Dolomite and Compost Amendments Enhance Cu Phytostabilization and Increase Microbiota of the Leachates from a Cu-Contaminated Soil

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Site Characteristics and Leachate Collection
2.2. Chemical Analysis of the Leachates and Microbial Toxicity Test
2.3. Cu Resistance of Culturable Bacteria
2.4. High throughput Sequencing Analysis
2.5. Statistical Analysis
3. Results
3.1. Chemical Parameters and Toxicity of the Soil Leachates
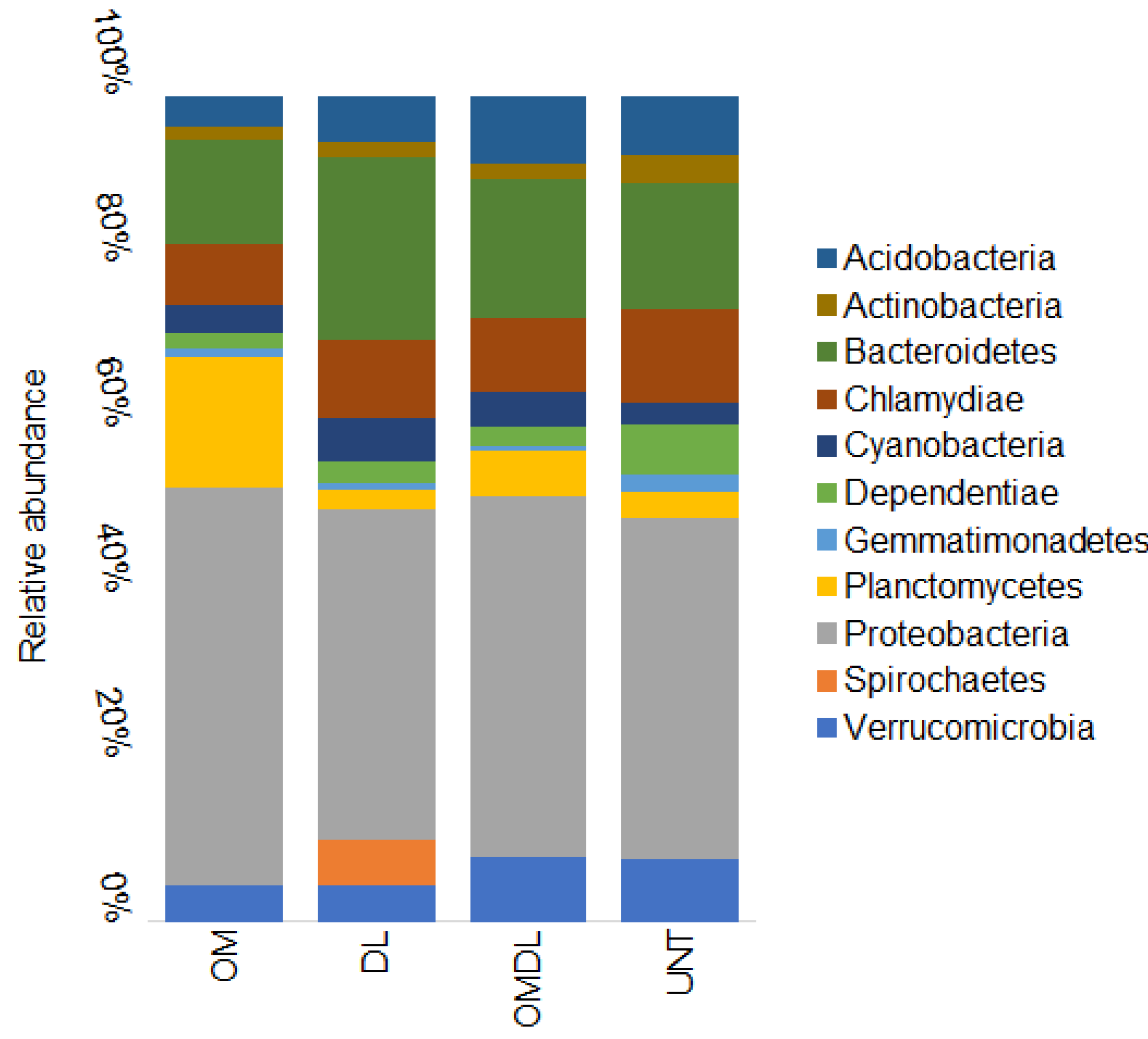
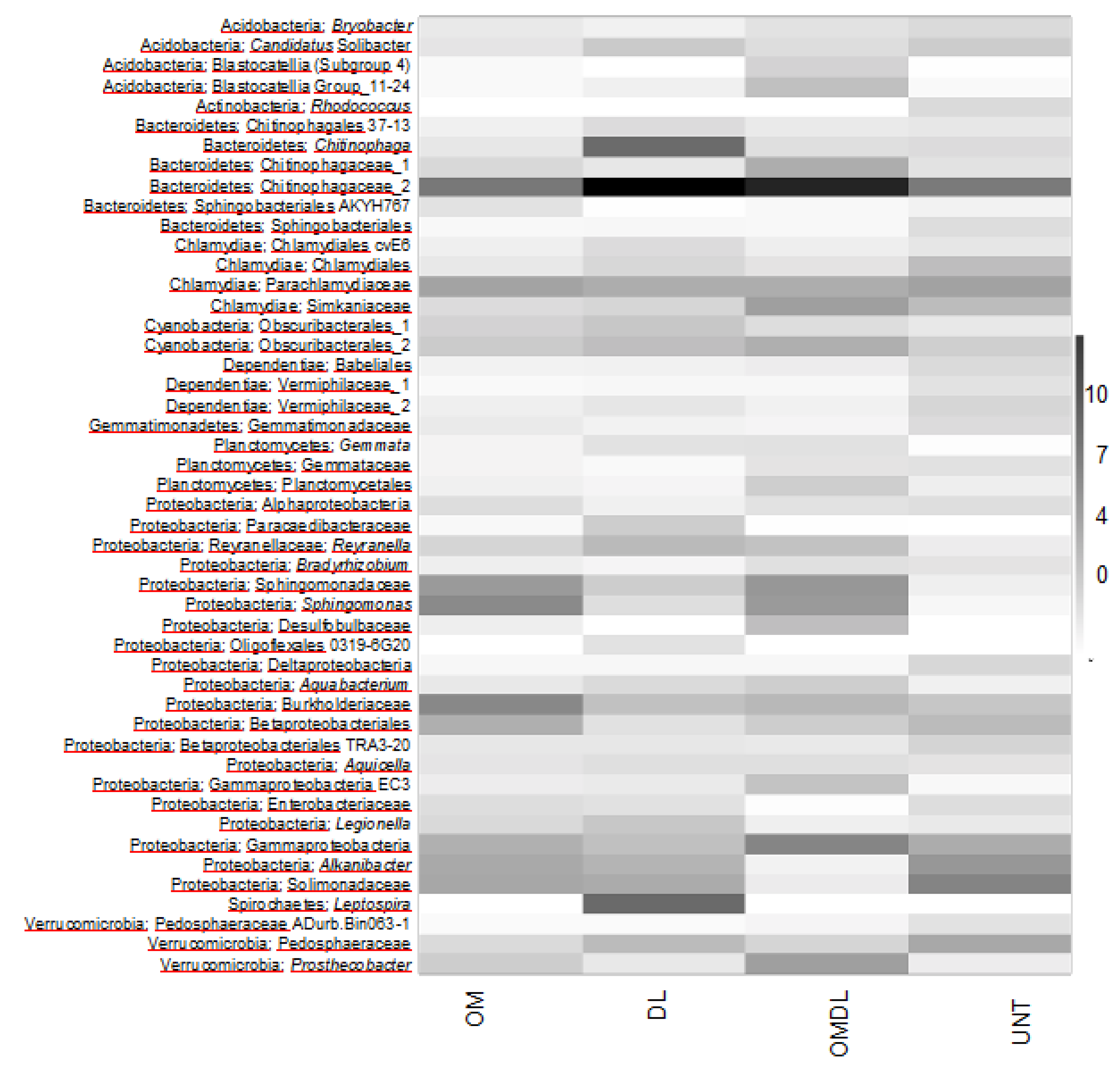
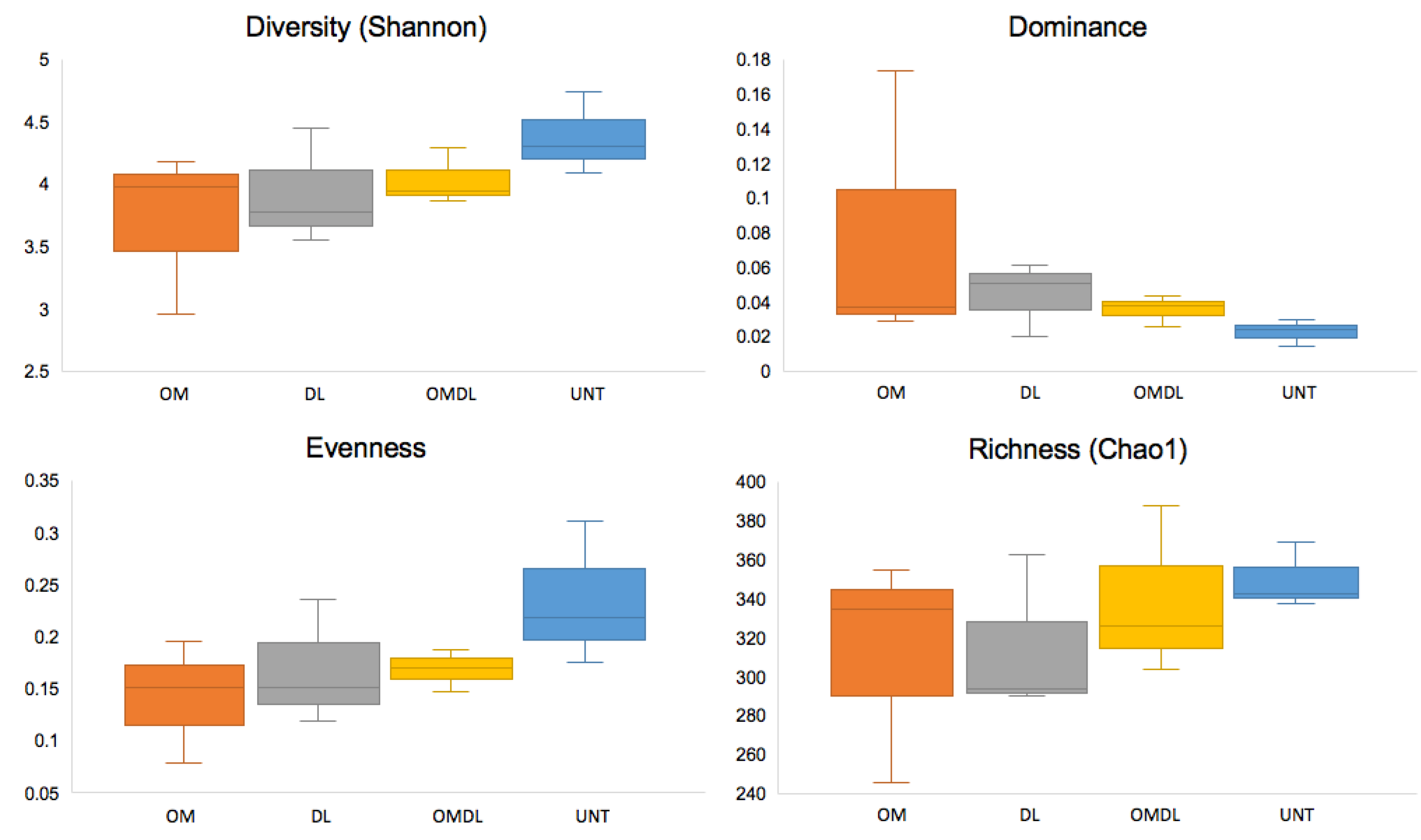
3.2. Microbial Diversity, Cultural Bacteria, and Cu Resistant Bacteria
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | 0–30 cm Soil Layer | 30–60 Soil Layer |
|---|---|---|
| Sand % | 85.8 | 85.5 |
| Silt% | 8.3 | 11.2 |
| Clay % | 5.9 | 3.3 |
| Organic matter (g/Kg) | 16 | 5.42 |
| C/N | 17.2 | 8.63 |
| CEC (cmol/Kg) | 3.5 | 1.17 |
| pH | 7 | 4.04 |
| As (mg/Kg) | 9.8 | 4.7 |
| Cd (mg/Kg) | 0.12 | |
| Co (mg/Kg) | <2 | 2.3 |
| Cu (mg/Kg) | 1110-1460 | 111-153 |
| Cr (mg/Kg) | 23 | 18.4 |
| Fe (mg/Kg) | 6090 | 7900 |
| Mn (mg/Kg) | 181 | 185 |
| Ni (mg/Kg) | 5 | 8 |
| Pb (mg/Kg) | 27 | |
| Tl (mg/Kg) | 0.24 | |
| Zn (mg/Kg) | 46 | 28.7 |
| pCu2+ | 7.66 | 5.12 |
References
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. The Earth microbiome project consortium. A communal catalogue reveals Earth’s multiscale microbial diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xia, B.; Treves, D.S.; Wu, L.-Y.; Marsh, T.L.; O’Neill, R.V.; Palumbo, A.V.; Tiedje, J.M. Spatial and resource factors influencing high microbial diversity in soil. Appl. Environ. Microbiol. 2002, 68, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Vos, M.; Wolf, A.B.; Jennings, S.J.; Kowalchuk, G.A. Micro-scale determinants of bacterial diversity in soil. FEMS Microbiol. Rev. 2013, 37, 936–954. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.; Newman, J.A.; Silverman, B.W.; Turner, S.L.; Lilley, A.K. The contribution of species richness and composition to bacterial services. Nature 2005, 436, 1157–1160. [Google Scholar] [CrossRef]
- Jousset, A.; Bienhold, C.; Chatzinotas, A.; Gallien, L.; Gobet, A.; Kurm, V.; Küsel, K.; Rillig, M.C.; Rivett, D.W.; Salles, J.F.; et al. Where less may be more: How the rare biosphere pulls ecosystems strings. ISME J. 2017, 11, 853–862. [Google Scholar] [CrossRef]
