1. Introduction
Cover crop (CC) residue is a source of soil organic matter, and its degradation is critical to subsequent crop productivity. Residue decomposition determines the soil nutrient pool and regulates nutrient release in soil [
1] through depolymerization of fibers and hydrolysis of sugars, mostly via heterotrophic soil microorganisms [
2]. External environmental conditions as modified by climate, agronomic practices such as tillage, cropping rotation, fertilizer, and irrigation, and plant residue quality may impact the rate of decomposition and nutrient release. Inherent properties of the residue such as carbon-nitrogen (CN) ratio, fiber fractions, and lignin concentration can greatly affect the litter decomposition and nutrient cycling [
3,
4,
5]. These properties differ between C3- and C4-derived soil organic matter [
6] and between grass and legume residue [
7], which may impact decomposition and nutrient release kinetics, indicating the possibility of the usefulness of the different approaches for modeling those kinetics. The choice of approach also depends on the desired degree of analytical simplicity, predictive power, and generality [
8]. Knowledge of decay mechanism and use of a suitable model specific to the substrate quality can provide valuable information for CC management, which is mostly lacking in comparative studies where a single model is chosen for a variety of crops to determine decay rate constants and half-lives. There is a lack of uniformity in using decay models for decomposition and mineralization studies for similar substrates, which vary from simple one parametric single exponential first-order models to the complex multiparametric consecutive exponential models.
A first-order single exponential decay model (
y = ae−bx, where
y is the mass of substrate at time
x,
b is the rate constant, and
e is the base of the natural logarithms (2.71828)) has been widely used for nutrient mineralization, residue decomposition, and plant population studies [
5,
7,
9,
10,
11,
12]. It was applied for modeling litter decomposition for numerous grasses and legumes [
7] and fine litter decomposition of forest soil [
12]. Ruffo and Bollero [
13] and Sievers and Cook [
5] used this model with an asymptote (
y =
ae−bx +
yₒ, where
yₒ is an asymptote) for cereal rye (
Secale cereale L.) and hairy vetch (
Vicia villosa Roth) decomposition and nutrient release. Polglase et al. [
14] used a single exponential model for P mineralization from soil organic matter in a pine forest, whereas Fernández et al. [
15] used it for modeling C mineralization in soils after wildfires in Spain. The strength of this model is that it produces a single rate constant, which can be used directly to compare decay rates from different treatments. However, it does not accurately describe decomposition or mineralization kinetics, where rate constants vary with time due to rapid loss or an extended lag phase in early decomposition [
9,
12,
16]. The CC-derived labile fraction of soil organic matter composed of light (low specific density or mineral-free) and heavy (high specific density or mineral-bound) fractions tends to follow a different kinetic model in describing the decomposition and mineralization [
17,
18].
The first-order double exponential model with two rate constants (
y =
ae−bx + ce−dx, where
b and
d are the rate constants) separates organic matter into a soluble fraction (e.g., sucrose) or fast pool and cell-wall (e.g., detergent fibers) or slow pool [
19] fraction. It was reported to have improved goodness of fit of single exponential models for residue decomposition and nutrient release mechanisms [
9,
18,
20]. Berndt [
9] suggested this model over the single exponential model when comparing kinetic parameters of decay of C remaining for hybrid bermudagrass (
Cynodon dactylon (L.) Pers. ×
Cynodon transvaalensis Burtt-Davy) thatch. Wang et al. [
20] predicted temperature- and moisture-dependent rate constants for soil N mineralization with a modified double exponential model under standard temperature (35 °C) and moisture conditions (55% water holding capacity). Dhakal et al. [
11] reported that the double exponential model described alfalfa (
Medicago sativa L.) population decline in a semiarid environment, which resulted in the highest adjusted
R2 (0.94 to 0.97) and the lowest standard error of estimate (SEE) for the upright-type alfalfa cultivars. Fernandez et al. [
15] and Camargo et al. [
21] reported this model as fitting mineralization data better than a single exponential model. Although all models generated an
R2 greater than 0.98 for in vitro mineralization of C, the double exponential model could not fit some of the samples, whereas exponential with a linear combination (
y =
ae−bx + cx +
yₒ, where
c is the slope of the linear function) yielded superior results to the double exponential, exponential plus an asymptote, and hyperbolic model [
y =
ab/(
b + x)] [
17]. Dendooven et al. [
22] reported poor fit of the double exponential function in fitting N mineralization data to characterize active and recalcitrant organic N pools derived from sugar-beet (
Beta vulgaris L.) and bean (
Phaseolus vulgaris L.) residue.
