Characterization and Grouping of All Primary Branches at Various Positions on a Rice Panicle Based on Grain Growth Dynamics


 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site and Cultivar Details
2.2. Experimental Design and Crop Management
2.3. Sampling and Measurements
2.4. Mathematical Simulation of Grain Growth
2.5. Statistical Analysis
3. Results
3.1. Dynamics of Dry-Matter Accumulation for Primary-Branch Grains
3.2. Mathematical Simulation of Grain Growth
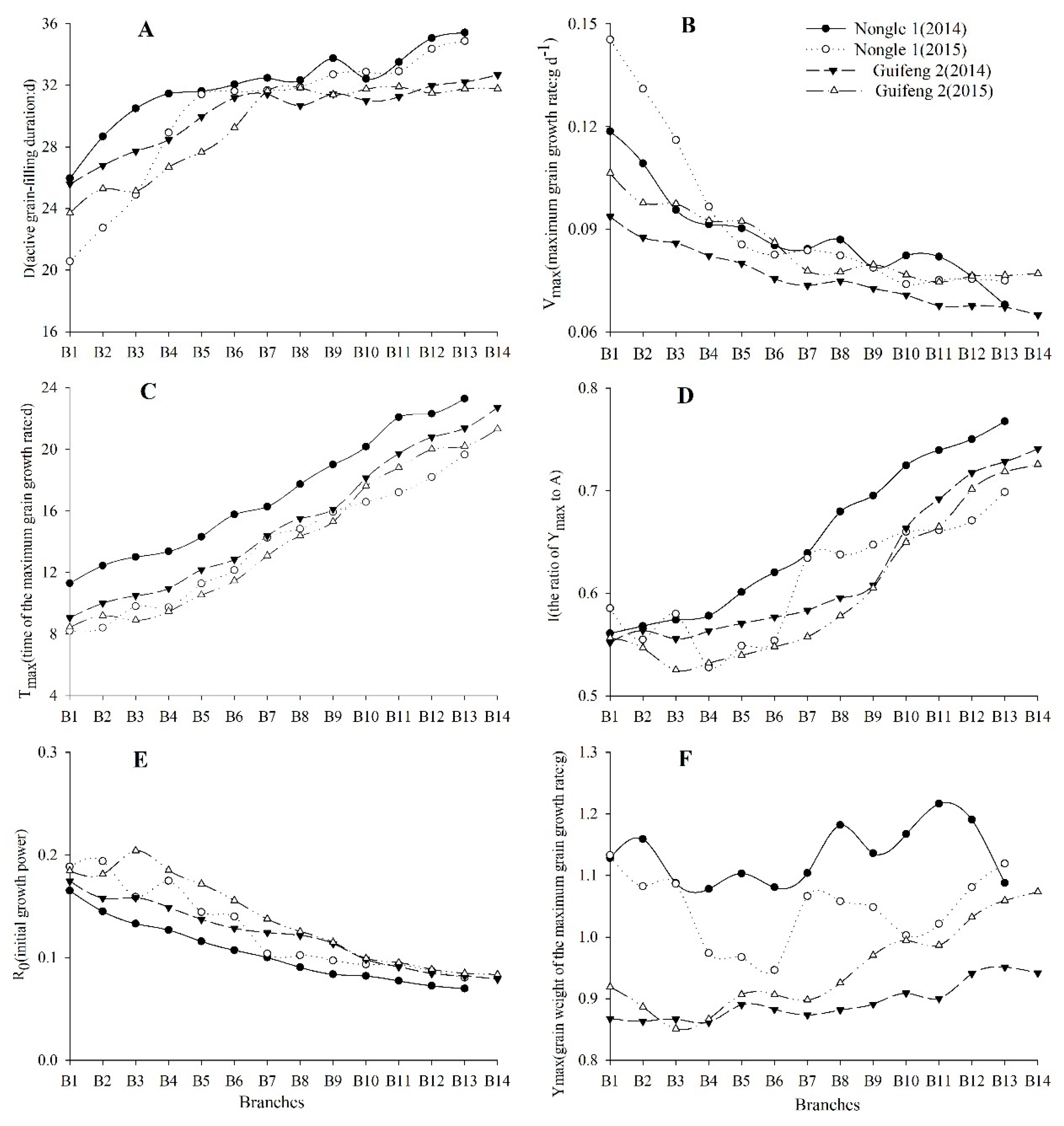
3.3. Characteristics of Growth Parameters
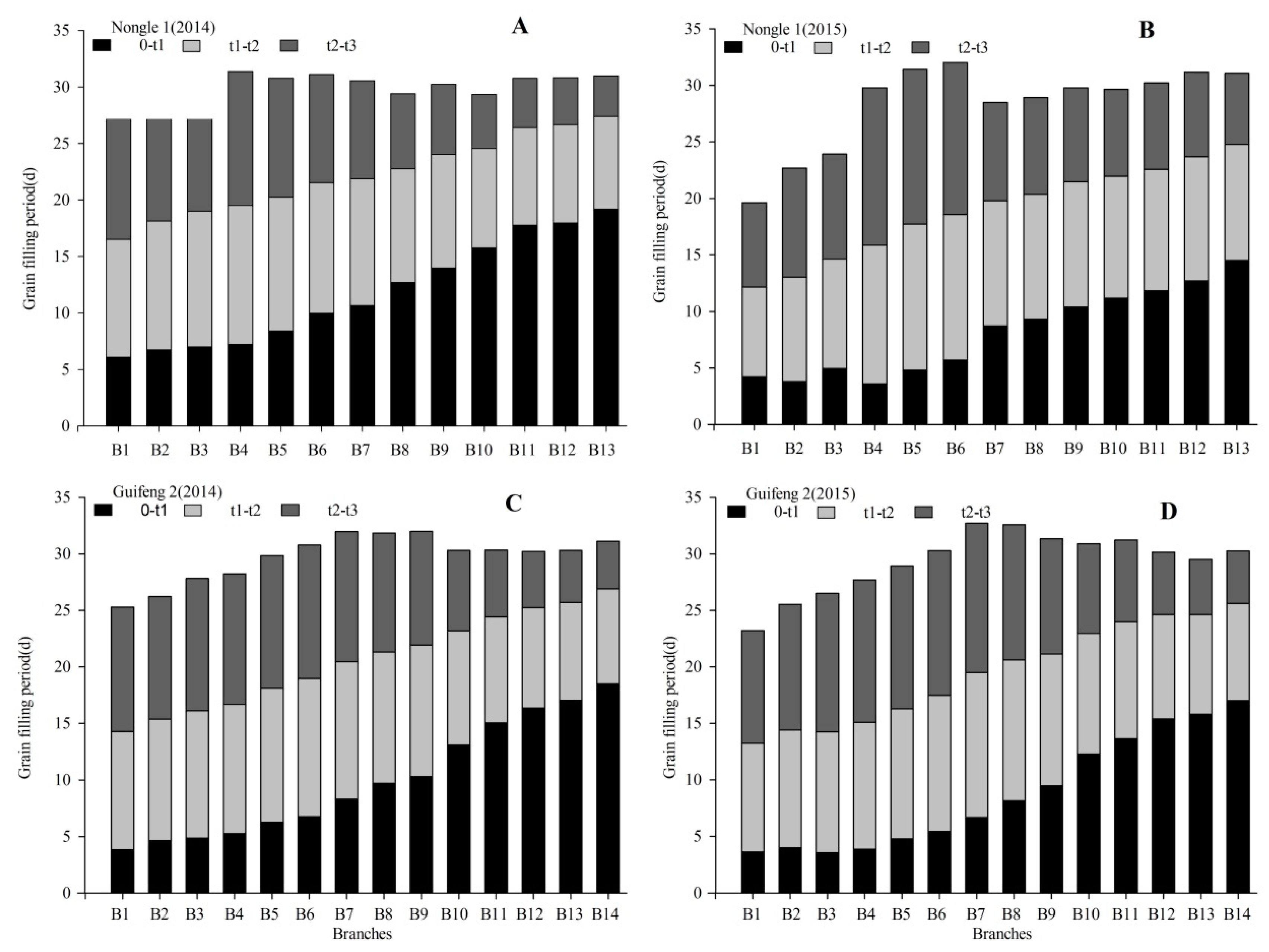
3.4. Division of Growth Stages
3.5. Correlation of Dry-Matter Accumulation between Primary-Branch Grains
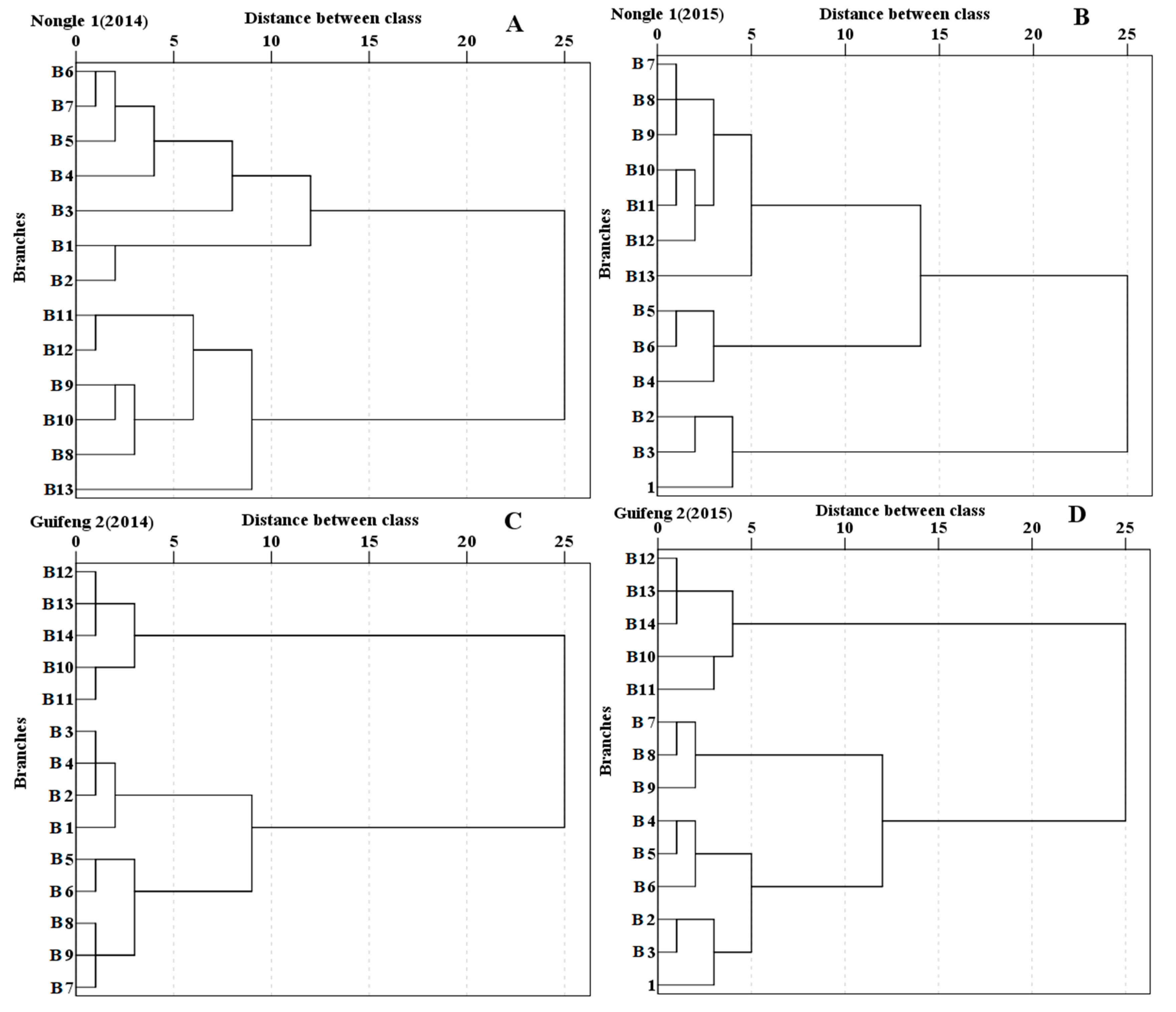
3.6. Grouping of Primary Branches
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fageria, N.K. Yield physiology of rice. J. Plant Nutr. 2007, 30, 843–879. [Google Scholar] [CrossRef]
- Fitzgerald, M.A.; Mccouch, S.R.; Hall, R.D. Not just a grain of rice: the quest for quality. Trends. Plant Sci. 2009, 14, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.; Adnan, M.; Noor, M.; Arif, M.; Alam, M.; Khan, I.A.; Ullah, H.; Wahid, F.; Mian, I.A.; Jamal, Y.; et al. Major constraints for global rice production. In Advances in Rice Research for Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Nahar, K., Biswas, J.K., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 1–22. [Google Scholar]
- Fu, J.; Xu, Y.J.; Chen, L.; Yuan, L.M.; Wang, Z.Q.; Yang, J.C. Changes in enzyme activities involved in starch synthesis and hormone concentrations in superior and inferior spikelets and their association with grain filling of super rice. Rice Sci. 2013, 20, 120–128. [Google Scholar] [CrossRef]
- Yang, J.C.; Cao, Y.Y.; Zhang, H.; Liu, L.J.; Zhang, J.H. Involvement of polyamines in the post-anthesis development of inferior and superior spikelets in rice. Planta 2008, 228, 137–149. [Google Scholar] [CrossRef]
- Huang, S.M.; Zou, Y.B.; Liu, C.L. Setting physiology of the superior and inferior grains of hybrid rice Liangyoupeijiu. Acta Agron. Sin. 2005, 31, 102–107. [Google Scholar]
- Cheng, W.D.; Zhang, G.P.; Zhao, G.P.; Yao, H.G.; Xu, H.M. Variation in rice quality of different cultivars and grain positions as affected by water management. Field Crops Res. 2003, 80, 245–252. [Google Scholar] [CrossRef]
- Sekhar, S.; Panda, B.B.; Mohapatra, T.; Das, K.; Shaw, B.P.; Kariali, E.; Mohapatra, P.K. Spikelet-specifc variation in ethylene production and constitutive expression of ethylene receptors and signal transducers during grain flling of compact and lax panicle rice (Oryza sativa) cultivars. J. Plant Physiol. 2015, 179, 21–34. [Google Scholar] [CrossRef]
- Yang, J.C. Mechanism and regulation in the filling of inferior spikelets of rice. Acta Agron. Sin. 2010, 36, 2011–2019. [Google Scholar]
- Ishimaru, T.; Matsuda, T.; Ohsugi, R.; Yamagishi, T. Morphological development of rice caryopses located at the different positions in a panicle from early to middle stage of grain filling. Funct. Plant Biol. 2003, 30, 1139–1149. [Google Scholar] [CrossRef]