- George, P.B.L.; Lallias, D.; Creer, S.; Seaton, F.M.; Kenny, J.G.; Eccles, R.M.; Griffiths, R.I.; Lebron, I.; Emmett, B.A.; Robinson, D.A.; et al. Divergent national-scale trends of microbial and animal biodiversity revealed across diverse temperate soil ecosystems. Nat. Commun. 2019, 10, 1107. [Google Scholar] [CrossRef]
- Kaiser, K.; Wemheuer, B.; Korolkow, V.; Wemheuer, F.; Nacke, H.; Schöning, I.; Schrumpf, M.; Daniel, R. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Sci. Rep. 2016, 6, 33696. [Google Scholar] [CrossRef]
- Heuer, H.; Smalla, K. Bacterial phyllosphere communities of Solanum tuberosum L. and T4-lysozyme-producing transgenic variants. FEMS Microbiol. Ecol. 1999, 28, 357–371. [Google Scholar] [CrossRef]
- Crecchio, C.; Gelsomino, A.; Ambrosoli, R.; Minati, J.L.; Ruggiero, P. Functional and molecular responses of soil microbial communities under differing soil management practices. Soil Biol. Biochem. 2004, 36, 1873–1883. [Google Scholar] [CrossRef]
- Hendgen, M.; Hoppe, B.; Döring, J.; Friedel, M.; Kauer, R.; Frisch, M.; Dahl, A.; Kellner, H. Effects of different management regimes on microbial biodiversity in vineyard soils. Sci. Rep. 2018, 8, 9393. [Google Scholar] [CrossRef] [PubMed]
- Wilpiszeski, R.L.; Aufrecht, J.A.; Retterer, S.T.; Sullivan, M.B.; Graham, D.E.; Pierce, E.M.; Zablocki, O.D.; Palumbo, A.V.; Elias, D.A. Soil aggregate microbial communities: Towards understanding microbiome interactions at biologically relevant scales. Appl. Environ. Microbiol. 2019, 85, e00324-19. [Google Scholar] [CrossRef]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Tecon, R.; Or, D. Biophysical processes supporting the diversity of microbial life in soil. FEMS Microbiol. Rev. 2017, 41, 599–623. [Google Scholar] [CrossRef] [PubMed]
- Nunan, N.; Leloup, J.; Ruamps, L.S.; Pouteau, V.; Chenu, C. Effects of habitat constraints on soil microbial community function. Sci. Rep. 2017, 7, 4280. [Google Scholar] [CrossRef]
- Powelson, D.K.; Simpson, J.R.; Gerba, C.P. Effects of organic matter on virus transport in unsaturated flow. Appl. Environ. Microbiol. 1991, 57, 2192–2196. [Google Scholar] [CrossRef]
- Scholl, M.A.; Harvey, R.W. Laboratory investigations on the role of sediment surface and groundwater chemistry in transport of bacteria through a contaminated sandy aquifer. Environ. Sci. Technol. 1992, 26, 1410–1417. [Google Scholar] [CrossRef]
