Effect of Salinity and Nitrogen Sources on the Leaf Quality, Biomass, and Metabolic Responses of Two Ecotypes of Portulaca oleracea


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Biomass
2.2. Measurement of Total Chlorophyll, Total Carotenoids, and Protein Content
2.3. Metabolite Analysis by GC-MS (Gas Chromatography-Mass Spectrometry)
2.3.1. Derivatization
2.3.2. Lipid Analysis
2.3.3. GC-MS Conditions and Analysis
2.3.4. Statistical Analysis
3. Results
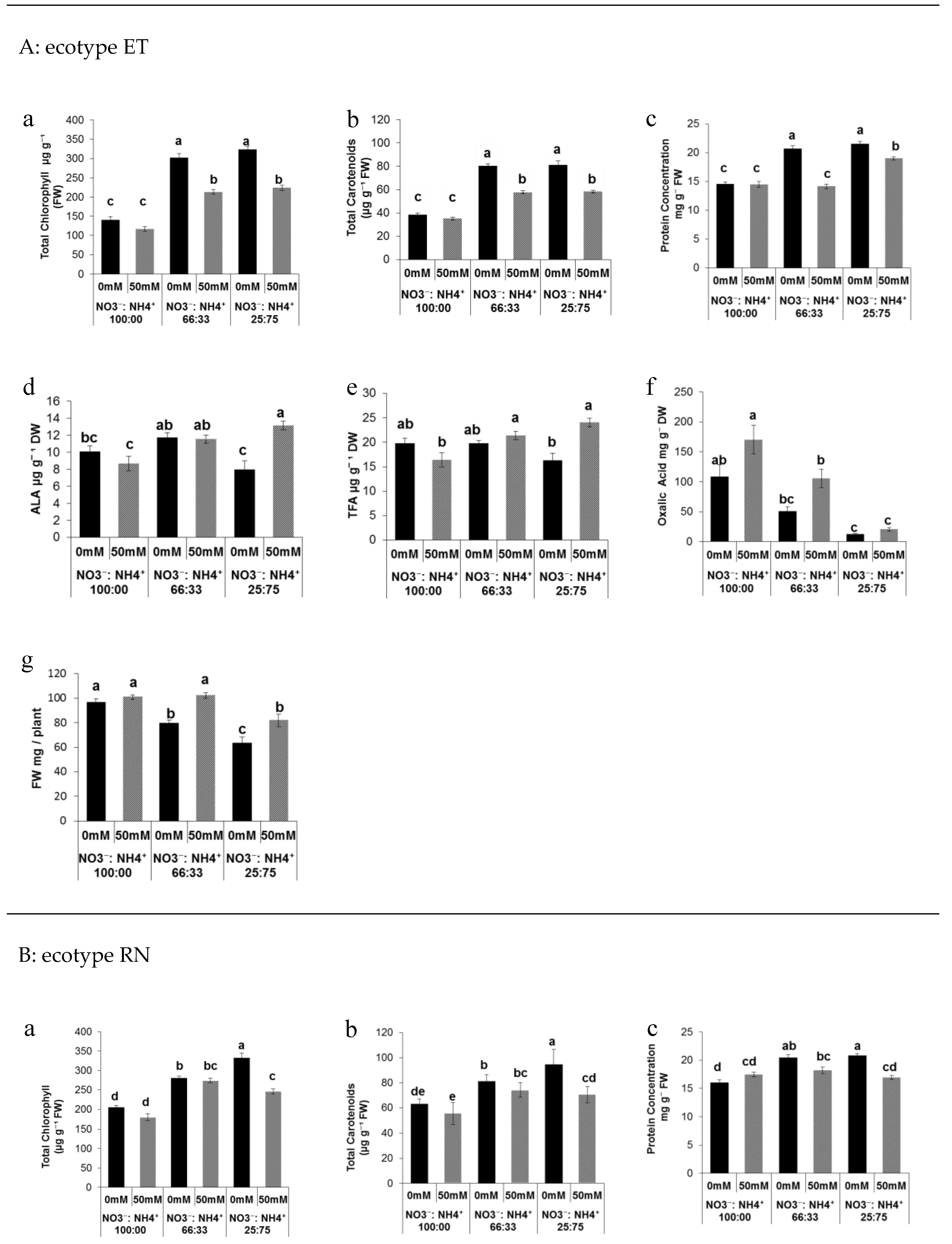
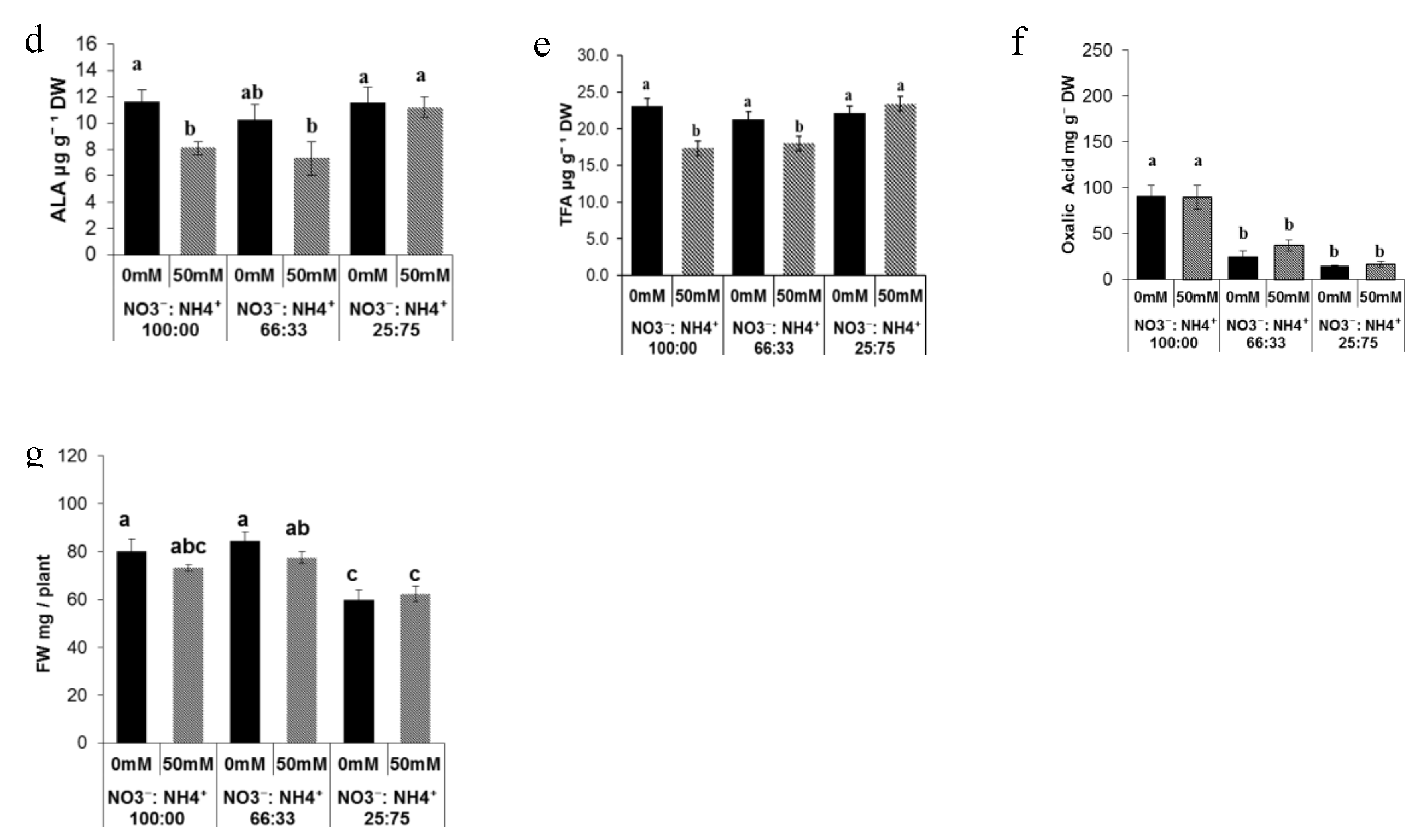
3.1. Effect of Nitrogen Forms and Salinity on the Leave Quality of Portulaca Plants and Biomass Production
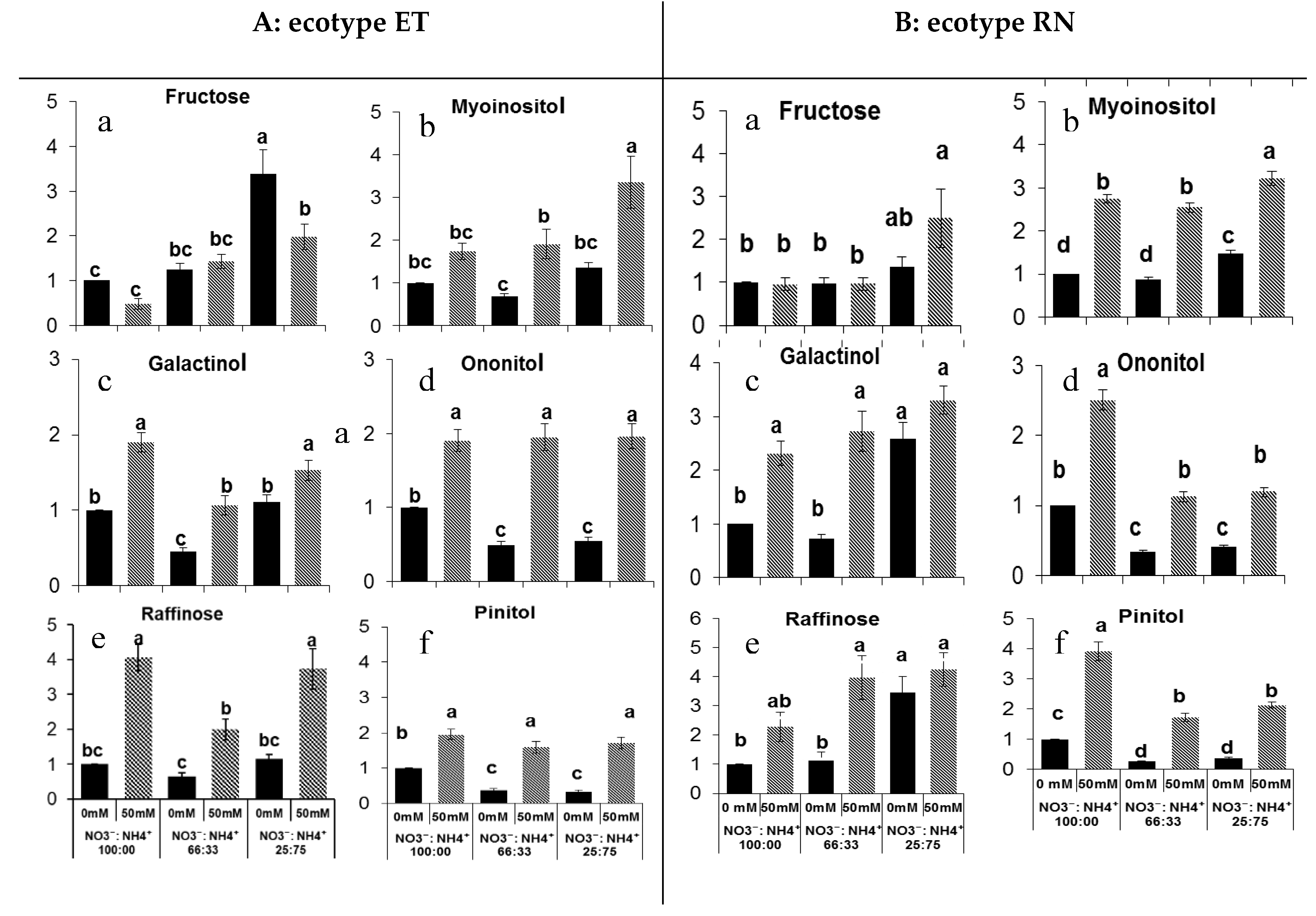
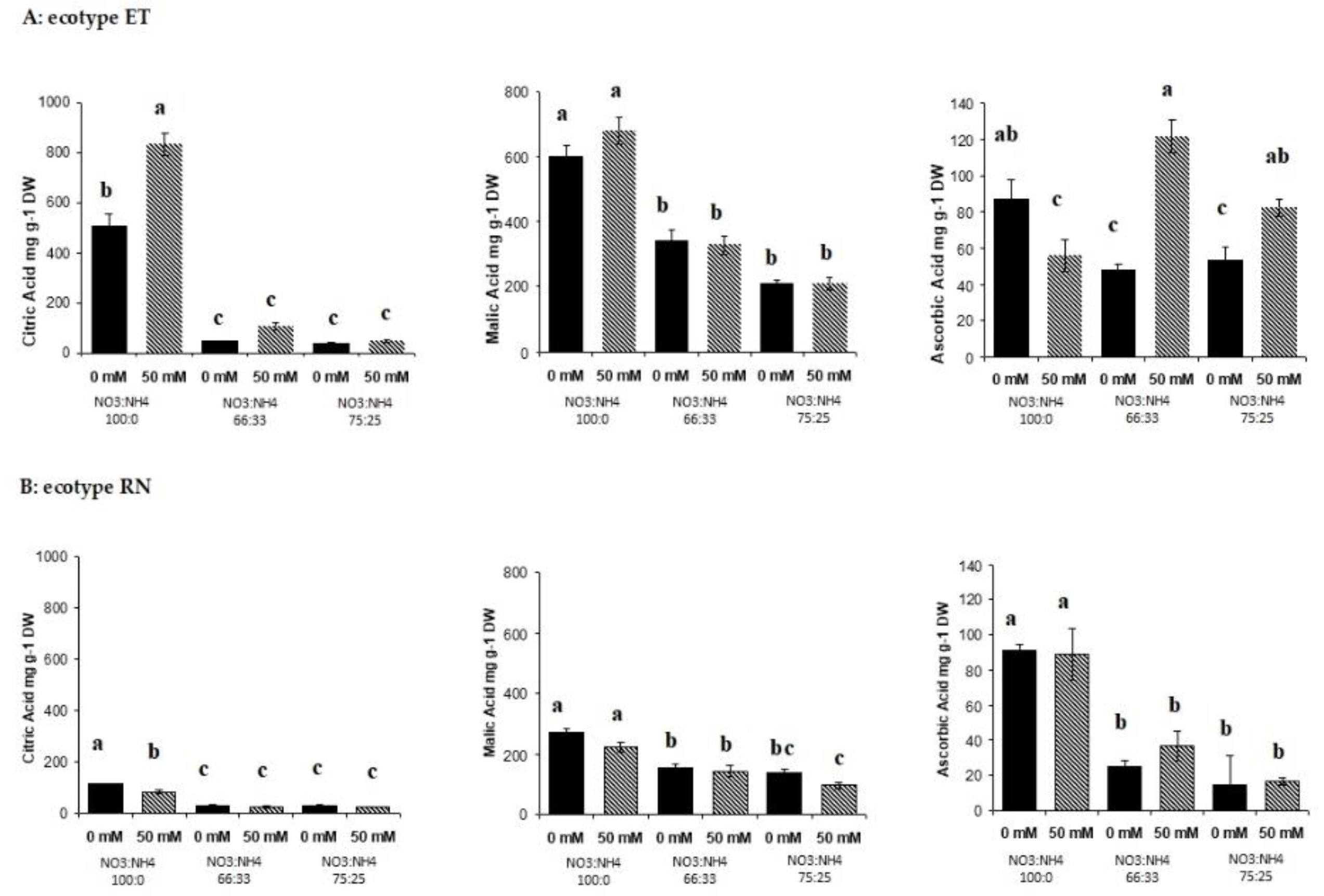
3.2. Sugar Metabolite Changes in Response to Salt and Nitrogen Forms
3.3. Sugar Metabolite Changes in Response to Salt and Nitrogen Forms
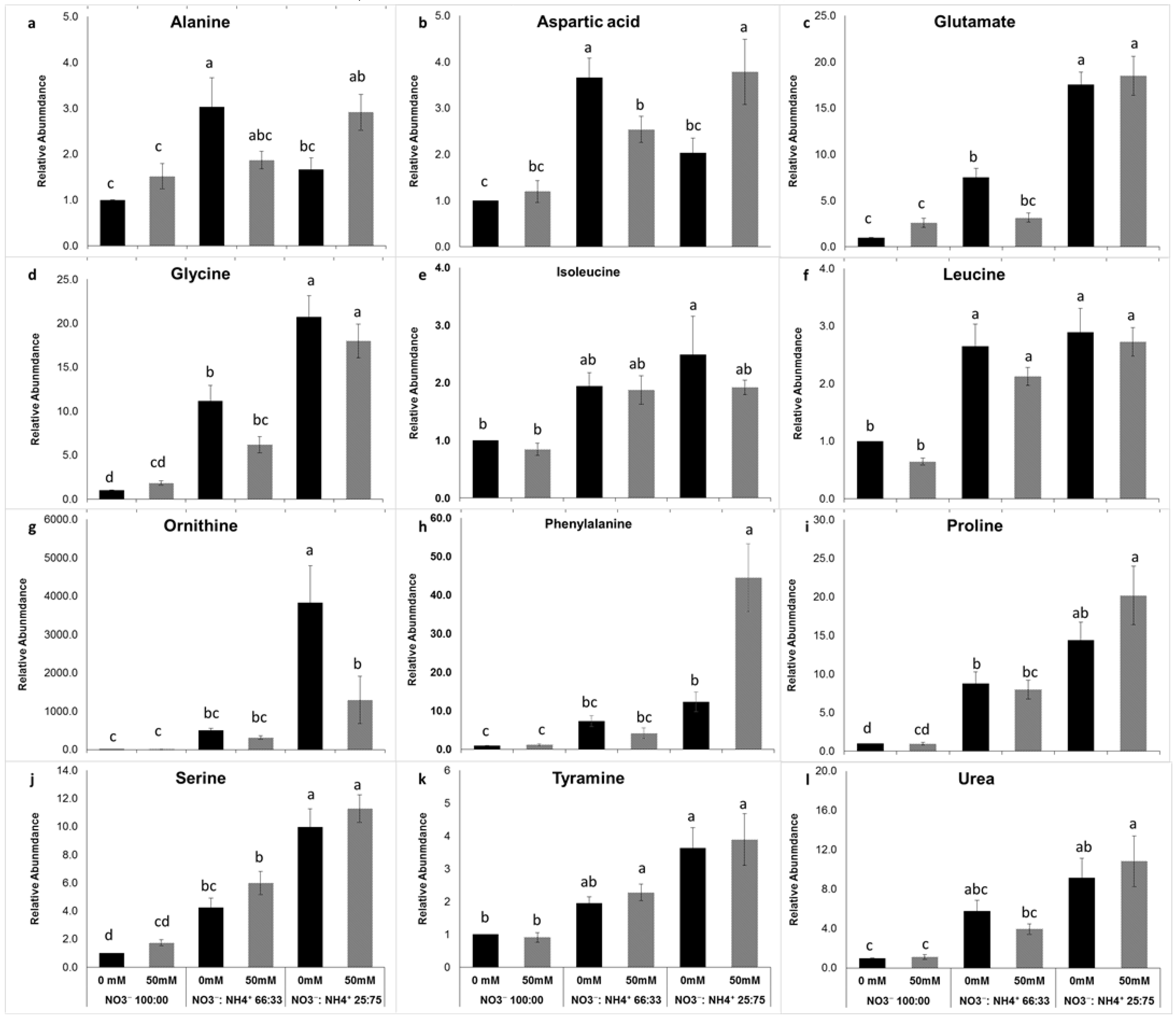
