Soybean Seed Vigor: Uniformity and Growth as Key Factors to Improve Yield

 , ,
, ,  , and
, and
Abstract
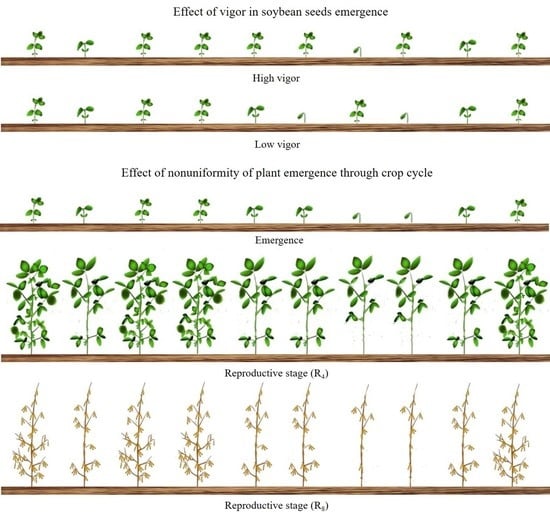
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
2.3. Procedure
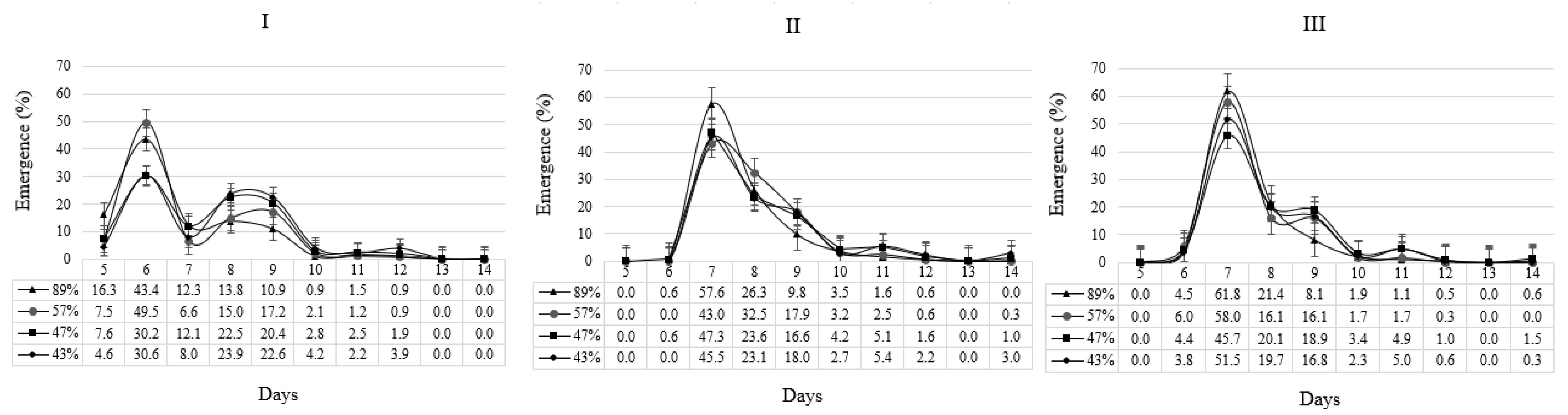
2.3.1. Emergence Speed, Plant Stand, and Uniformity Index
2.3.2. Plant Growth and Development
2.3.3. Nodulation
2.3.4. Grain Yield Components
2.4. Statistical Analysis
3. Results
3.1. Seedling Emergence, Plant Stand, and Uniformity
3.2. Soybean Growth and Development
3.3. Nodulation
3.4. Shoot, Root, and Stem
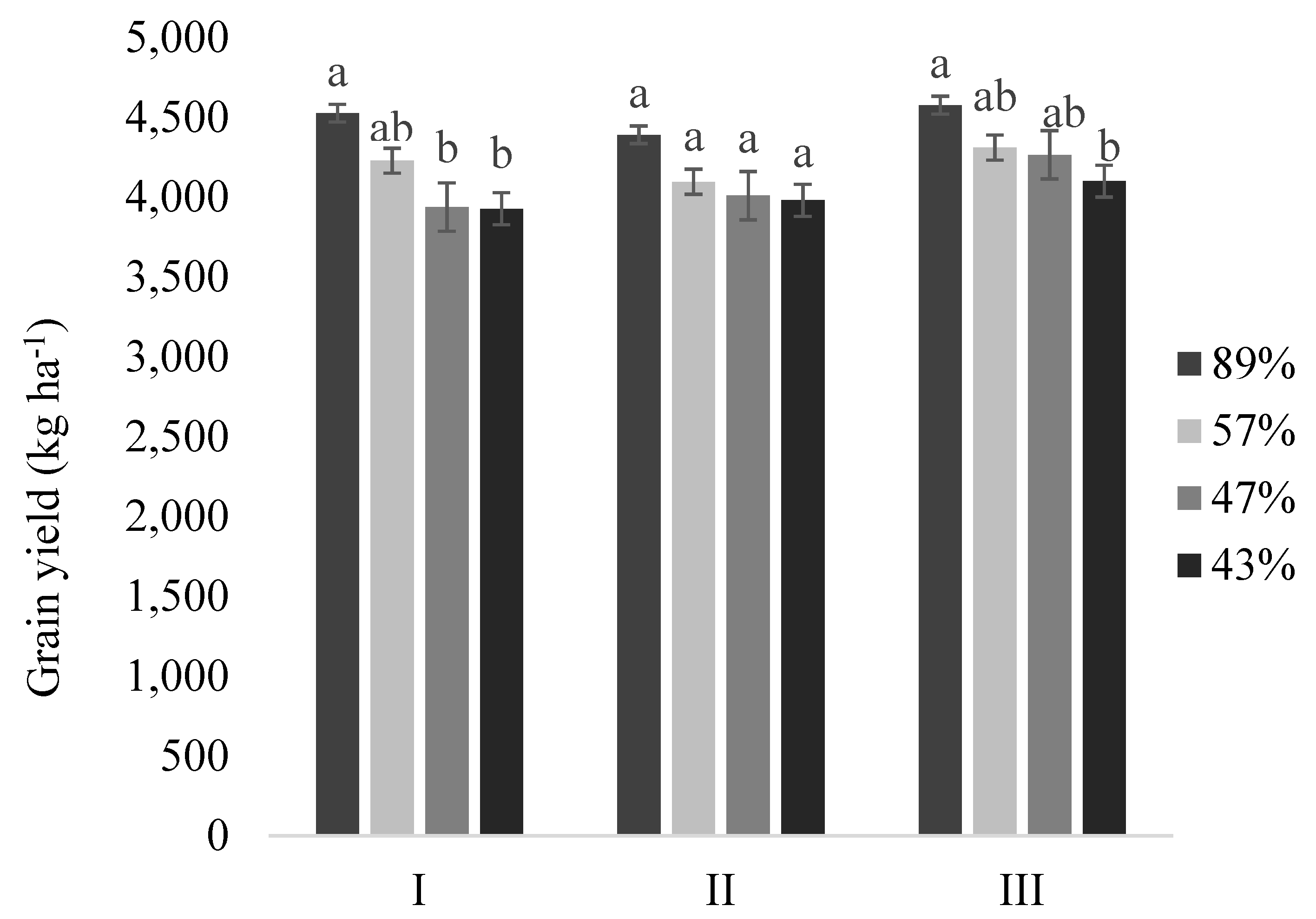
3.5. Yield Components
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Friedman, M.; Brandon, D.L. Nutritional and health benefits of soy proteins. J. Agric Food Chem. 2001, 49, 1069–1086. [Google Scholar] [CrossRef] [PubMed]
- Grau, R.; Gasparri, N.I.; Aide, T.M. Agriculture expansion and deforestation in seasonally dry forests of north-west Argentina. Environ. Conserv. 2005, 32, 140–148. [Google Scholar] [CrossRef]
- Ebone, L.A.; Caverzan, A.; Chavarria, G. Physiologic alterations in orthodox seeds due to deterioration processes. Plant Physiol. Biochem. 2019, 145, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Wimalasekera, R. Role of Seed Quality in Improving Crop Yields. In Production and Global Environmental Issues; Hakeem, K., Ed.; Springer: Berlin, Germany, 2015; pp. 153–168. [Google Scholar]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigour and crop establishment: Extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, W.Q.; Liu, S.J.; Moller, I.M.; Song, S.Q. Proteome analysis of poplar seed vigor. PLoS ONE 2015, 10, e0132509. [Google Scholar] [CrossRef]
- Matthews, S.; Khajeh, H.M. Mean germination time as an indicator of emergence performance in soil of seed lots of maize (Zea mays). Seed Sci Technol. 2006, 24, 339–347. [Google Scholar] [CrossRef]
- Tao, L.; Rui, L.; Xiuliang, J.; Jinfeng, D.; Xinkai, Z.; Chengming, S.; Guo, W. Evaluation of seed emergence uniformity of mechanically sown wheat with UAV RGB imagery. Remote Sens. 2017, 9, 1241. [Google Scholar] [CrossRef]
