz, M. Integrating green manure and grazing systems: A review. Can. J. Plant Sci. 2011, 91, 811–824. [Google Scholar] [CrossRef]
- Egan, J.F.; Bohnenblust, E.; Goslee, S.; Mortensen, D.; Tooker, J. Herbicide drift can affect plant and arthropod communities. Agric. Ecosyst. Environ. 2014, 185, 77–87. [Google Scholar] [CrossRef]
- Olszyk, D.; Pfleeger, T.; Lee, E.H.; Plocher, M. Glyphosate and dicamba herbicide tank mixture effects on native plant and non-genetically engineered soybean seedlings. Ecotoxicology 2015, 24, 1014–1027. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Tillage effects on soil degradation, soil resilience, soil quality, and sustainability. Soil Tillage Res. 1993, 27, 1–8. [Google Scholar] [CrossRef]
- Conant, R.T.; Easter, M.; Paustian, K.; Swan, A.; Williams, S. Impacts of periodic tillage on soil C stocks: A synthesis. Soil Tillage Res. 2007, 95, 1–10. [Google Scholar] [CrossRef]
- Miller, Z.J.; Menalled, F.D.; Sainju, U.M.; Lenssen, A.W.; Hatfield, P.G. Integrating sheep grazing into cereal-based crop rotations: Spring wheat yields and weed communities. Agron. J. 2015, 107, 104–112. [Google Scholar] [CrossRef]
- McKenzie, S.C.; Goosey, H.B.; O’Neill, K.M.; Menalled, F.D. Impact of integrated sheep grazing for cover crop termination on weed and ground beetle (Coleoptera:Carabidae) communities. Agric. Ecosyst. Environ. 2016, 218, 141–149. [Google Scholar] [CrossRef]
- Lemaire, G.; Franzluebbers, A.; de Faccio Carvalho, P.C.; Dedieu, B. Integrated crop-livestock systems: Strategies to achieve synergy between agricultural production and environmental quality. Agric. Ecosyst. Environ. 2014, 190, 4–8. [Google Scholar] [CrossRef]
- Fisher, J.; Tozer, P.; Abrecht, D. Livestock in no-till cropping systems—A story of trade-offs. Anim. Prod. Sci. 2012, 52, 197–214. [Google Scholar] [CrossRef]
- Funk, J.L.; Cleland, E.E.; Suding, K.N.; Zavaleta, E.S. Restoration through reassembly: Plant traits and invasion resistance. Trends Ecol. Evol. 2008, 23, 695–703. [Google Scholar] [CrossRef]
- Larochelle, A. The food of carabid beetles (Coleoptera: Carabidae, including Cicindelidae). In Fabreries Supplement; Association des entomologistes amateurs du Québec: Varennes, France, 1990; Volume 5, pp. 1–132. [Google Scholar]
- Sunderland, K.D. The Diet of some predatory arthropods in cereal crops. J. Appl. Ecol. 1975, 12, 507. [Google Scholar] [CrossRef]
- Gaines, H.R.; Gratton, C. Seed predation increases with ground beetle diversity in a Wisconsin (USA) potato agroecosystem. Agric. Ecosyst. Environ. 2010, 137, 329–336. [Google Scholar] [CrossRef]
- Honek, A.; Martinkova, Z.; Jarosik, V. Ground beetles (carabidae) as seed predators. EJE 2013, 100, 531–544. [Google Scholar] [CrossRef]
- Barbosa, P. Agroecosystems and conservation biological control. In Conservation Biological Control; Elsevier: Amsterdam, The Netherlands, 1998; pp. 39–54. [Google Scholar]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef] [PubMed]
- Johnston, A.M.; Tanaka, D.L.; Miller, P.R.; Brandt, S.A.; Nielsen, D.C.; Lafond, G.P.; Riveland, N.R. Oilseed crops for semiarid cropping systems in the Northern Great Plains. Agron. J. 2002, 94, 231–240. [Google Scholar] [CrossRef]