- Ishimaru, T.; Hirose, T.; Matsuda, T.; Goto, A.; Takahashi, K.; Sasaki, H.; Terao, T.; Ishii, R.; Ohsugi, R.; Yamagishi, T. Expression patterns of genes encoding carbohydrate-metabolizing enzymes and their relationship to grain filling in rice (Oryza sativa L.): Comparison of caryopses located at different positions in a panicle. Plant. Cell Physiol. 2005, 46, 620–628. [Google Scholar] [CrossRef]
- Mohapatra, P.K.; Panigrahi, R.; Turner, N.C. Physiology of spikelet development on the rice panicle: Is manipulation of apical dominance crucial for grain yield improvement? In Advance Agron; Sparks, D.L., Ed.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 110, pp. 333–359. [Google Scholar]
- Yang, J.C.; Peng, S.B.; Visperas, R.M.; Sanico, A.L.; Zhu, Q.S.; Gu, S.L. Grain filling pattern and cytokinin content in the grains and roots of rice plants. Plant Growth Regul. 2000, 30, 261–270. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H. Grain filling problem in “super” rice. J. Exp. Bot. 2010, 61, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Huang, Z.H.; Wang, Z.Q.; Yang, J.C.; Zhang, J.H. Pre-anthesis non-structural carbohydrate reserve in the stem enhances the sink strength of inferior spikelets during grain filling of rice. Field Crops Res. 2011, 123, 170–182. [Google Scholar] [CrossRef]
- Kennedy, S.P.; Lynch, J.P.; Spink, J.; Bingham, I.J. Grain number and grain filling of two-row malting barley in response to variation in post-anthesis radiation: Analysis by grain position on the ear and its implications for yield improvement and quality. Field Crops Res. 2018, 225, 74–82. [Google Scholar] [CrossRef]
- Liang, W.X.; Zhang, Z.C.; Wen, X.X.; Liao, Y.C.; Liu, Y. Effect of non-structural carbohydrate accumulation in the stem pre-anthesis on grain filling of wheat inferior grain. Field Crops Res. 2017, 211, 66–76. [Google Scholar] [CrossRef]
- Li, Y.; Cui, Z.Y.; Ni, Y.L.; Zheng, M.J.; Yang, D.Q.; Jin, M.; Chen, J.; Wang, Z.L.; Yin, Y.P. Plant density effect on grain number and weight of two winter wheat cultivars at different spikelet and grain positions. PLoS ONE 2016, 11, e0155351. [Google Scholar] [CrossRef]
- Chen, Y.J.; Hoogenboom, G.; Ma, Y.T.; Li, B.G.; Guo, Y. Maize kernel growth at different floret positions of the ear. Field Crops Res. 2013, 149, 177–186. [Google Scholar] [CrossRef]
- Yu, T.; Li, G.; Liu, P.; Dong, S.T.; Zhang, J.W.; Zhao, B. Proteomics analysis of maize (Zea mays L.) grain based on iTRAQ reveals molecular mechanisms of poor grain filling in inferior grains. Plant Physiol. Biochem. 2017, 115, 83–96. [Google Scholar] [CrossRef]
- Nagato, K. Differences in grain weight of spikelets located at different positions within a rice panicle. Jpn. J. Crop Sci. 1941, 13, 156–169. [Google Scholar] [CrossRef][Green Version]
- Wang, F.; Chen, S.; Cheng, F.M.; Liu, Y.; Zhang, G.P. The differences in grain weight and quality within a rice (Oryza sativa L.) panicle as affected by panicle type and source-sink relation. J. Agron. Crop Sci. 2007, 193, 63–73. [Google Scholar] [CrossRef]
- Peng, T.; Lv, Q.; Zhao, Y.F.; Sun, H.Z.; Han, Y.C.; Du, Y.X.; Zhang, J.; Li, J.Z.; Wang, L.L.; Zhao, Q.Z. Superior grains determined by grain weight are not fully correlated with the flowering order in rice. J. Integr. Agric. 2015, 14, 847–855. [Google Scholar] [CrossRef]