- Masino, A.; Rugeroni, P.; Borrás, L.; Rotundo, J.L. Spatial and temporal plant-to-plant variability effects on soybean yield. Eur. J. Agron. 2018, 98, 14–24. [Google Scholar] [CrossRef]
- Pinthus, M.J.; Kimel, U. Speed of germination as a criterion of seed vigor in soybeans. Crop Sci. 1979, 19, 291–292. [Google Scholar] [CrossRef]
- Krzyzanowski, F.C.; Vieira, R.D.; França-Neto, J.B. Vigor de Sementes: Conceitos e Testes, 1st ed.; Associação Brasileira de Tecnologia de Sementes-ABRATES: Londrina, Brazil, 1999. [Google Scholar]
- Brasil—Ministério da Agricultura, Pecuária e Abastecimento. Regras Para Análise de Sementes, 1st ed.; Ministério da Agricultura, Pecuária e Abastecimento, Secretária de Defesa Agropecuária: Brasília, Brazil, 2009; pp. 147–220.
- Caverzan, A.; Giacomin, R.; Müller, M.; Biazus, C.; Lângaro, N.C.; Chavarria, G.L. How does seed vigor affect soybean yield components? Agron J. 2018, 110, 1318–1327. [Google Scholar] [CrossRef]
- Fehr, W.R.; Caviness, C.E. Stages of Soybean Development; Spec. Rep. 80; Iowa State University of Science and Technology: Ames, IA, USA, 1977; pp. 2–11. [Google Scholar]
- Egli, D.B.; Rucker, M. Seed vigor and the uniformity of emergence of corn seedlings. Crop Sci. 2012, 52, 2774–2782. [Google Scholar] [CrossRef]
- Marcos-Filho, J. Seed vigor testing: An overview of the past, present and future perspective. Sci. Agri. 2015, 72, 363–374. [Google Scholar] [CrossRef]
- Park, S.E.; Benjamin, L.R.; Watkinson, A.R. The theory and application of plant competition models: An agronomic perspective. Ann Bot. 2003, 92, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Higley, L.G.; Pedigo, L.P. Soybean growth responses and intraspecific competition from simulated seed corn maggot injury. Agron J. 1990, 82, 1057–1063. [Google Scholar] [CrossRef]
- Green-Tracewicz, E.; Page, E.R.; Swanton, C.J. Shade avoidance in soybean reduces branching and increases plant-to-plant variability in biomass and yield per plant. Weed Sci. 2011, 59, 43–49. [Google Scholar] [CrossRef]