- Jonason, D.; Andersson, G.K.S.; Öckinger, E.; Rundlöf, M.; Smith, H.G.; Bengtsson, J. Assessing the effect of the time since transition to organic farming on plants and butterflies. J. Appl. Ecol. 2011, 48, 543–550. [Google Scholar] [CrossRef]
- Jabbour, R.; Pisani-Gareau, T.; Smith, R.G.; Mullen, C.; Barbercheck, M. Cover crop and tillage intensities alter ground-dwelling arthropod communities during the transition to organic production. Renew. Agric. Food Syst. 2016, 31, 361–374. [Google Scholar] [CrossRef]
- Burgio, G.; Campanelli, G.; Leteo, F.; Ramilli, F.; Depalo, L.; Fabbri, R.; Sgolastra, F. Ecological sustainability of an organic four-year vegetable rotation system: Carabids and other soil arthropods as bioindicators. Agroecol. Sustain. Food Syst. 2015, 39, 295–316. [Google Scholar] [CrossRef]
- Sommaggio, D.; Peretti, E.; Burgio, G. The effect of cover plants management on soil invertebrate fauna in vineyard in Northern Italy. BioControl 2018, 63, 795–806. [Google Scholar] [CrossRef]
- Sainju, U.M.; Lenssen, A.W.; Goosey, H.B.; Snyder, E.; Hatfield, P.G. Sheep grazing in a wheat–fallow system affects dryland soil properties and grain yield. Soil Sci. Soc. Am. J. 2011, 75, 1789. [Google Scholar] [CrossRef]
- Barsotti, J.L.; Sainju, U.M.; Lenssen, A.W.; Montagne, C.; Hatfield, P.G. Crop yields and soil organic matter responses to sheep grazing in US Northern Great Plains. Soil Tillage Res. 2013, 134, 133–141. [Google Scholar] [CrossRef]
- Brown, G.R.; Matthews, I.M. A review of extensive variation in the design of pitfall traps and a proposal for a standard pitfall trap design for monitoring ground-active arthropod biodiversity. Ecol. Evol. 2016, 6, 3953–3964. [Google Scholar] [CrossRef] [PubMed]
- Lindroth, C.H. The ground beetles (Carabidae, excl. Cicindelinae) of Canada and Alaska, Part 1–6; Entomologiska Siillskapet: Lund, Sweden, 1969; pp. 1–1192. [Google Scholar]
- Bousquet, Y. Catalogue of Geadephaga (Coleoptera, Adephaga) of America, North of Mexico. Zookeys 2012, 245, 1–1722. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [PubMed]
- Pielou, E.C. Species-diversity and pattern-diversity in the study of ecological succession. J. Theor. Biol. 1966, 10, 370–383. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution and measurement of specis diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Package “Emmeans”:Estimated Marginal Means, aka Least-Squares Means. 2019. Available online: https://cran.r-project.org/web/packages/emmeans/emmeans.pdf (accessed on 10 January 2020).
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B.; Urban, D.L. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002; Volume 3–12, pp. 1–300. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- R Development Core Team. The R Manuals. 2016. Available online: https://cran.r-project.org/manuals.html (accessed on 20 May 2020).
- Oksanen, J. CRAN—Package Vegan. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 10 January 2020).