- Jiang, Q.; Du, Y.L.; Tian, X.Y.; Wang, Q.S.; Xiong, R.H.; Xu, G.C.; Yan, C.; Ding, Y.F. Effect of panicle nitrogen on grain filling characteristics of high-yielding rice cultivars. Eur. J. Agron. 2016, 74, 185–192. [Google Scholar] [CrossRef]
- Zhang, H.; Li, H.W.; Yuan, L.M.; Wang, Z.Q.; Yang, J.C.; Zhang, J.H. Post-anthesis alternate wetting and moderate soil drying enhances activities of key enzymes in sucrose-to-starch conversion in inferior spikelets of rice. J. Exp. Bot. 2012, 63, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.X.; Chen, J.; Lin, S.S.; Li, Z.; Cheng, R.H.; Fang, C.X.; Chen, H.F.; Lin, W.X. Proteomic and phosphoproteomic determination of ABA’s effects on grain-filling of Oryza sativa L. inferior spikelets. Plant Sci. 2012, 185–186, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Cheng, F.M.; Zhang, G.P. The relationship between grain filling and hormone content as affected by genotype and source-sink relation. Plant Growth Regul. 2006, 49, 1–8. [Google Scholar]
- Wei, H.Y.; Zhu, Y.; Qiu, S.; Han, C.; Hu, L.; Xu, D.; Zhou, N.B.; Xing, Z.P.; Hu, Y.J.; Cui, P.Y.; et al. Combined effect of shading time and nitrogen level on grain filling and grain quality in japonica super rice. J. Integr. Agric. 2018, 17, 2405–2417. [Google Scholar] [CrossRef]
- Richards, F.J. A flexible growth function for empirical use. J. Exp. Bot. 1959, 10, 290–301. [Google Scholar] [CrossRef]
- Cheng, M.L. Parameter estimation of Richards model and its application. Math. Pract. Theory. 2010, 40, 139–143. [Google Scholar]
- Zhu, Q.S.; Cao, X.Z.; Luo, Y.Q. Growth analysis on the process of grain filling in rice. Acta Agron. Sin. 1988, 14, 182–193. [Google Scholar]
- Zhang, H.; Liu, K.; Wang, Z.Q.; Liu, L.J.; Yang, J.C. Abscisic acid, ethylene and antioxidative systems in rice grains in relation with grain filling subjected to postanthesis soil-drying. Plant Growth Regul. 2015, 76, 135–146. [Google Scholar] [CrossRef]
- Adolfsson, A.; Ackerman, M.; Brownstein, N.C. To cluster, or not to cluster: An analysis of clusterability methods. Pattern Recogn. 2019, 88, 13–26. [Google Scholar] [CrossRef]
- Mohapatra, P.K.; Patel, R.; Sahu, S.K. Time of flowering affects grain quality and spikelet partitioning within the rice panicle. Aust. J. Plant Physiol. 1993, 20, 231–242. [Google Scholar] [CrossRef]
- Naik, P.K.; Mohapatra, P.K. Ethylene inhibitors promote male gametophyte survival in rice. Plant Growth Regul. 1999, 28, 29–39. [Google Scholar] [CrossRef]
- Chen, L.; Deng, Y.; Zhu, H.L.; Hu, Y.X.; Jiang, Z.G.; Tang, S.; Wang, S.H.; Ding, Y.F. The initiation of inferior grain filling is affected by sugar translocation efficiency in large panicle rice. Rice 2019, 12, 75. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.L.; Zhu, Q.S.; Yang, J.C.; Peng, S.B. Analysis on grain filling characteristics for different rice types. Acta Agron. Sin. 2001, 27, 7–14. [Google Scholar]
- Liang, J.S.; Zhang, J.H.; Cao, X.Z. Grain sink strength may be related to the poor grain filling of indica-japonica rice (Oryza sativa) hybrids. Physiol. Plantarum. 2001, 112, 470–477. [Google Scholar] [CrossRef]
- Leonard, M.; Kinet, J.M.; Bodson, M.; Bernier, G. Enhanced inflorescence development in tomato by growth substance treatments in relation to 14C-assimilate distribution. Physiol. Plantarum. 1983, 57, 85–89. [Google Scholar] [CrossRef]