- Adegbuyi, E.; Burris, J.S. Field criteria used in determining the vigor of seed corn (Zea mays L.) as influenced by drying injury. J. Agron Crop. Sci. 1988, 161, 171–177. [Google Scholar] [CrossRef]
- TeKrony, D.M.; Egli, D.B. Relationship of seed vigor to crop yield: A review. Crop Sci. 1991, 31, 816–822. [Google Scholar] [CrossRef]
- Müller, M.; Rakocevic, M.; Caverzan, A.; Chavarria, G. Grain yield differences of soybean cultivars due to solar radiation interception. Am. J. Plant Sci. 2017, 8, 2795–2810. [Google Scholar] [CrossRef]
- Bullock, D.; Khan, S.; Rayburn, A. Soybean yield response to narrow rows is largely due to enhanced early growth. Crop Sci. 1998, 38, 1011–1016. [Google Scholar] [CrossRef]
- Hardy, R.W.F.; Havelka, U.D. Photosynthate as a Major Factor Limiting Nitrogen Fixation by Field Grown Legumes with Emphasis on Soybeans. In Symbiotic Nitrogen Fixation in Plant; Nutman, P.S., Ed.; Cambridge University Press: Cambrigde, UK, 1976; pp. 421–439. [Google Scholar]
- Finn, G.A.; Brun, W.A. Effect of atmospheric CO(2) enrichment on growth, nonstructural carbohydrate content, and root nodule activity in soybean. Plant Physiol. 1982, 69, 327–331. [Google Scholar] [CrossRef]
- Hartmann-Filho, C.P.; Goneli, A.L.D.; Masetto, T.E.; Martins, E.A.S.; Oba, G.C. The effect of drying temperatures and storage of seeds on the growth of soybean seedlings. J. Seed Sci. 2016, 38, 287–295. [Google Scholar] [CrossRef]
- Singh, J.; Paroha, S.; Mishra, R.P. Effect of storage on germination and viability of soybean (Glycine max) and niger (Guizotia abyssinica) seeds. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 484–491. [Google Scholar] [CrossRef]
- Engels, C. Effect of root and shoot meristem temperature on shoot to root dry matter partitioning and the internal concentrations of nitrogen and carbohydrates in maize and wheat. Ann Bot. 1994, 73, 211–219. [Google Scholar] [CrossRef]
- Weraduwage, S.M.; Chen, J.; Anozie, F.C.; Morales, A.; Weise, S.E.; Sharkey, T.D. The relationship between leaf area growth and biomass accumulation in Arabidopsis thaliana. Front Plant Sci. 2015, 6, 167. [Google Scholar] [CrossRef]