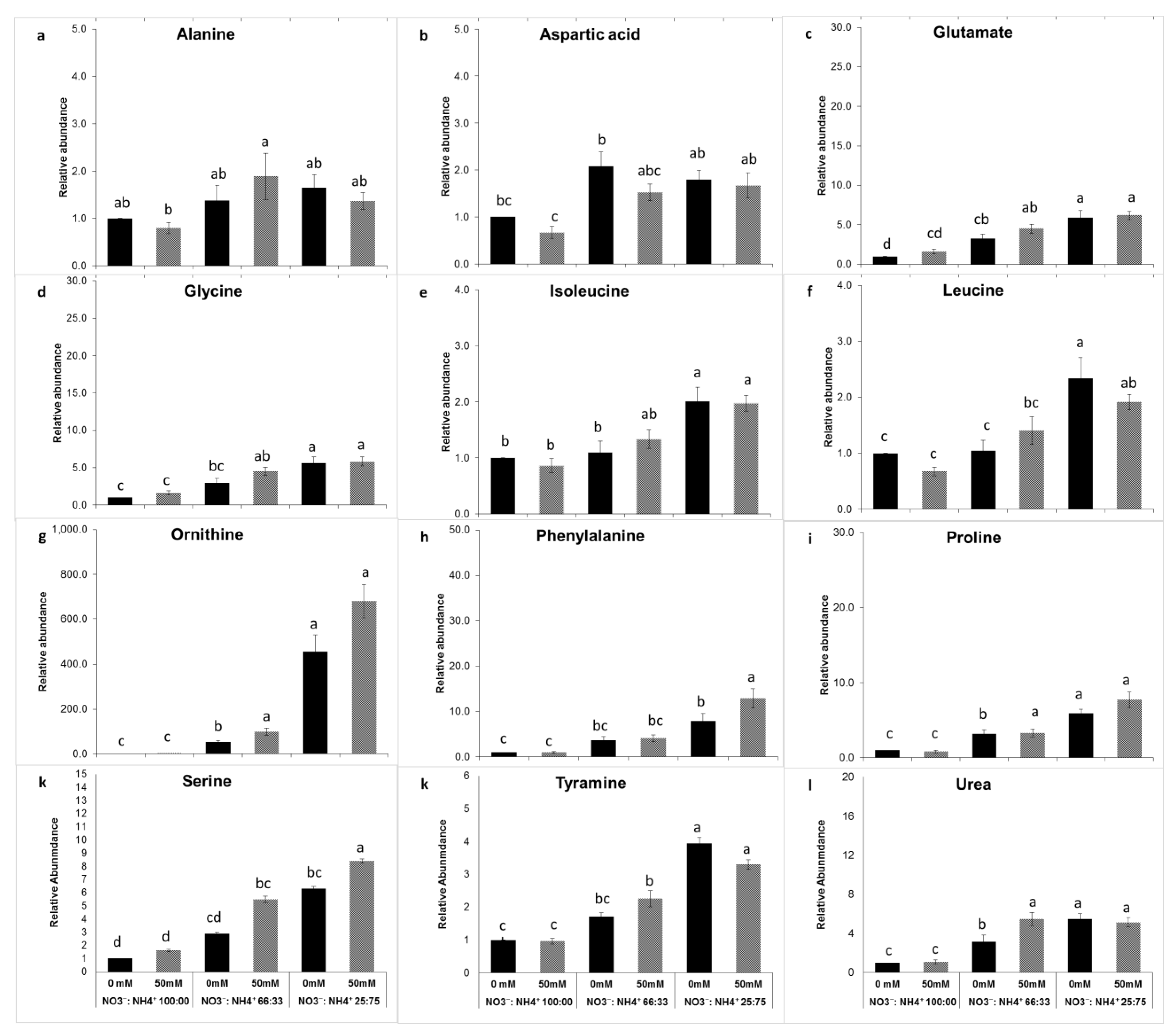
3.4. Free Amino Acid Metabolite Changes in Response to Salt and Nitrogen Forms
4. Discussion and Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chinnusamy, V.; Zhu, J.K. Plant salt tolerance. Topics in current genetics. In Plant Responses to Abiotic Stress; Hirt, K., Shinozaki, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; Volume 4, pp. 241–270. [Google Scholar]
- FAO. FAO Soils Portal. Available online: http://www.fao.org/soils-portal/soilmanagement/management-of-some-problem-soils/salt-affected-soils/moreinformation-on-salt-affected-soils/en/ (accessed on 12 October 2016).
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Neumann, P.M. Inhibition of root growth by salinity stress: Toxicity of an adaptive biophysical response? In Structure and Function of Roots. Development in Plant and Soil Sciences; Baluska, F., Ciamporova, M., Gasparikova, O., Barlow, P.W., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 299–304. [Google Scholar]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. J. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Lieth, H.; Moschenko, M.; Lohmann, M.; Koyro, H.-W.; Hamdy, A. Halophyte uses in different climates I. Ecological and ecophysiological studies. In Progress in Biometeorology; Lieth, H., Ed.; Backhuys Publishers: Leiden, The Netherlands, 1999; pp. 158–258. [Google Scholar]
- Flowers, T.J.; Hajibagheri, M.A.; Clipson, N.J.W. Halophytes. Quart. Rev. Biol. 1986, 61, 313–337. [Google Scholar] [CrossRef]
- Grieve, C.M.; Suarez, D.L. Purslane (Portulaca oleracea L.): A halophytic crop for drainage water reuses systems. Plant Soil 1997, 192, 277–283. [Google Scholar] [CrossRef]
- Ventura, Y.; Eshel, A.; Pasternak, D.; Sagi, M. The development of halophyte-based agriculture: Past and present. Ann. Bot. 2015, 115, 529–540. [Google Scholar] [CrossRef]
- Ventura, Y.; Myrzabayeva, M.; Alikulov, Z.; Cohen, S.; Shemer, Z.; Sagi, M. The importance of iron supply during repetitive harvesting of Aster tripolium. Funct. Plant Biol. 2013, 40, 968–976. [Google Scholar] [CrossRef]
- Feigin, A.; Rylski, I.; Meiri, A.; Shalhevet, J. Response of melon and tomato plants to chloride-nitrate ratios in saline nutrient solutions. J. Plant Nutr. 1987, 10, 1787–1794. [Google Scholar] [CrossRef]
- Palaniswamy, U.R.; Bibble, B.B.; McAvoy, R. Effect of nitrate: Ammonium nitrogen ratio on oxalate levels of purslane. In Trends in New Crops and New Uses; Janick, J., Whipkey, A., Eds.; ASHS Press: Alexandria, VA, USA, 2002; pp. 453–455. [Google Scholar]
- Fontana, E.; Hoeberechts, J.; Nicole, S.; Cros, V.; Palmegiano, B.G.; Peiretti, G.P. Nitrogen concentration and nitrate/ammonium ratio affect yield and change the oxalic acid concentration and fatty acid profile of purslane (Portulaca oleracea L.) grown in a soilless culture system. J. Sci. Food Agric. 2006, 86, 2417–2424. [Google Scholar] [CrossRef]