- Kinet, J.M.; Zime, V.; Linotte, A.; Jacqmard, A.; Bernier, G. Resumption of cellular activity induced by cytokinin and gibberellin treatments in tomato flowers targeted for abortion unfavorable light conditions. Physiol. Plantarum. 1986, 64, 67–73. [Google Scholar] [CrossRef]
- Feng, S.W.; Hu, T.Z.; Li, G.; Dong, N.; Li, X.H.; Ru, Z.G.; Cheng, Z.H. Analysis of grain filling characteristics of different wheat varieties. J. Triticeae Crops 2009, 29, 643–646. [Google Scholar]
- Dong, M.H.; Chen, P.F.; Xie, Y.L.; Qiao, Z.Y.; Yang, J.C. Variations in carbohydrate and protein accumulation among spikelets at different positions within a panicle during rice grain filling. Rice Sci. 2012, 19, 223–232. [Google Scholar] [CrossRef]
- Lv, X.K.; Li, T.; Wen, X.X.; Liao, Y.C.; Liu, Y. Effect of potassium foliage application post-anthesis on grain filling of wheat under drought stress. Field Crops Res. 2017, 206, 95–105. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H.; Wang, Z.Q.; Xu, G.W.; Zhu, Q.S. Activities of key enzymes in sucrose-to-starch conversion in wheat grains subjected to water deficit during grain filling. Plant Physiol. 2004, 135, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Jeng, T.L.; Wang, C.S.; Chen, C.L.; Sung, J.M. Effects of grain position on the panicle on starch biosynthetic enzyme activity in developing grains of rice cultivar Tainung 67 and its NaN3-induced mutant. J. Agric. Sci. 2003, 141, 303–311. [Google Scholar] [CrossRef]
- Das, K.; Panda, B.B.; Shaw, B.P.; Das, S.R.; Dash, S.K.; Kariali, E.; Mohapatra, P.K. Grain density and its impact on grain filling characteristic of rice: mechanistic testing of the concept in genetically related cultivars. Sci. Rep. 2018, 8, 4149. [Google Scholar] [CrossRef] [PubMed]
- Panigrahi, R.; Kariali, E.; Panda, B.B.; Lafarge, T.; Mohapatra, P.K. Controlling the trade-off between spikelet number and grain filling: the hierarchy of starch synthesis in spikelets of rice panicle in relation to hormone dynamics. Funct. Plant Biol. 2019, 46, 507–523. [Google Scholar] [CrossRef]
- Chen, T.T.; Xu, G.W.; Wang, Z.Q.; Zhang, H.; Yang, J.C.; Zhang, J.H. Expression of proteins in superior and inferior spikelets of rice during grain filling under different irrigation regimes. Proteomics 2016, 16, 102–121. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H.; Wang, Z.Q.; Zhu, Q.S.; Wang, W. Remobilization of carbon reserves in response to water deficit during grain filling of rice. Field Crops Res. 2001, 71, 47–55. [Google Scholar] [CrossRef]
- Li, Z.; Li, Z.; Muhammad, W.; Lin, M.H.; Azeem, S.; Zhao, H.; Lin, S.; Chen, T.; Fang, C.X.; Letuma, P.; et al. Proteomic analysis of positive influence of alternate wetting and moderate soil drying on the process of rice grain filling. Plant Growth Regul. 2018, 84, 533–548. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H.; Wang, Z.Q.; Liu, K.; Wang, P. Post-anthesis development of inferior and superior spikelets in rice in relation to abscisic acid and ethylene. J. Exp. Bot. 2006, 57, 149–160. [Google Scholar] [CrossRef]