- Dechesne, A.; Wang, G.; Gülez, G.; Or, D.; Smets, B.F. Hydration controlled bacterial motility and dispersal on surfaces. Proc. Natl. Acad. Sci. USA 2010, 107, 14369–14372. [Google Scholar] [CrossRef]
- McBride, M.B. Reactions controlling heavy metal solubility in soils. In Advances in Soil Science; Stewart, B.A., Ed.; Springer-Verlag: New York, NY, USA, 1989; pp. 1–56. [Google Scholar]
- Sauvé, S.; Dumestre, A.; McBride, M.B.; Hendershot, W.H. Soil quality criteria using chemical speciation of Pb2+ and Cu2+. Environ. Toxicol. Chem. 1998, 17, 1481–1489. [Google Scholar] [CrossRef]
- Odem, W.I.; Amy, G.L.; Conklin, M. Subsurface interactions of humic substances with Cu(II) in saturated media. Environ. Sci. Technol. 1993, 27, 1045–1049. [Google Scholar] [CrossRef]
- Osman, D.; Cavet, J.S. Copper homeostasis in bacteria. Adv. Appl. Microbiol. 2008, 65, 217–247. [Google Scholar] [PubMed]
- Ballabio, C.; Panagos, P.; Lugato, E.; Huang, J.-H.; Orgiazzi, A.; Jones, A.; Fernández-Ugalde, O.; Borrelli, P.; Montanarella, L. Copper distribution in European topsoils: An assessment based on LUCAS soil survey. Sci. Total Environ. 2018, 636, 282–298. [Google Scholar] [CrossRef] [PubMed]
- Bes, C.M.; Mench, M.; Aulen, M.; Gaste, H.; Taberly, J. Spatial variation of plant communities and shoot Cu concentrations of plant species at a timber treatment site. Plant Soil 2010, 330, 267–280. [Google Scholar] [CrossRef]
- Rehman, M.; Liu, L.J.; Wang, Q.; Saleem, M.H.; Bashir, S.; Ullah, S.; Peng, D.X. Copper environmental toxicology, recent advances, and future outlook: A review. Environ. Sci. Poll. Res. 2019, 26, 18003–18016. [Google Scholar] [CrossRef]
- Brandt, K.K.; Frandsen, R.J.N.; Holm, P.E.; Nybroe, O. Development of pollution-induced community tolerance is linked to structural and functional resilience of a soil bacterial community following a five-year field exposure to copper. Soil Biol. Biochem. 2010, 42, 748–757. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Chu, G.X.; Lardner, R.; Liang, Y.C.; McLaughlin, M.J. A single application of Cu to field soil has long-term effects on bacterial community structure, diversity, and soil processes. Pedobiologia 2010, 53, 149–158. [Google Scholar] [CrossRef]