Besides exponential models, a hyperbolic function was recommended to minimize standard errors, compared to the first-order exponential model in fitting the N mineralization data [
16]. Decay and N release of cereal rye and hairy vetch residue has been well described using the hyperbolic model when compared to linear and first-order models [
23]. In contrast, Berndt [
9] reported poor fit statistics for the two-parametric hyperbolic decay function, relative to exponential models. This indicates the need to test various empirical models, specific to the plant species. Since mass loss and N release from cereal rye and hairy vetch residue have been studied using a variety of empirical models [
5,
13,
23], performances of those functions have not been compared yet to suggest the best fit model.
The current study provides an overview of performances of the commonly used empirical models in CC decomposition and N mineralization studies. The objective of this research was to examine eight mathematical models to test their statistical significance in explaining cereal rye and hairy vetch decay and N mineralization in a sub-humid environment. Mass and N remaining of CC residue were fitted with six exponential and two hyperbolic models, and statistical parameters were compared for those models. An empirical model with the highest adjusted
R2 and lowest residual sum of squares and Akaike information criterion (AIC; [
24]) may be considered best for decomposition studies.
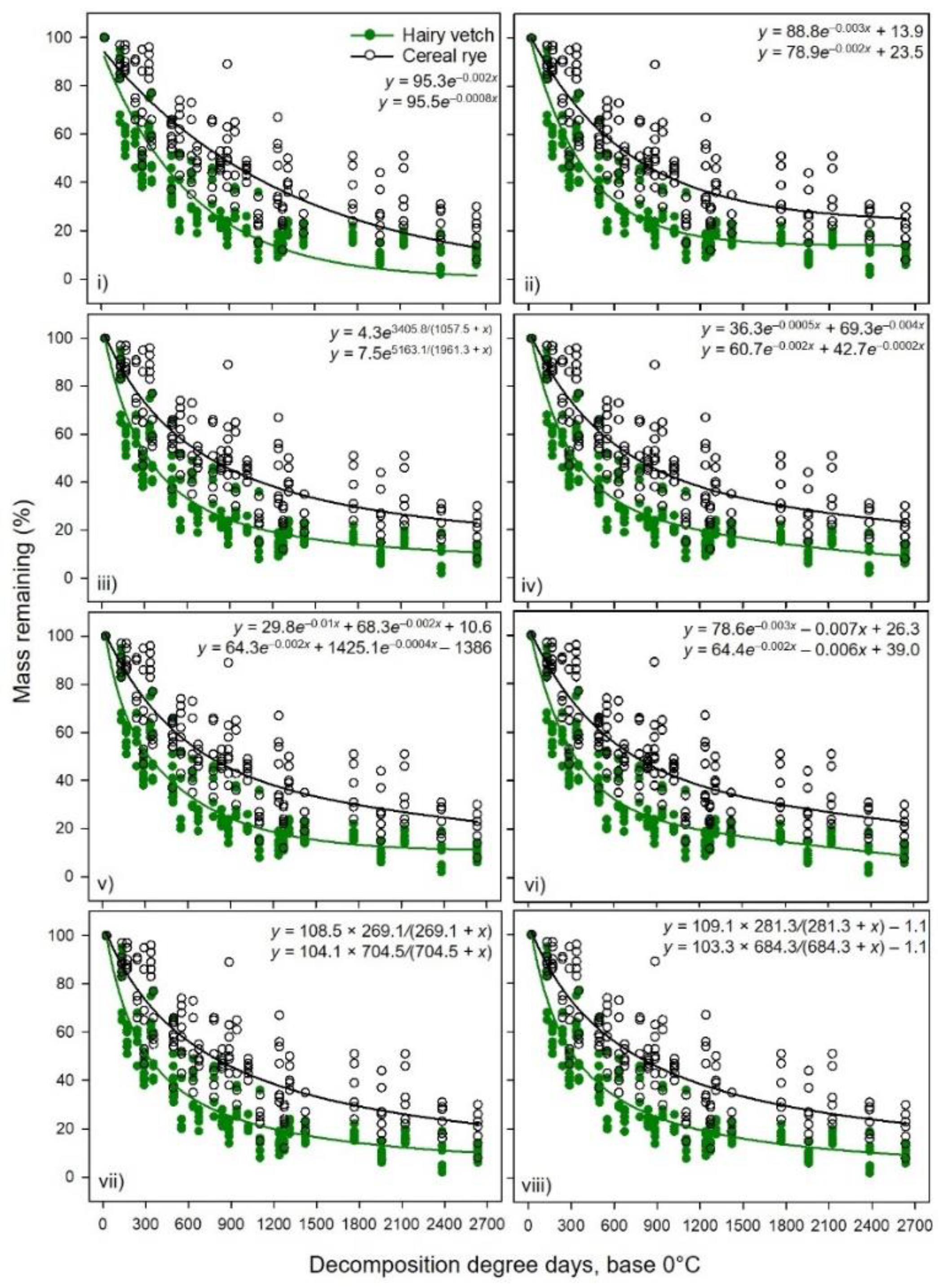
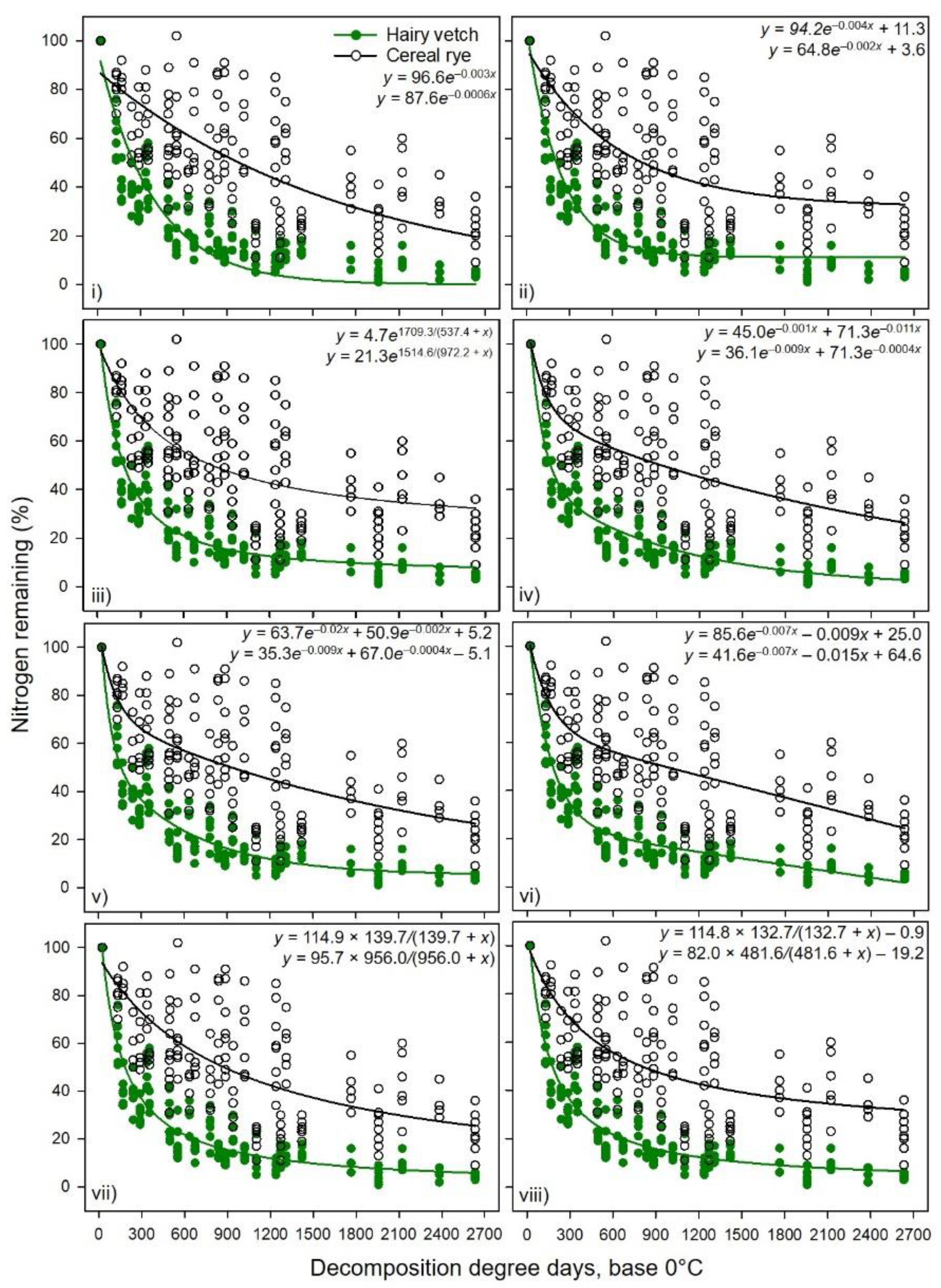
4. Discussion
Results showed a stronger relationship between percent mass or N remaining (high adjusted R
2,
Table 1,
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7 and
Table 8) of hairy vetch CC residue and cumulative DDD than that of cereal rye residue (
Figure 1 and
Figure 2). The decomposition rate constants and asymptotes were dissimilar for these two CCs (
Table 1,
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7 and
Table 8). Although Exp. 2 had two tillage systems (reduced vs. no-tillage), there was no effect on decomposition rate constant and N release in 2017 and 2018 [
25], indicating that the management factor had lesser impact than the substrate quality factor. The inherent plant chemical constituents determine the rate of decomposition and N mineralization into the soil [
4]. In both CC decomposition experiments, the CN ratio was greater for cereal rye (35:1 and 24:1 for Exp. 1 and 2, respectively) than hairy vetch (10:1, 9:1 for Exp. 1 and 2, respectively) [
5,
25]. Greater N concentration and less fiber content in hairy vetch may have accelerated the decomposition and N release in the early days or with less accumulated DDD, a relationship reported by Ruffo and Bollero [
13] and Otte et al. [
39]. Nitrogen and soluble carbohydrate fraction of the plant residue enhanced the microbial growth efficiency and improved the bermudagrass residue decomposition in FL, USA [
9]. In both experiments, the residuals were higher for the cereal rye mass remaining and N release data than for hairy vetch. The lack of fit was due to large variability in initial mass and N content of cereal rye and immobilization of the N in the first 4 wks [
5]. Rufo and Bollero [
13] also reported the lack of fit for cereal rye when compared to hairy vetch. The presence of high neutral detergent fiber and lignin concentration [
5] likely influenced the tensile strength of leaves and decomposability of cereal rye, which provide mechanical and chemical defense against microbial and chemical degradation [
40]. Cornelissen et al. [
41] reported that leaf tensile strength was related to litter decomposition for C3 grasses and also suggested that the complex leaf base content of the grass species can result in high variation in mass and N loss.