- Liu, Y.; Sui, Y.W.; Gu, D.D.; Wen, X.X.; Chen, Y.; Li, C.J.; Liao, Y.C. Effects of conservation tillage on grain filling and hormonal changes in wheat under simulated rainfall conditions. Field Crops Res. 2013, 144, 43–51. [Google Scholar] [CrossRef]
- Xu, G.W.; Zhang, J.H.; Lam, H.M.; Wang, Z.Q.; Yang, J.C. Hormonal changes are related to the poor grain filling in the inferior spikelets of rice cultivated under non-flooded and mulched condition. Field Crops Res. 2007, 101, 53–61. [Google Scholar] [CrossRef]
- Xu, X.B.; Vergara, B.S. Morphological changes in rice panicle development: a review of literature. IRRI Res. Pap. Ser. 1986, 117, 1–16. [Google Scholar]
- Okamura, M.; Arai-Sanoh, Y.; Yoshida, H.; Mukouyama, T.; Adachi, S.; Yabe, S.; Nakagawa, H.; Tsutsumi, K.; Taniguchi, Y.; Kobayashi, N.; et al. Characterization of high-yielding rice cultivars with different grain-filling properties to clarify limiting factors for improving grain yield. Field Crops Res. 2018, 219, 139–147. [Google Scholar] [CrossRef]
- You, C.C.; Zhu, H.L.; Xu, B.B.; Huang, W.X.; Wang, S.H.; Ding, Y.F.; Liu, Z.H.; Li, G.H.; Chen, L.; Ding, C.Q.; et al. Effect of removing superior spikelets on grain filling of inferior spikelets in rice. Front. Plant Sci. 2016, 7, 1161. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2014 (Late Rice) | 2015 (Early Rice) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cultivars | Branches | A | B | k | N | R2 | Branches | A | B | k | N | R2 |
| Nongle 1 | B1 | 2.011 | 45.84 | 0.289 | 1.750 | 0.996 | B1 | 1.935 | 57.56 | 0.402 | 2.133 | 0.995 |
| B2 | 2.040 | 52.75 | 0.269 | 1.856 | 0.994 | B2 | 1.951 | 24.91 | 0.322 | 1.661 | 0.999 | |
| B3 | 1.895 | 56.62 | 0.259 | 1.948 | 0.997 | B3 | 1.873 | 49.37 | 0.325 | 2.043 | 0.999 | |
| B4 | 1.865 | 60.82 | 0.255 | 2.011 | 0.997 | B4 | 1.845 | 12.17 | 0.229 | 1.311 | 0.997 | |
| B5 | 1.835 | 131.13 | 0.279 | 2.411 | 0.998 | B5 | 1.762 | 20.67 | 0.228 | 1.580 | 0.998 | |
| B6 | 1.742 | 311.13 | 0.299 | 2.791 | 0.998 | B6 | 1.708 | 27.32 | 0.231 | 1.651 | 0.998 | |
| B7 | 1.727 | 597.14 | 0.321 | 3.211 | 0.995 | B7 | 1.681 | 305.80 | 0.322 | 3.100 | 0.995 | |
| B8 | 1.739 | 4521.61 | 0.392 | 4.334 | 0.999 | B8 | 1.659 | 393.61 | 0.325 | 3.180 | 0.998 | |
| B9 | 1.634 | 11139.48 | 0.407 | 4.867 | 0.997 | B9 | 1.620 | 665.46 | 0.331 | 3.411 | 0.996 | |
| B10 | 1.611 | 140115.85 | 0.498 | 6.071 | 0.998 | B10 | 1.519 | 1239.35 | 0.350 | 3.750 | 0.993 | |
| B11 | 1.645 | 756627.73 | 0.526 | 6.812 | 0.996 | B11 | 1.544 | 1614.66 | 0.352 | 3.790 | 0.989 | |
| B12 | 1.587 | 1186041.88 | 0.537 | 7.410 | 0.992 | B12 | 1.611 | 2507.23 | 0.353 | 4.064 | 0.994 | |
| B13 | 1.417 | 8710836.16 | 0.594 | 8.519 | 0.993 | B13 | 1.602 | 13194.86 | 0.401 | 4.990 | 0.990 | |
| panicle | 1.855 | 69.38 | 0.22 | 2.122 | 0.999 | panicle | 1.765 | 14.45 | 0.199 | 1.394 | 0.998 | |
| Guifeng 2 | B1 | 1.572 | 21.11 | 0.283 | 1.622 | 0.999 | B1 | 1.651 | 23.31 | 0.311 | 1.687 | 0.998 |
| B2 | 1.531 | 30.47 | 0.283 | 1.792 | 0.999 | B2 | 1.621 | 20.56 | 0.281 | 1.551 | 0.975 | |
| B3 | 1.560 | 27.06 | 0.265 | 1.674 | 0.998 | B3 | 1.620 | 13.09 | 0.261 | 1.279 | 0.993 | |
| B4 | 1.529 | 33.10 | 0.266 | 1.787 | 0.998 | B4 | 1.629 | 14.82 | 0.252 | 1.361 | 0.995 | |
| B5 | 1.560 | 45.09 | 0.260 | 1.895 | 0.997 | B5 | 1.680 | 20.31 | 0.250 | 1.457 | 0.997 | |