- Wu, Y.; Gong, W.; Yang, W. Shade inhibits leaf size by controlling cell proliferation and enlargement in soybean. Sci Rep. 2017, 7, 9259. [Google Scholar] [CrossRef]
- Wiebold, W.J.D.; Ashley, A.; Boerma, H.R. Reproductive abscission levels and patterns for eleven determinate soybean cultivars. Agron J. 1981, 73, 43–46. [Google Scholar] [CrossRef]
- Egli, D.B. Flowering, pod set and reproductive success in soya bean. J. Agron Crop Sci. 2005, 191, 283–291. [Google Scholar] [CrossRef]
- Nico, M.; Mantese, A.I.; Miralles, D.J.; Kantolic, A.G. Soybean fruit development and set at the node level under combined photoperiod and radiation conditions. J. Exp Bot. 2016, 67, 365–377. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Yendrek, C.R.; Skoneczka, J.A.; Long, S.P. Accelerating yield potential in soybean: Potential targets for biotechnological improvement. Plant Cell Environ. 2012, 35, 38–52. [Google Scholar] [CrossRef]
- Ribeiro, A.B.M.; Bruzi, A.T.; Zuffo, A.M.; Zambiazzi, E.V.; Soares, I.O.; Vilela, N.J.D.; Pereira, J.L.A.R.; Moreira, S.G. Productive performance of soybean cultivars grown in different plant densities. Cien. Rural. 2017, 47, e20160928. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vigor | Emergence Speed (Days) | Uniformity Index (%) | ||||
|---|---|---|---|---|---|---|
| Site I | Site II | Site III | Site I | Site II | Site III | |
| 89% | 6.8 a | 7.7 a | 7.6 a | 70.6 aB | 82.5 aA | 86.3 aA |
| 57% | 7.0 a | 8.0 ab | 7.7 a | 61.5 aB | 73.8 abA | 80.9 abA |
| 47% | 7.5 b | 8.1 b | 8.0 ab | 48.3 bB | 70.5 bA | 76.7 abA |
| 43% | 7.8 b | 8.2 b | 8.1 b | 38.6 bB | 69.8 bA | 72.1 bA |
| CV (%) | 3.1 | 8.9 | ||||
| First Trifoliate Area (cm2) | Leaf Area V4 (cm2) | Leaf Area R2 (cm2) | Nodules Dry Mass (g) | Number of Nodules | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EDAS | I | II | III | I | II | III | I | II | III | I | II | III | I | II | III |
| 5 | 30.4 a | 1254 a | 2878 a | 0.298 a | 61.6 a | ||||||||||
| 6 | 27.3 a | 45.7 a | 34.8 a | 1103 ab | 1138 a | 2534 a | 3345 abc | 2573 ab | 0.276 ab | 0.1334 ab | 53.4 a | 41 a | |||