- Macdonald, C.A.; Clark, I.M.; Zhao, F.J.; Hirsch, P.R.; Singh, B.K.; McGrath, S.P. Long-term impacts of zinc and copper enriched sewage sludge additions on bacterial, archaeal and fungal communities in arable and grassland soils. Soil Biol. Biochem. 2011, 43, 932–941. [Google Scholar] [CrossRef]
- Burges, A.; Fievet, V.; Oustriere, N.; Epelde, L.; Garbisu, C.; Becerril, J.M.; Mench, M. Long-term phytomanagement with compost and a sunflower—Tobacco rotation influences the structural microbial diversity of a Cu-contaminated soil. Sci. Total Environ. 2020, 700, 134529. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Chu, G.X.; Broos, K.; Clarke, K.R.; Liang, Y.C.; McLaughlin, M.J. Structural and functional response of soil microbiota to addition of plant substrate are moderated by soil Cu levels. Biol. Fertil. Soils. 2010, 46, 333–342. [Google Scholar] [CrossRef]
- Khalid, S.; Shahid, M.; Niazi, N.K.; Murtaza, B.; Bibi, I.; Dumat, C.A. Comparison of technologies for remediation of heavy metal contaminated soils. J. Geochem. Explor. 2016, 182, 247–268. [Google Scholar] [CrossRef]
- Guo, X.; Wei, Z.; Wu, Q.; Li, C.; Qian, T.; Zheng, W. Effect of soil washing with only chelators or combining with ferric chloride on soil heavy metal removal and phyto availability: Field experiments. Chemosphere 2016, 147, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Race, M.; Marotta, R.; Fabbricino, M.; Pirozzi, F.; Andreozzi, R.; Cortese, L.; Giudicianni, P. Copper and zinc removal from contaminated soils through soil washing process using ethylenediamine disuccinic acid as a chelating agent: A modeling investigation. J. Environ. Chem. Engin. 2016, 4, 2878–2891. [Google Scholar] [CrossRef]
- Kidd, P.S.; Álvarez-López, V.; Becerra-Castro, C.; Cabello-Conejo, M.; Prieto-Fernández, Á. Potential Role of Plant-Associated Bacteria in Plant Metal Uptake and Implications in Phytotechnologies. Adv. Bot. Res. 2017, 83, 87–126. [Google Scholar]
- Touceda-Gonzalez, M.; Prieto-Fernandez, A.; Renella, G.; Giagnoni, L.; Sessitsch, A.; Brader, G.; Kumpiene, J.; Dimitriou, I.; Eriksson, J.; Friesl-Hanl, W.; et al. Microbial community structure and activity in trace element-contaminated soils phytomanaged by Gentle Remediation Options (GRO). Environ. Pollut. 2017, 231, 237–251. [Google Scholar] [CrossRef]