| B6 | 1.530 | 53.49 | 0.256 | 1.992 | 0.997 | B6 | 1.654 | 25.67 | 0.244 | 1.568 | 0.997 | |
| B7 | 1.497 | 89.62 | 0.261 | 2.100 | 0.996 | B7 | 1.610 | 36.38 | 0.234 | 1.701 | 0.995 | |
| B8 | 1.480 | 180.82 | 0.281 | 2.311 | 0.996 | B8 | 1.601 | 75.35 | 0.252 | 2.011 | 0.997 | |
| B9 | 1.465 | 268.17 | 0.289 | 2.544 | 0.996 | B9 | 1.603 | 197.98 | 0.286 | 2.488 | 0.995 | |
| B10 | 1.370 | 3598.77 | 0.377 | 3.844 | 0.996 | B10 | 1.530 | 1516.54 | 0.345 | 3.477 | 0.995 | |
| B11 | 1.301 | 23999.85 | 0.432 | 4.751 | 0.995 | B11 | 1.484 | 4021.75 | 0.369 | 3.885 | 0.994 | |
| B12 | 1.311 | 138239.93 | 0.485 | 5.751 | 0.993 | B12 | 1.471 | 42367.89 | 0.451 | 5.102 | 0.994 | |
| B13 | 1.306 | 354717.29 | 0.512 | 6.250 | 0.993 | B13 | 1.474 | 118240.28 | 0.491 | 5.801 | 0.990 | |
| B14 | 1.271 | 1603900.56 | 0.544 | 6.890 | 0.990 | B14 | 1.479 | 328969.32 | 0.511 | 6.119 | 0.984 | |
| panicle | 1.527 | 25.84 | 0.207 | 1.629 | 0.998 | panicle | 1.659 | 12.31 | 0.189 | 1.252 | 0.998 | |
| Cultivars | Season | Branches | B2 | B3 | B4 | B5 | B6 | B7 | B8 | B9 | B10 | B11 | B12 | B13 | Panicle |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nongle 1 | 2014 (late rice) | B1 | 0.989 ** | 0.970 ** | 0.973 ** | 0.961 ** | 0.908 ** | 0.911 ** | 0.876 ** | 0.835 ** | 0.765 ** | 0.664 ** | 0.657 ** | 0.571 ** | 0.909 ** |
| B2 | 0.980 ** | 0.995 ** | 0.978 ** | 0.951 ** | 0.949 ** | 0.914 ** | 0.892 ** | 0.830 ** | 0.757 ** | 0.744 ** | 0.666 ** | 0.949 ** | |||
| B3 | 0.977 ** | 0.975 ** | 0.949 ** | 0.950 ** | 0.926 ** | 0.893 ** | 0.847 ** | 0.764 ** | 0.763 ** | 0.694 ** | 0.954 ** | ||||
| B4 | 0.987 ** | 0.974 ** | 0.971 ** | 0.939 ** | 0.927 ** | 0.871 ** | 0.807 ** | 0.796 ** | 0.727 ** | 0.974 ** | |||||
| B5 | 0.983 ** | 0.987 ** | 0.973 ** | 0.947 ** | 0.906 ** | 0.817 ** | 0.823 ** | 0.757 ** | 0.983 ** | ||||||
| B6 | 0.999 ** | 0.987 ** | 0.987 ** | 0.959 ** | 0.908 ** | 0.908 ** | 0.857 ** | 0.996 ** | |||||||
| B7 | 0.993 ** | 0.986 ** | 0.960 ** | 0.898 ** | 0.904 ** | 0.852 ** | 0.995 ** | ||||||||
| B8 | 0.987 ** | 0.976 ** | 0.905 ** | 0.921 ** | 0.873 ** | 0.981 ** | |||||||||
| B9 | 0.990 ** | 0.954 ** | 0.960 ** | 0.924 ** | 0.977 ** | ||||||||||
| B10 | 0.969 ** | 0.982 ** | 0.957 ** | 0.946 ** | |||||||||||
| B11 | 0.994 ** | 0.983 ** | 0.889 ** | ||||||||||||
| B12 | 0.992 ** | 0.892 ** | |||||||||||||
| B13 | 0.842 ** | ||||||||||||||
| 2015 (early rice) | B1 | 0.995 ** | 0.969 ** | 0.936 ** | 0.871 ** | 0.808 ** | 0.798 ** | 0.748 ** | 0.692 ** | 0.647 ** | 0.610 ** | 0.557 ** | 0.42 | 0.827 ** | |
| B2 | 0.986 ** | 0.965 ** | 0.913 ** | 0.860 ** | 0.851 ** | 0.807 ** | 0.756 ** | 0.714 ** | 0.680 ** | 0.631 ** | 0.501 * | 0.876 ** | |||
| B3 | 0.986 ** | 0.959 ** | 0.922 ** | 0.912 ** | 0.878 ** | 0.836 ** | 0.797 ** | 0.762 ** | 0.717 ** | 0.614 ** | 0.929 ** | ||||
| B4 | 0.986 ** | 0.961 ** | 0.953 ** | 0.926 ** | 0.893 ** | 0.864 ** | 0.838 ** | 0.800 ** | 0.693 ** | 0.971 ** | |||||
| B5 | 0.993 ** | 0.986 ** | 0.972 ** | 0.950 ** | 0.927 ** | 0.902 ** | 0.874 ** | 0.796 ** | 0.996 ** | ||||||
| B6 | 0.995 ** | 0.990 ** | 0.978 ** | 0.960 ** | 0.939 ** | 0.918 ** | 0.858 ** | 0.997 ** | |||||||