| 7 | 26.1 ab | 32.5 b | 30 b | 950 bc | 833 a | 880 b | 2491 ab | 2501 a | 2483 a | 0.177 cd | 0.0905 a | 0.1640 a | 41.1 ab | 38 a | 44.2 a |
| 8 | 24.1 b | 28.4 c | 27.4 c | 904 cd | 726 b | 802 b | 2486 ab | 2140 cd | 2095 bc | 0.259 abc | 0.0939 a | 0.1442 ab | 54.7 a | 38.2 a | 43.4 a |
| 9 | 20.5 c | 24.1 d | 24.5 cd | 720 cde | 597 bc | 596 c | 1927 bc | 1947 bcd | 1849 cd | 0.2439 bc | 0.0936 a | 0.0944 b | 51 ab | 38.3 a | 35.4 ab |
| 10 | 16.5 cd | 23.2 d | 17.5 de | 570 def | 570 abc | 457 bc | 1062 bc | 1220 d | 1624 bcde | 0.195 abcd | 0.0502 a | 0.0787 ab | 49.4 ab | 30 a | 23.5 ab |
| 11 | 9.6 d | 17 e | 16.1 e | 309 ef | 283c | 444 c | 918 c | 1118 d | 1146 de | 0.077 bcd | 0.0392 a | 0.0333 ab | 21.6 ab | 16.5 a | 9.5 b |
| 12 | 9.6 d | 7.7 e | 10.5 ef | 190 f | 300 c | 580 e | 0.048 d | 0.0379 ab | 14 b | 30 ab | |||||
| 14 | 10.5 e | 4.9 f | 266 c | 226 c | 421 cde | 0.0395 a | 0.0204 ab | 17.3 a | 10.5 ab | ||||||
| CSD | 5.5 | 4.6 | 5.2 | 263 | 207 | 223 | 802 | 725 | 627 | 0.11 | 0.07 | 0.08 | 24.9 | 20.4 | 18.1 |
| Shoot Dry Mass (g) | Root Dry Mass (g) | SDM/TDM | Shoot Length (cm) | Root Length (cm) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EDAS | I | II | III | I | II | III | I | II | III | I | II | III | I | II | III |
| 5 | 8.69 a | 1.93 a | 0.82 a | 32.1 a | 16.3 a | ||||||||||
| 6 | 8.15 a | 8.05 a | 1.82 a | 1.39 a | 0.81 a | 0.83 a | 31.1 ab | 28.8 a | 16.5 a | 10.6 a | |||||
| 7 | 7.20 ab | 6.10 a | 6.65 ab | 1.68 ab | 1.30 a | 1.34 a | 0.81 a | 0.8 a | 0.82 a | 30.7 ab | 26.2 a | 26.7 abc | 13 a | 10.3 a | 10.8 a |
| 8 | 6.66 bc | 5.67 a | 5.91 bc | 1.55 abc | 1.20 a | 1.22 a | 0.79 ab | 0.81 a | 0.82 ab | 29 ab | 26.3 a | 27.0 ab | 15.3 a | 10.5 a | 9.5 a |
| 9 | 5.32 cd | 5.54 a | 4.58 cd | 1.46 bc | 1.12 a | 1.22 a | 0.78 b | 0.83 a | 0.77 c | 28.2 b | 25 a | 24.8 cd | 15 a | 9.7 a | 9.3 a |
| 10 | 4.78 bcde | 4.58 a | 3.36 cd | 1.44 abc | 0.83 a | 1 a | 0.76 b | 0.84 a | 0.80 abc | 27.2 abc | 25 a | 24 abcd | 16.8 a | 9 a | 9 a |
| 11 | 2.50 de | 3.47 a | 3.31 bcd | 0.79 cd | 0.87 a | 1.01 a | 0.76 b | 0.79 a | 0.77 abc | 24.3 abc | 25 a | 23.7 abcd | 15 a | 8 a | 8.5 a |