- Narula, N.; Reinicke, M.; Haferburg, G.; Kothe, E.; Behl, R.K. Plant–Microbe Interaction in Heavy-Metal-Contaminated Soils. In Bio-Geo Interactions in Metal-Contaminated; Kothe, E., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 143–162. [Google Scholar]
- Marchand, L.; Mench, M.; Marchand, C.; Lecoustumer, P.; Kolbas, A.; Maalouf, J.P. Phytotoxicity testing of lysimeter leachates from aided phytostabilized Cu-contaminated soils using duckweed (Lemna minor L.). Sci. Total Environ. 2011, 410, 146–153. [Google Scholar] [CrossRef]
- Bes, C.; Mench, M. Remediation of copper-contaminated topsoils from a wood treatment facility using in situ stabilisation. Environ. Pollut. 2008, 156, 128–139. [Google Scholar] [CrossRef]
- Lappalainen, J.; Juvonen, R.; Vaajasaari, K.; Karp, M. A new flash method for measuring the toxicity of solid and colored samples. Chemosphere 1999, 38, 1069–1083. [Google Scholar] [CrossRef]
- Mergeay, M. Heavy metal resistance in microbial ecosystems. In Molecular Microbial Ecology Manual; Akkermans, A.D.L., Van Elsas, J.D., De Bruijn, F.J., Eds.; Springer: Dordrecht, The Netherlands, 1995; Volume 6, pp. 7–17. [Google Scholar]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef]
- Vieira, C.K.; dos Anjos Borges, L.G.; Marconatto, L.; Giongo, A.; Stürmer, S.L. Microbiome of a revegetated iron-mining site and pristine ecosystems from the Brazilian Cerrado. Appl. Soil Ecol. 2018, 131, 55–65. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Meth. 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Juang, K.W.; Lee, Y.I.; Lai, H.Y.; Chen, B.C. Influence of magnesium on copper phytotoxicity to and accumulation and translocation in grapevines. Ecotoxicol. Environ. Saf. 2014, 104, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Ambrosini, V.G.; Rosa, D.J.; Prado, J.P.C.; Borghezan, M.; Melo, G.W.B.; Soares, J.J.; Comin, C.R.F.S.; Simao, D.G.; Brunetto, G. Plant Physiology and Biochemistry Reduction of copper phytotoxicity by liming: A study of the root anatomy of young vines (Vitis labrusca L.). Plant Physiol. Biochem. 2015, 96, 270–280. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Nakata, P.A. Calcium oxalate in plants: Formation and Function. Ann. Rev. Plant Biol. 2005, 56, 41–71. [Google Scholar] [CrossRef]
- Yruela, I. Copper in plants: Acquisition, transport and interactions. Funct. Plant Biol. 2009, 36, 409–430. [Google Scholar] [CrossRef]