- Szalai, G.; Dai, N.; Avinoam, D.; Dudai, N.; Barazani, O. Effect of nitrogen source in the fertilizing solution on nutritional quality of three members of the Portulaca oleracea aggregate. J. Sci. Food Agric. 2010, 90, 2039–2045. [Google Scholar] [CrossRef]
- Ahmed, A.K.; Johnson, K.A. The effect of the ammonium:nitrogen ratio, total nitrogen, salinity (NaCl) and calcium on the oxalate levels of Tetragonia tetragonioides Pallas. Kunz. J. Hortic. Sci. Biotechnol. 2000, 75, 533–538. [Google Scholar] [CrossRef]
- Zhang, Y.; Xianyong, L.Y.; Zhang, Y.; Zheng, S.J.; Du, S. Effects of nitrogen levels and nitrate/ammonium ratios on oxalate concentrations of different forms in edible parts of spinach. J. Plant Nutr. 2005, 28, 2011–2025. [Google Scholar] [CrossRef]
- Libert, B.; Franceschi, V.R. Oxalate in crop plants. J. Agric. Food Chem. 1987, 35, 926–938. [Google Scholar] [CrossRef]
- Sagi, M.; Savidov, N.A.; Lvov, N.P.; Lips, S.H. Nitrate reductase and molybdenum cofactor in annual ryegrass as affected by salinity and nitrogen source. Phys. Plant 1997, 99, 546–553. [Google Scholar] [CrossRef]
- Lincoln, S.D.; Black, B. Halogeton poisoning in range cattle. J. Am. Vet. Med. Assoc. 1980, 176, 717–718. [Google Scholar] [PubMed]
- Palaniswamy, U.R.; Bibble, B.B.; McAvoy, R.J. Oxalic acid concentration in purslane (Portulaca oleracea L.) is altered by the stage of harvest and the nitrate to ammonium ratios in hydroponics. Sci. Hortic. 2004, 102, 267–275. [Google Scholar] [CrossRef]
- Carvalho, I.S.; Teixaira, M.; Brodelius, M. Effect of salt stress on purslane and potential health benefits: Oxalic acid and fatty acids profile. Proc. Int. Plant Nut. Coll. XVI 2009. Available online: https://escholarship.org/uc/item/4cc78714 (accessed on 12 October 2016).
- Wintermans, J.F.; De Mots, A. Spectrophotometric characteristics of chlorophylls a and b and their pheophytins in ethanol. Biochim. Biophys. Acta 1965, 109, 448–453. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatography mass spectrometry-based metabolite profiling in plants. Nat. Protoc. 2006, 1, 387–396. [Google Scholar] [CrossRef]
- Brychkova, G.; Yarmolinsky, D.; Batushansky, A.; Grishkevich, V.; Khozin-Goldberg, I.; Fait, A.; Amir, R.; Fluhr, R.; Sagi, M. Sulfite oxidase activity is essential for normal sulfur, nitrogen and carbon metabolism in tomato leaves. Plants 2015, 4, 573–605. [Google Scholar] [CrossRef]
- R. Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-projectorg/ (accessed on 8 July 2019).
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.5. 2017. Available online: https://rdrr.io/cran/factoextra/ (accessed on 8 August 2018).