The greater adjusted R
2 values and lower standard errors with the double exponential model with two rate constants when compared to a single exponential with or without an asymptote, especially for hairy vetch CC residue, indicated two residue pools, i.e., fast and slow decomposing fractions. This could be due to the presence of labile versus and recalcitrant fractions of the plant materials. However, in some cases, the addition of asymptotic or linear or another exponential component to the simple single exponential model resulted in non-finite residuals and failed to cross-validate the model, mainly for cereal rye for individual datasets from Exp. 1 and 2. In such cases, the slopes (or rate constants) for the double exponential model or exponential model with the linear combination had poor predictability at α ≤ 0.01. This indicates that the sample size was too small to have a sufficient sampling frame for the model, which gave undue weight to the initial data points for the short study period. We noticed that the issue of non-finite residuals was eliminated with combined data from Exp. 1 and 2, which extended the x-axis from 14 wk (cumulative DDD = 2382) to 16 wk (cumulative DDD = 2633) and increased the number of XY pairs. That was the reason Otte et al. [
39] suggested using the simple asymptotic model rather than a double exponential model for cereal rye biomass decomposition. However, an asymptotic model may not always give the best results when compared to non-asymptotic models. The plant chemical constituents and environmental conditions may also affect the model performances [
40,
42]. Both of our experiments received more than half of the total cumulative rain in the first few weeks and the volumetric soil water content was near the field capacity, which might help accelerate the residue degradation as most of the soil microbes are highly active and thrive under moist warm conditions [
42]. Hairy vetch aboveground biomass was well represented by a double exponential model with an asymptote with high R
2 and minimal residual errors and AIC with shorter fast-pool turnover time (~25% of the total accumulated DDD), compared to that of cereal rye (~40% of the total accumulated DDD). Juma et al. [
16] suggested that a model of N cycling in soil should have at least three to four compartments to account for different sources of N.
Only hyperbolic models produced relatively low RMS, SEE, AIC, and PRESS statistics for the belowground hairy vetch and cereal rye percent mass and N remaining, while exponential models showed poorer fits. The fast-pool turnover time of belowground residue mass was shorter than that of aboveground biomass owing to its immediate proximity to soil biotic and abiotic environments [
43,
44,
45]. Similar to aboveground biomass, there was a difference in mass loss and N release from belowground mass between hairy vetch and cereal rye. However, the pattern of decaying was different for belowground biomass even though the CN ratio of root mass was in line with aboveground litter, which was 17:1 for hairy vetch and 40:1 for cereal rye [
5]. Sun et al. [
43] found distinct traits other than the CN ratio that controls root decomposition where N was not the major driver, but C compounds associated with mycorrhiza were the major factors. They reported a poorer fit of the double exponential model for the root mass remaining data. The parameter estimates vary more with the model fitting procedure for exponential equations than the hyperbolic models [
16]. The reason is that the first-order equation assumes that the rate of mineralization is proportional to the initial size (
MR0 or
NR0) of the mineralizable pool [
27], which might be affected by environmental and management factors and generate more variance. In our study, the belowground initial mineralizable pool was considerably smaller than the aboveground pool, and a significant portion of root mass remained undecomposed after 550 DDD [
5]; thus, exponential models produced an ambiguous estimate of parameters. The evidence of a wide range of parameter estimates of the first-order equation was also reported by Nicolardot et al. [
46] and Talpaz et al. [
47].
The asymptotic approach used by Sievers and Cook [
5] and Singh et al. [
25] in Exp. 1 and 2, respectively, to estimate decomposition and N mineralization rate had higher RMS and SEE than the double exponential and hyperbolic models for both crops. The point of contention was that the single exponential asymptotic model mostly departed from the assumption of normally distributed population, constant variance, and independence of residuals. However, the lignin content of the plant materials leaves a certain amount of highly recalcitrant compound in the late stages of decomposition, despite having a high rate of decomposition, which can typically be addressed by the asymptotic form of equations [
43]. However, use of an asymptote in a single exponential model for a short study period may limit the first-order decomposable pool to a smaller fraction and might give undue weight to the slowly decomposing fraction than the relatively long period. It is difficult to infer the persistence of the recalcitrant fraction and conversion of residue into the soil organic matter based on mass loss and N remaining data [
48,
49]. The addition of C dynamics and microbial growth rate factors in the study would be a step towards the empirical calculation of soil organic matter conversion. However, the current study was able to detect the statistical precision and fitness of the commonly used models in decomposition and N dynamics studies. Finding control mechanisms that fitted hyperbolic function to the root decomposition data would extend the prospect of this study.





{kind=link}
{kind=link}