- Fan, J.H.; He, Z.L.; Ma, L.Q.; Yang, Y.G.; Yang, X.E.; Stoffella, P.J. Immobilization of copper in contaminated sandy soils using calcium water treatment residue. J. Hazard. Mater. 2011, 189, 708–710. [Google Scholar] [CrossRef]
- Trentin, E.; Basso Facco, D.; Hammerschmitt, R.K.; Ferreira, P.A.A.; Morsch, L.; Belles, S.W.; Ricachenevsky, F.K.; Nicoloso, F.T.; Ceretta, C.A.; Tiecher, T.L.; et al. Potential of vermicompost and limestone in reducing copper toxicity in young grapevines grown in Cu-contaminated vineyard soil. Chemosphere 2019, 226, 421–430. [Google Scholar] [CrossRef]
- Bender, M.; Conrad, R. Effect of CH4 concentration and soil conditions on the induction of CH4 oxidation activity. Soil Biol. Biochem. 1995, 27, 1517–1527. [Google Scholar] [CrossRef]
- Roesch, L.F.; Fulthorpe, R.R.; Riva, A.; Casella, G.; Hadwin, A.K.; Kent, A.D.; Daroub, S.H.; Camargo, F.A.; Farmerie, W.G.; Triplett, E.W. Pyrosequencing enumerates and contrasts soil microbial diversity. ISME J. 2007, 1, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.Y.; Xi, L.L.; Jin, D.C.; Mi, B.B.; Wang, D.H.; Li, X.F.; Dai, X.Z.; Zou, X.X.; Zhang, Z.; Ma, Y.Q.; et al. Bacterial community response to cadmium contamination of agricultural paddy soil. Appl. Soil Ecol. 2019, 139, 100–106. [Google Scholar] [CrossRef]
- Schneider, A.R.; Gommeaux, M.; Duclercq, J.; Fanin, N.; Conreux, A.; Alahmad, A.; Lacoux, J.; Roger, D.; Spicher, F.; Ponthieu, M.; et al. Response of bacterial communities to Pb smelter pollution in contrasting soils. Sci. Total Environ. 2017, 605, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, G.; Barua, S.; Sani, R.K.; Peyton, B.M. Investigation of microbial populations in the extremely metal-contaminated Coeur d’Alene River sediments. Microb. Ecol. 2011, 62, 1–13. [Google Scholar] [CrossRef]
- Li, X.; Qu, C.; Bian, Y.; Gu, C.; Jiang, X.; Song, Y. New insights into the responses of soil microorganisms to polycyclic aromatic hydrocarbon stress by combining enzyme activity and sequencing analysis with metabolomics. Environ. Pollut. 2019, 255, 113312. [Google Scholar] [CrossRef]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil. Appl. Environ. Microb. 2011, 77, 6295–6300. [Google Scholar] [CrossRef]
- Rappé, M.S.; Giovannoni, S.J. The uncultured microbial majority. Annu. Rev. Microbiol. 2003, 57, 369–394. [Google Scholar] [CrossRef]