- Covarrubias, J.I.; Pisi, A.; Rombolà, A.D. Evaluation of sustainable management techniques for preventing iron chlorosis in the grapevine. Aust. J. Grape Wine Res. 2014, 20, 149–159. [Google Scholar] [CrossRef]
- López-Millán, A.; Morales, F.; Abadía, A.; Abadía, J. Changes induced by Fe deficiency and Fe resupply in the organic acid metabolism of sugar beet (Beta vulgaris) leaves. Phys. Plant 2001, 112, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, E.; Orcutt, D. Nutrient deficiency stress and plant growth and development. Chapter 3. Salinity stress. In The Physiology of Plants under Stress—Soil and Biotic Factors; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2000; pp. 43–213. [Google Scholar]
- Xu, N.; Zhang, X.; Fan, X.; Han, L.; Zeng, C. Effect of nitrogen source and concentration on growth rate and fatty acid composition of Ellipsioidion SP. (Eustigmatophyta). J. Appl. Phyol. 2001, 13, 463–469. [Google Scholar] [CrossRef]
- Jin, R.; Wang, Y.; Liu, R.; Gou, J.; Chan, Z. Physiological and Metabolic Changes of Purslane (Portulaca oleracea L.) in Response to Drought, Heat, and Combined Stresses. Front. Plant Sci. 2016, 6, 1123. [Google Scholar] [CrossRef]
- Orthen, B.; Popp, M.; Smirnoff, N. Hydroxyl radical scavenging properties of cyclitols. Proceedings of the Royal Society of Edinburgh B. Biol. Sci. 1994, 2, 269–272. [Google Scholar]
- Bohnert, H.J.; Nelson, D.E.; Jensen, R.G. Adaptations to environmental stresses. Plant Cell 1995, 7, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.E.; Rammesmayer, G.; Bohnert, H.J. Regulation of cell-specific inositol metabolism and transport in plant salinity tolerance. Plant Cell 1998, 10, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Nasir, F.; Batarseh, M.; Abdel-Ghani, A.H.; Jiries, A. Free amino acids content in some halophytes under salinity stress in arid environment, Jordan. Clean Soil Air Water 2010, 38, 592–600. [Google Scholar] [CrossRef]
- Kalamaki, M.S.; Merkouropoulos, G.; Kanellis, A.K. Can ornithine accumulation modulate abiotic stress tolerance n Arabidopsis? Plant Signal. Behav. 2009, 4, 1099–1101. [Google Scholar] [CrossRef]
- Muscolo, A.; Junker, A.; Klukas, C.; Weigelt-Fischer, K.; Riewe, D.; Altmann, T. Phenotypic and metabolic responses to drought and salinity of four contrasting lentil accessions. J. Exp. Bot. 2015, 66, 5467–5480. [Google Scholar] [CrossRef]
- Yang, C.; Chong, J.; Kim, C.; Li, C.; Shi, D.; Wang, D. Osmotic adjustment and ion balance traits of an alkaline resistant halophyte Kochia sieversiana during adaptation to saline and alkaline conditions. Plant Soil 2007, 76, 263–276. [Google Scholar] [CrossRef]
- Smirnoff, N. The function and metabolism of ascorbic acid in plants. Ann. Bot. 1996, 78, 661–669. [Google Scholar] [CrossRef]
- Conklin, P.L.; Barth, C. Ascorbic acid, a familiar small molecule intertwined in the response of plants to ozone, pathogens and the onset of senescence. Plant Cell Environ. 2004, 27, 959–970. [Google Scholar] [CrossRef]
- Hemavathi, C.P.U.; Akula, N.; Young, K.E.; Chun, S.C.; Kim, D.H.; Park, S.W. Enhanced ascorbic acid accumulation in transgenic potato confers tolerance to various abiotic stresses. Biotechnol. Lett. 2010, 32, 321–330. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camalle, M.; Standing, D.; Jitan, M.; Muhaisen, R.; Bader, N.; Bsoul, M.; Ventura, Y.; Soltabayeva, A.; Sagi, M. Effect of Salinity and Nitrogen Sources on the Leaf Quality, Biomass, and Metabolic Responses of Two Ecotypes of Portulaca oleracea. Agronomy 2020, 10, 656. https://doi.org/10.3390/agronomy10050656
Camalle M, Standing D, Jitan M, Muhaisen R, Bader N, Bsoul M, Ventura Y, Soltabayeva A, Sagi M. Effect of Salinity and Nitrogen Sources on the Leaf Quality, Biomass, and Metabolic Responses of Two Ecotypes of Portulaca oleracea. Agronomy. 2020; 10(5):656. https://doi.org/10.3390/agronomy10050656
Chicago/Turabian StyleCamalle, Maria, Dominic Standing, Mohammed Jitan, Rana Muhaisen, Nidal Bader, Majed Bsoul, Yvonne Ventura, Aigerim Soltabayeva, and Moshe Sagi. 2020. "Effect of Salinity and Nitrogen Sources on the Leaf Quality, Biomass, and Metabolic Responses of Two Ecotypes of Portulaca oleracea" Agronomy 10, no. 5: 656. https://doi.org/10.3390/agronomy10050656
APA StyleCamalle, M., Standing, D., Jitan, M., Muhaisen, R., Bader, N., Bsoul, M., Ventura, Y., Soltabayeva, A., & Sagi, M. (2020). Effect of Salinity and Nitrogen Sources on the Leaf Quality, Biomass, and Metabolic Responses of Two Ecotypes of Portulaca oleracea. Agronomy, 10(5), 656. https://doi.org/10.3390/agronomy10050656