| 12 | 1.53 e | 2.31 cd | 0.42 d | 0.99 a | 0.76 b | 0.67 bcd | 18.7 c | 21 bcd | 9 a | 7.5 a | |||||
| 14 | 3.36 a | 1.36 d | 0.71 a | 0.98 a | 0.83 a | 0.57 d | 21.3 a | 20 d | 7.3 a | 7.5 a | |||||
| CSD | 1.8 | 2.0 | 1.9 | 0.45 | 0.4 | 0.42 | 0.03 | 0.08 | 0.06 | 4.6 | 3.2 | 3 | 6 | 3.2 | 3.2 |
| Stem Diameter (cm) | Shoot Length (cm) | ALI (cm) | Total Number of Nodes | Number of Fertile Nodes | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EDAS | I | II | III | I | II | III | I | II | III | I | II | III | I | II | III |
| 5 | 0.91 a | 96.7 a | 4.70 b | 20.6 a | 17.4 a | ||||||||||
| 6 | 0.85 a | 1 abc | 1 a | 97.1 a | 94.4 a | 4.72 b | 4.91 a | 20.6 a | 19.2 a | 17.1 a | 16.6 a | ||||
| 7 | 0.85 ab | 0.90 a | 0.96 a | 97.7 a | 94.4 a | 94.3 a | 4.76 b | 4.87 b | 5.62 a | 20.6 a | 19.4 a | 19.1 a | 17 a | 15.8 a | 15.2 ab |
| 8 | 0.81 ab | 0.88 a | 0.89 ab | 97 a | 92 ab | 94.6 a | 4.84 ab | 4.89 b | 5.04 a | 20.1 a | 18.8 ab | 18.8 a | 16.2 ab | 14.9 a | 14.3 abc |
| 9 | 0.72 bc | 0.73 c | 0.72 c | 96.3 a | 91.1 bc | 95.4 a | 5.04 ab | 5.02 ab | 5.19 a | 19.5 ab | 18.2 bc | 18.4 ab | 15 b | 13.3 b | 13 cd |
| 10 | 0.46 cd | 0.6 bc | 0.66 abc | 95 a | 90.8 abc | 94.1 a | 5.18 ab | 5.38 a | 5.39 a | 18.4 ab | 17 c | 17.5 ab | 13.6 bc | 12 b | 11.6 d |
| 11 | 0.4 d | 0.62 bc | 0.62 bc | 93.3 a | 89.3 abc | 93.4 a | 5.47 a | 5.42 a | 5.69 a | 17.1 b | 16.5 c | 16.4 b | 11 c | 11.1 b | 10.1 d |
| 12 | 0.4 c | 82.6 c | 93 ab | 5.08 ab | 6.2 a | 16.3 c | 15 ab | 11 b | 9 bcd | ||||||
| 14 | 0.4 abc | 71 b | 5.46 a | 13 b | 7 d | ||||||||||
| CSD | 0.17 | 0.19 | 0.19 | 6 | 5.1 | 6.8 | 0.5 | 0.37 | 2.3 | 2 | 1.4 | 1.8 | 2.3 | 2 | 2.2 |
| Number of Pods | Grains for Pod | Number of Grains | Thousand-Seed Weight (g) | Grain Mass (g) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EDAS | I | II | III | I | II | III | I | II | III | I | II | III | I | II | III |
| 5 | 80.9 a | 2.41 a | 194.9 a | 166.3 a | 32 a | ||||||||||
| 6 | 72.6 a | 71.9 a | 2.42 a | 2.49 a | 175.2 a | 179.5 a | 164.3 a | 168.8 bc | 28.8 a | 30.2 a | |||||