- Goosey, H.B.; Mckenzie, S.C.; Rolston, M.G.; O’Neill, K.M.; Menalled, F.D. Impacts of contrasting alfalfa production systems on the drivers of carabid beetle (Coleoptera: Carabidae) community dynamics. Environ. Entomol. 2015, 44, 1052–1064. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, S.; Menalled, F.D. Impacts of dryland farm management systems on weeds and ground beetles (Carabidae) in the Northern Great Plains. Sustainability 2018, 10, 2146. [Google Scholar] [CrossRef]
- Djoudi, E.A.; Plantegenest, M.; Aviron, S.; Pétillon, J. Local vs. landscape characteristics differentially shape emerging and circulating assemblages of carabid beetles in agroecosystems. Agric. Ecosyst. Environ. 2019, 270–271, 149–158. [Google Scholar] [CrossRef]
- Menalled, F.D.; Smith, R.G.; Dauer, J.T.; Fox, T.B. Impact of agricultural management on carabid communities and weed seed predation. Agric. Ecosyst. Environ. 2007, 118, 49–54. [Google Scholar] [CrossRef]
- Lalonde, O.; Légèe, A.; Stevenson, F.C.; Roy, M.; Vanasse, A. Carabid beetle communities after 18 years of conservation tillage and crop rotation in a cool humid climate. Can. Entomol. 2012, 144, 645–657. [Google Scholar] [CrossRef]
- Lichtenberg, E.M.; Kennedy, C.M.; Kremen, C.; Batáry, P.; Berendse, F.; Bommarco, R.; Bosque-Pérez, N.A.; Carvalheiro, L.G.; Snyder, W.E.; Williams, N.M.; et al. A global synthesis of the effects of diversified farming systems on arthropod diversity within fields and across agricultural landscapes. Glob. Chang. Biol. 2017, 23, 4946–4957. [Google Scholar] [CrossRef]
- Rivers, A.; Mullen, C.; Wallace, J.; Barbercheck, M. Cover crop-based reduced tillage system influences Carabidae (Coleoptera) activity, diversity and trophic group during transition to organic production. Renew. Agric. Food Syst. 2017, 32, 538–551. [Google Scholar] [CrossRef]
- Rivers, A.; Voortman, C.; Barbercheck, M. Cover crops support arthropod predator activity with variable effects on crop damage during transition to organic management. Biol. Control 2020, 151, 104377. [Google Scholar] [CrossRef]
- Depalo, L.; Burgio, G.; Magagnoli, S.; Sommaggio, D.; Montemurro, F.; Canali, S.; Masetti, A. Influence of Cover Crop Termination on Ground Dwelling Arthropods in Organic Vegetable Systems. Insects 2020, 11, 445. [Google Scholar] [CrossRef] [PubMed]
- Crowder, D.W.; Northfield, T.D.; Strand, M.R.; Snyder, W.E. Organic agriculture promotes evenness and natural pest control. Nature 2010, 466, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Snyder, W.E. Give predators a complement: Conserving natural enemy biodiversity to improve biocontrol. Biol. Control 2019, 135, 73–82. [Google Scholar] [CrossRef]
- Menalled, F.D.; Lee, J.C.; Landis, D.A. Manipulating carabid beetle abundance alters prey removal rates in corn fields. BioControl 1999, 43, 441–456. [Google Scholar] [CrossRef]
- Frei, B.; Guenay, Y.; Bohan, D.A.; Traugott, M.; Wallinger, C. Molecular analysis indicates high levels of carabid weed seed consumption in cereal fields across Central Europe. J. Pest Sci. 2019, 92, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Carbonne, B.; Bohan, D.A.; Petit, S. Key carabid species drive spring weed seed predation of Viola arvensis. Biol. Control 2020, 141, 104148. [Google Scholar] [CrossRef]
- Kulkarni, S.S.; Dosdall, L.M.; Spence, J.R.; Willenborg, C.J. Depth of seed burial and gender influence weed seed predation by three species of ground beetle (Coleoptera: Carabidae). Weed Sci. 2015, 63, 910–915. [Google Scholar] [CrossRef]
- Petit, S.; Trichard, A.; Biju-Duval, L.; McLaughlin, B.; Bohan, D.A. Interactions between conservation agricultural practice and landscape composition promote weed seed predation by invertebrates. Agric. Ecosyst. Environ. 2017, 240, 45–53. [Google Scholar] [CrossRef]
- Garratt, M.P.D.; Wright, D.J.; Leather, S.R. The effects of farming system and fertilisers on pests and natural enemies: A synthesis of current research. Agric. Ecosyst. Environ. 2011, 141, 261–270. [Google Scholar] [CrossRef]
- Adhikari, S.; Seipel, T.; Menalled, F.D.; Weaver, D.K. Farming system and wheat cultivar affect infestation of and parasitism on Cephus cinctus in the Northern Great Plains. Pest Manag. Sci. 2018, 74, 2480–2487. [Google Scholar] [CrossRef]
- Adhikari, S. Adopting Diversified Organic Farming to Increase Ecosystem Services|Organic Farmer. Available online: http://organicfarmermag.com/2019/09/adopting-diversified-organic-farming-to-increase-ecosystem-services/ (accessed on 20 May 2020).