| B7 | 0.997 ** | 0.987 ** | 0.974 ** | 0.956 ** | 0.938 ** | 0.878 ** | 0.990 ** | ||||||||
| B8 | 0.987 ** | 0.971 ** | 0.960 ** | 0.913 ** | 0.982 ** | ||||||||||
| B9 | 0.996 ** | 0.985 ** | 0.977 ** | 0.942 ** | 0.967 ** | ||||||||||
| B10 | 0.996 ** | 0.990 ** | 0.953 ** | 0.949 ** | |||||||||||
| B11 | 0.993 ** | 0.951 ** | 0.928 ** | ||||||||||||
| B12 | 0.972 ** | 0.905 ** | |||||||||||||
| B13 | 0.833 ** |
| Cultivars | Season | Branches | B2 | B3 | B4 | B5 | B6 | B7 | B8 | B9 | B10 | B11 | B12 | B13 | B14 | Panicle |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Guifeng 2 | late rice (2014) | B1 | 0.994 ** | 0.982 ** | 0.974 ** | 0.937 ** | 0.912 ** | 0.832 ** | 0.750 ** | 0.741 ** | 0.644 ** | 0.537 * | 0.481 * | 0.448 * | 0.377 | 0.875 ** |
| B2 | 0.996 ** | 0.992 ** | 0.967 ** | 0.948 ** | 0.881 ** | 0.804 ** | 0.797 ** | 0.708 ** | 0.604 ** | 0.548 * | 0.517 * | 0.451 * | 0.914 ** | |||
| B3 | 0.999 ** | 0.985 ** | 0.971 ** | 0.916 ** | 0.848 ** | 0.842 ** | 0.759 ** | 0.659 ** | 0.604 ** | 0.574 ** | 0.509 * | 0.942 ** | ||||
| B4 | 0.991 ** | 0.979 ** | 0.930 ** | 0.866 ** | 0.860 ** | 0.781 ** | 0.684 ** | 0.630 ** | 0.601 ** | 0.537 * | 0.953 ** | |||||
| B5 | 0.997 ** | 0.971 ** | 0.925 ** | 0.920 ** | 0.856 ** | 0.770 ** | 0.721 ** | 0.693 ** | 0.629 ** | 0.980 ** | ||||||
| B6 | 0.986 ** | 0.949 ** | 0.945 ** | 0.890 ** | 0.813 ** | 0.767 ** | 0.741 ** | 0.680 ** | 0.991 ** | |||||||
| B7 | 0.987 ** | 0.986 ** | 0.952 ** | 0.895 ** | 0.858 ** | 0.837 ** | 0.784 ** | 0.990 ** | ||||||||
| B8 | 0.999 ** | 0.987 ** | 0.949 ** | 0.922 ** | 0.904 ** | 0.849 ** | 0.965 ** | |||||||||
| B9 | 0.989 ** | 0.956 ** | 0.930 ** | 0.914 ** | 0.864 ** | 0.963 ** | ||||||||||
| B10 | 0.986 ** | 0.970 ** | 0.957 ** | 0.914 ** | 0.916 ** | |||||||||||
| B11 | 0.997 ** | 0.992 ** | 0.964 ** | 0.855 ** | ||||||||||||
| B12 | 0.998 ** | 0.974 ** | 0.814 ** | |||||||||||||
| B13 | 0.987 ** | 0.792 ** | ||||||||||||||
| B14 | 0.738 ** | |||||||||||||||
| early rice (2015) | B1 | 0.994 ** | 0.978 ** | 0.978 ** | 0.945 ** | 0.918 ** | 0.834 ** | 0.756 ** | 0.742 ** | 0.638 ** | 0.550 ** | 0.467 * | 0.475 * | 0.376 | 0.868 ** | |
| B2 | 0.983 ** | 0.991 ** | 0.970 ** | 0.952 ** | 0.888 ** | 0.821 ** | 0.808 ** | 0.717 ** | 0.635 ** | 0.558 ** | 0.564 ** | 0.472 * | 0.916 ** | |||
| B3 | 0.994 ** | 0.976 ** | 0.954 ** | 0.890 ** | 0.808 ** | 0.804 ** | 0.702 ** | 0.625 ** | 0.530 * | 0.541 * | 0.440 * | 0.909 ** | ||||
| B4 | 0.987 ** | 0.973 ** | 0.922 ** | 0.854 ** | 0.847 ** | 0.758 ** | 0.683 ** | 0.598 ** | 0.607 ** | 0.515 * | 0.941 ** | |||||
| B5 | 0.996 ** | 0.960 ** | 0.912 ** | 0.907 ** | 0.828 ** | 0.763 ** | 0.688 ** | 0.697 ** | 0.606 ** | 0.969 ** | ||||||
| B6 | 0.981 ** | 0.945 ** | 0.940 ** | 0.874 ** | 0.817 ** | 0.749 ** | 0.757 ** | 0.673 ** | 0.986 ** | |||||||
| B7 | 0.986 ** | 0.985 ** | 0.946 ** | 0.908 ** | 0.851 ** | 0.857 ** | 0.792 ** | 0.993 ** | ||||||||
| B8 | 0.998 ** | 0.982 ** | 0.956 ** | 0.921 ** | 0.925 ** | 0.874 ** | 0.972 ** | |||||||||
| B9 | 0.986 ** | 0.963 ** | 0.926 ** | 0.932 ** | 0.881 ** | 0.964 ** | ||||||||||
| B10 | 0.993 ** | 0.975 ** | 0.977 ** | 0.946 ** | 0.917 ** | |||||||||||
| B11 | 0.990 ** | 0.992 ** | 0.971 ** | 0.871 ** | ||||||||||||