- Ramírez-Díaz, M.I.; Díaz-Pérez, C.; Vargas, E.; Riveros-Rosas, H.; Campos-García, J.; Cervantes, C. Mechanisms of bacterial resistance to chromium com- pounds. Biometals 2008, 21, 321–332. [Google Scholar] [CrossRef]
- Berg, J.; Brandt, K.K.; Al-Soud, W.A.; Holm, P.E.; Hansen, L.H.; Sørensen, S.J.; Nybroe, Ø. Selection for Cu-tolerant bacterial communities with altered composition, but unaltered richness, via long-term Cu exposure. Appl. Environ. Microbiol. 2012, 78, 7438–7446. [Google Scholar] [CrossRef]
- Ventorino, V.; Pascale, A.; Adamo, P.; Rocco, C.; Fiorentino, N.; Mori, M.; Faraco, V.; Pepe, O.; Fagnano, M. Comparative assessment of autochthonous bacterial and fungal communities and microbial biomarkers of polluted agricultural soils of the Terra dei Fuochi. Sci. Rep. 2018, 8, 14281. [Google Scholar] [CrossRef]
- Virto, I.; Gartzia-Bengoetxea, N.; Fernandez-Ugalde, O. Role of organic matter and carbonates in soil aggregation estimated using laser diffractometry. Pedosphere 2011, 21, 566–572. [Google Scholar] [CrossRef]
- Yao, F.; Yang, S.; Wang, Z.; Wang, X.; Ye, J.; Wang, X.; DeBruyn, J.M.; Feng, X.; Jiang, Y.; Li, H. Microbial taxa distribution is associated with ecological trophic cascades along an elevation gradient. Front. Microbiol. 2017, 8, 2071. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.; Di Lonardo, D.P.; Bodelier, P.L.E. Revisiting life strategy concepts in environmental microbial ecology. FEMS Microbiol. Ecol. 2017, 93, 1–14. [Google Scholar] [CrossRef]
- Lejon, D.P.H.; Pascault, N.; Ranjard, L. Differential copper impact on density, diversity and resistance of adapted culturable bacterial populations according to soil organic status. Eur. J. Soil Biol. 2010, 46, 168–174. [Google Scholar] [CrossRef]
- Tsai, K.P. Management of target algae by using copper-based algaecides: Effects of algal cell density and sensitivity to copper. Water Air Soil Pollut. 2016, 227, 1–11. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef]




| Treatment | pH | EC (µS) | Ca (mg L−1) | Mg (mg L−1) | Cu Concentration (μM) | Total Organic C (mg L−1) | Total N (mg kg−1) | Microbial Toxicity (Inhibition %) |
|---|---|---|---|---|---|---|---|---|
| Unt | 5.91 (±0.32) a | 255 (±75) a | 4.70 (±1.33) a | 1.21 (±0.17) a | 17.0 (±0.37) a | 12.9 (±5.8) a | 1.15 (±0.16) b | 28.9 (±5.4) a |