| 7 | 69.8 ab | 65.6 a | 62.2 a | 2.39 a | 2.44 a | 2.51 a | 166.7 ab | 160 a | 156.2 a | 165.1 a | 168.7 b | 172.8 c | 27.4 ab | 26.8 a | 26.9 ab |
| 8 | 59.7 bc | 56.4 b | 52.7 b | 2.42 a | 2.46 a | 3.12 a | 144.5 bc | 138.8 b | 135.7 b | 170.6 a | 170.2 b | 181.9 abc | 24.6 bc | 23.4 b | 24.1 abc |
| 9 | 54.2 c | 46.2 c | 42.6 c | 2.47 a | 2.46 a | 2.53 a | 133.7 c | 114.3 c | 108.3 c | 165.7 a | 180.1 ab | 183.3 abc | 22 c | 20.5 b | 20.1 c |
| 10 | 40.6 cd | 39.2 c | 39.7 bc | 2.48 a | 2.41 a | 2.54 a | 100.6 cd | 95.3 c | 101.2 bc | 166.8 a | 201.7 a | 179.7 abc | 16.8 cd | 19.1 b | 18 bc |
| 11 | 25.8 d | 36.3 c | 32.8 c | 2.41 a | 2.52 a | 2.52 a | 62.5 d | 92 c | 83.3 c | 161.4 a | 184.2 ab | 227.2 a | 9.9 cd | 16.7 b | 18.1 c |
| 12 | 33 c | 18 bc | 2.53 a | 2.55 a | 83.7 c | 46 bc | 180.5 ab | 285.7 abc | 15.2 b | 13.1 abc | |||||
| 14 | 16 bc | 2.43 a | 39 bc | 216.2 abc | 8.4 abc | ||||||||||
| CSD | 15.4 | 13.6 | 13.8 | 0.16 | 0.15 | 1.8 | 36.2 | 31.8 | 34.7 | 16 | 21 | 37.2 | 6.1 | 5.4 | 7.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebone, L.A.; Caverzan, A.; Tagliari, A.; Chiomento, J.L.T.; Silveira, D.C.; Chavarria, G. Soybean Seed Vigor: Uniformity and Growth as Key Factors to Improve Yield. Agronomy 2020, 10, 545. https://doi.org/10.3390/agronomy10040545
Ebone LA, Caverzan A, Tagliari A, Chiomento JLT, Silveira DC, Chavarria G. Soybean Seed Vigor: Uniformity and Growth as Key Factors to Improve Yield. Agronomy. 2020; 10(4):545. https://doi.org/10.3390/agronomy10040545
Chicago/Turabian StyleEbone, Luciano Antônio, Andréia Caverzan, Alberto Tagliari, José Luís Trevizan Chiomento, Diógenes Cecchin Silveira, and Geraldo Chavarria. 2020. "Soybean Seed Vigor: Uniformity and Growth as Key Factors to Improve Yield" Agronomy 10, no. 4: 545. https://doi.org/10.3390/agronomy10040545
APA StyleEbone, L. A., Caverzan, A., Tagliari, A., Chiomento, J. L. T., Silveira, D. C., & Chavarria, G. (2020). Soybean Seed Vigor: Uniformity and Growth as Key Factors to Improve Yield. Agronomy, 10(4), 545. https://doi.org/10.3390/agronomy10040545