- Hatten, T.D.; Bosque-Pérez, N.A.; Labonte, J.R.; Guy, S.O.; Eigenbrode, S.D. Effects of tillage on the activity density and biological diversity of carabid beetles in spring and winter crops. Environ. Entomol. 2007, 36, 356–368. [Google Scholar] [CrossRef]
- Eyre, M.D.; Luff, M.L.; Leifert, C. Crop, field boundary, productivity and disturbance influences on ground beetles (Coleoptera, Carabidae) in the agroecosystem. Agric. Ecosyst. Environ. 2013, 165, 60–67. [Google Scholar] [CrossRef]
- Adhikari, S.; Burkle, L.A.; O’Neill, K.M.; Weaver, D.K.; Menalled, F.D. Dryland organic farming increases floral resources and bee colony success in highly simplified agricultural landscapes. Agric. Ecosyst. Environ. 2019, 270–271, 9–18. [Google Scholar] [CrossRef]
- Adhikari, S.; Burkle, L.A.; O’Neill, K.M.; Weaver, D.K.; Delphia, C.M.; Menalled, F.D. Dryland organic farming partially offsets negative effects of highly simplified agricultural landscapes on forbs, bees, and bee–flower networks. Environ. Entomol. 2019, 48, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Blubaugh, C.K.; Kaplan, I. Tillage compromises weed seed predator activity across developmental stages. Biol. Control 2015, 81, 76–82. [Google Scholar] [CrossRef]
- Jowett, K.; Milne, A.E.; Garrett, D.; Potts, S.G.; Senapathi, D.; Storkey, J. Above- and below-ground assessment of carabid community responses to crop type and tillage. Agric. For. Entomol. 2020, 12397. [Google Scholar] [CrossRef]
- de Pedro, L.; Perera-Fernández, L.G.; López-Gallego, E.; Pérez-Marcos, M.; Sanchez, J.A. The effect of cover crops on the biodiversity and abundance of ground-dwelling arthropods in a Mediterranean pear orchard. Agronomy 2020, 10, 580. [Google Scholar] [CrossRef]
- Meyer, M.; Ott, D.; Götze, P.; Koch, H.; Scherber, C. Crop identity and memory effects on aboveground arthropods in a long-term crop rotation experiment. Ecol. Evol. 2019, 9, 7307–7323. [Google Scholar] [CrossRef]
- Patterson, E.S.P.; Sanderson, R.A.; Eyre, M.D. Soil tillage reduces arthropod biodiversity and has lag effects within organic and conventional crop rotations. J. Appl. Entomol. 2019, 143, 430–440. [Google Scholar] [CrossRef]
- Murrell, E.G.; Ray, S.; Lemmon, M.E.; Luthe, D.S.; Kaye, J.P. Cover crop species affect mycorrhizae-mediated nutrient uptake and pest resistance in maize. Renew. Agric. Food Syst. 2019, 1–8. [Google Scholar] [CrossRef]
- Mesbah, A.; Nilahyane, A.; Ghimire, B.; Beck, L.; Ghimire, R. Efficacy of cover crops on weed suppression, wheat yield, and water conservation in winter wheat–sorghum–fallow. Crop Sci. 2019, 59, 1745–1752. [Google Scholar] [CrossRef]
- Murrell, E.G. Can agricultural practices that mitigate or improve crop resilience to climate change also manage crop pests? Curr. Opin. Insect Sci. 2017, 23, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Wittwer, R.A.; Dorn, B.; Jossi, W.; Van Der Heijden, M.G.A. Cover crops support ecological intensification of arable cropping systems. Sci. Rep. 2017, 7, 41911. [Google Scholar]