| DL | 5.83 (±0.32) a | 121 (±19) b | 8.78 (±4.39) a | 2.66 (±0.66) bc | 7.40 (±0.12) b | 7.58 (±1.5) b | 1.05 (±0.18) b | 11.4 (±3.9) c |
| OM | 5.63 (±0.09) a | 124 (±37) b | 5.15 (±0.57) a | 1.85 (±0.20) ab | 16.2 (±0.13) a | 13.7 (±1.1) a | 1.56 (±0.13) a | 22.5 (±5.0) b |
| OMDL | 5.81 (±0.75) a | 155 (±19) b | 10.25 (±2.21) a | 3.14 (±0.21) c | 6.77 (±0.22) b | 14.8 (±3.36) a | 0.86 (±0.73) b | 9.6 (±5.0) c |
| Treatment | Total Culturable Bacteria (CFU mL−1) | Oligotrophic Bacteria (CFU mL−1) | Cu Resistant Bacteria (CFU mL−1) | Cu Resistant Bacteria (%) |
|---|---|---|---|---|
| Unt | 2.4 (±2.1) × 103 b | 2.7 (±2.1) × 103 b | 5.8 (±1.9) × 102 b | 21.4 (±2.1) a |
| DL | 9.4 (±3.7) × 103 b | 1.5 (±3.7) × 104 a | 5.6 (±1.0) × 102 b | 3.8 (±1.3) d |
| OM | 2.8 (±2.6) × 104 a | 8.3 (±2.6) × 104 a | 1.3 (±2.8) × 103 a | 15.6 (±2.4) b |
| OMDL | 1.0 (±0.1) × 104 a | 1.4 (±0.1) × 104 a | 1.3 (±4.7) × 103 a | 9.8 (±2.1) c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giagnoni, L.; dos Anjos Borges, L.G.; Giongo, A.; de Oliveira Silveira, A.; Ardissone, A.N.; Triplett, E.W.; Mench, M.; Renella, G. Dolomite and Compost Amendments Enhance Cu Phytostabilization and Increase Microbiota of the Leachates from a Cu-Contaminated Soil. Agronomy 2020, 10, 719. https://doi.org/10.3390/agronomy10050719
Giagnoni L, dos Anjos Borges LG, Giongo A, de Oliveira Silveira A, Ardissone AN, Triplett EW, Mench M, Renella G. Dolomite and Compost Amendments Enhance Cu Phytostabilization and Increase Microbiota of the Leachates from a Cu-Contaminated Soil. Agronomy. 2020; 10(5):719. https://doi.org/10.3390/agronomy10050719
Chicago/Turabian StyleGiagnoni, Laura, Luiz Gustavo dos Anjos Borges, Adriana Giongo, Andressa de Oliveira Silveira, Alexandria N. Ardissone, Eric W. Triplett, Michel Mench, and Giancarlo Renella. 2020. "Dolomite and Compost Amendments Enhance Cu Phytostabilization and Increase Microbiota of the Leachates from a Cu-Contaminated Soil" Agronomy 10, no. 5: 719. https://doi.org/10.3390/agronomy10050719
APA StyleGiagnoni, L., dos Anjos Borges, L. G., Giongo, A., de Oliveira Silveira, A., Ardissone, A. N., Triplett, E. W., Mench, M., & Renella, G. (2020). Dolomite and Compost Amendments Enhance Cu Phytostabilization and Increase Microbiota of the Leachates from a Cu-Contaminated Soil. Agronomy, 10(5), 719. https://doi.org/10.3390/agronomy10050719