| B12 | 0.999 ** | 0.993 ** | 0.811 ** | |||||||||||||
| B13 | 0.991 ** | 0.815 ** | ||||||||||||||
| B14 | 0.745** |
| Cultivars | Season | Branch Groups | Branches | A | Vmax | D | Tmax | R0 | 0–t1 | t1–t2 | t2–t3 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nongle 1 | late rice (2014) | I | B1-B2 | 2.081a | 0.101a | 31.4b | 12.0b | 0.143a | 5.6b | 12.8a | 13.5a |
| II | B3-B7 | 1.857b | 0.082b | 34.8a | 14.5b | 0.112ab | 7.7b | 13.5a | 12.8a | ||
| III | B8-B13 | 1.619c | 0.079b | 34.1a | 21.3a | 0.076b | 16.9a | 8.7b | 4.3b | ||
| early rice (2015) | I | B1-B3 | 1.941a | 0.120a | 24.3b | 7.7c | 0.238a | 2.4b | 10.6b | 13.0a | |
| II | B4-B6 | 1.785b | 0.086b | 31.6a | 11.0b | 0.151b | 4.4b | 13.2a | 14.5a | ||
| III | B7-B13 | 1.610c | 0.079b | 33.3a | 17.6a | 0.085c | 12.6a | 10.0b | 6.3b | ||
| Guifeng 2 | late rice (2014) | I | B1-B4 | 1.563a | 0.085a | 28.0b | 10.0c | 0.165a | 4.2c | 11.5a | 12.4a |
| II | B5-B9 | 1.524a | 0.072b | 32.7a | 14.3b | 0.118b | 8.0b | 12.5a | 11.7a | ||
| III | B10-B14 | 1.339b | 0.068b | 33.2a | 22.2a | 0.075c | 18.2a | 8.0b | 3.6b | ||
| early rice (2015) | I | B1-B6 | 1.658a | 0.091a | 27.6b | 9.4c | 0.190a | 3.5c | 11.8a | 13.7a | |
| II | B7-B9 | 1.629a | 0.077a | 32.5a | 14.6b | 0.121b | 8.2b | 12.7a | 12.0a | ||
| III | B10-B14 | 1.519b | 0.077a | 32.8a | 20.9a | 0.081c | 16.4a | 8.8b | 4.7b |
| Nongle 1 | Guifeng 2 | |||
|---|---|---|---|---|
| 2014 | 2015 | 2014 | 2015 | |
| B1 | 16.6 | 16.1 | 15.6 | 15.6 |
| B2 | 18.1 | 18.5 | 16.96 | 17.2 |
| B3 | 18.7 | 20.7 | 16.50 | 17.3 |
| B4 | 18.7 | 23.2 | 16.5 | 16.7 |
| B5 | 18.4 | 23.3 | 16.5 | 17.2 |
| B6 | 17.1 | 22.3 | 16.30 | 16.5 |
| B7 | 17.9 | 23.3 | 14.6 | 14.7 |
| B8 | 17.5 | 22.5 | 13.50 | 14.1 |
| B9 | 15.6 | 20.9 | 13.50 | 13.9 |
| B10 | 16.3 | 21.8 | 11.7 | 12.0 |
| B11 | 14.3 | 19.0 | 10.4 | 10.7 |
| B12 | 14.6 | 18.7 | 9.9 | 10.2 |
| B13 | 15.2 | 18.5 | 9.6 | 10.3 |
| B14 | 9.4 | 9.2 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Q.; Hao, X.; Ali, I.; Iqbal, A.; Ullah, S.; Huang, M.; Kong, F.; Li, T.; Xuan, Y.; Li, F.; et al. Characterization and Grouping of All Primary Branches at Various Positions on a Rice Panicle Based on Grain Growth Dynamics. Agronomy 2020, 10, 223. https://doi.org/10.3390/agronomy10020223
Zhao Q, Hao X, Ali I, Iqbal A, Ullah S, Huang M, Kong F, Li T, Xuan Y, Li F, et al. Characterization and Grouping of All Primary Branches at Various Positions on a Rice Panicle Based on Grain Growth Dynamics. Agronomy. 2020; 10(2):223. https://doi.org/10.3390/agronomy10020223
Chicago/Turabian StyleZhao, Quan, Xiangyang Hao, Izhar Ali, Anas Iqbal, Saif Ullah, Min Huang, Feiyang Kong, Tianyuan Li, Ying Xuan, Faqiao Li, and et al. 2020. "Characterization and Grouping of All Primary Branches at Various Positions on a Rice Panicle Based on Grain Growth Dynamics" Agronomy 10, no. 2: 223. https://doi.org/10.3390/agronomy10020223
APA StyleZhao, Q., Hao, X., Ali, I., Iqbal, A., Ullah, S., Huang, M., Kong, F., Li, T., Xuan, Y., Li, F., Yan, B., Luo, Y., Liang, H., Wei, S., Chen, N., & Jiang, L. (2020). Characterization and Grouping of All Primary Branches at Various Positions on a Rice Panicle Based on Grain Growth Dynamics. Agronomy, 10(2), 223. https://doi.org/10.3390/agronomy10020223