- Lee, J.C.; Menalled, F.D.; Landis, D.A. Refuge habitats modify impact of insecticide disturbance on carabid beetle communities. J. Appl. Ecol. 2001, 38, 472–483. [Google Scholar] [CrossRef]
- Ward, M.J.; Ryan, M.R.; Curran, W.S.; Barbercheck, M.E.; Mortensen, D.A. Cover crops and disturbance influence activity-density of weed seed predators Amara aenea and Harpalus pensylvanicus (Coleoptera: Carabidae). Weed Sci. 2011, 59, 76–81. [Google Scholar] [CrossRef]
- McKenzie, S.C.; Goosey, H.B.; O’Neill, K.M.; Menalled, F.D. Integration of sheep grazing for cover crop termination into market gardens: Agronomic consequences of an ecologically based management strategy. Renew. Agric. Food Syst. 2017, 32, 389–402. [Google Scholar] [CrossRef]
- Kumar, S.; Sieverding, H.; Lai, L.; Thandiwe, N.; Wienhold, B.; Redfearn, D.; Archer, D.; Ussiri, D.; Faust, D.; Landblom, D.; et al. Facilitating crop–livestock reintegration in the northern great plains. Agron. J. 2019, 111, 2141–2156. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fixed Effect Variables | Activity Density | Species Diversity | Species Richness | Species Evenness | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DF | F-Value | p-Value | DF | F-Value | p-Value | DF | F-Value | p-Value | DF | F-Value | p-Value | |
| Year | 4,58 | 42.0 | <0.001 | 4,58 | 8.3 | <0.001 | 4,58 | 17.7 | <0.001 | 4,58 | 8.2 | <0.001 |
| Crop | 1,58 | 4.1 | <0.050 | 1,58 | 11.8 | 0.001 | 1,58 | 9.3 | 0.003 | 1,58 | 12.4 | <0.001 |
| System | 2,58 | 55.9 | <0.001 | 2,58 | 36.1 | <0.001 | 2,58 | 43.1 | <0.001 | 2,58 | 33.9 | <0.001 |
| Year: Crop | 4,58 | 13.3 | <0.001 | 4,58 | 4.1 | 0.005 | 4,58 | 6.0 | <0.001 | 4,58 | 3.5 | 0.010 |
| Year: System | 8,58 | 5.0 | <0.001 | 8,58 | 3.7 | 0.002 | 8,58 | 1.6 | 0.150 | 8,58 | 3.7 | 0.001 |
| Crop: System | 2,58 | 13.4 | <0.001 | 2,58 | 1.1 | 0.350 | 2,58 | 1.6 | 0.210 | 2,58 | 2.0 | 0.140 |
| Year: Crop: System | 8,47 | 0.6 | 0.810 | 8,47 | 0.2 | 0.990 | 8,47 | 0.5 | 0.850 | 8,47 | 0.4 | 0.910 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhikari, S.; Menalled, F.D. Supporting Beneficial Insects for Agricultural Sustainability: The Role of Livestock-Integrated Organic and Cover Cropping to Enhance Ground Beetle (Carabidae) Communities. Agronomy 2020, 10, 1210. https://doi.org/10.3390/agronomy10081210
Adhikari S, Menalled FD. Supporting Beneficial Insects for Agricultural Sustainability: The Role of Livestock-Integrated Organic and Cover Cropping to Enhance Ground Beetle (Carabidae) Communities. Agronomy. 2020; 10(8):1210. https://doi.org/10.3390/agronomy10081210
Chicago/Turabian StyleAdhikari, Subodh, and Fabian D. Menalled. 2020. "Supporting Beneficial Insects for Agricultural Sustainability: The Role of Livestock-Integrated Organic and Cover Cropping to Enhance Ground Beetle (Carabidae) Communities" Agronomy 10, no. 8: 1210. https://doi.org/10.3390/agronomy10081210
APA StyleAdhikari, S., & Menalled, F. D. (2020). Supporting Beneficial Insects for Agricultural Sustainability: The Role of Livestock-Integrated Organic and Cover Cropping to Enhance Ground Beetle (Carabidae) Communities. Agronomy, 10(8), 1210. https://doi.org/10.3390/agronomy10081210