Effects of Depolymerized Gellan with Different Molecular Weights on the Growth of Four Bedding Plant Species


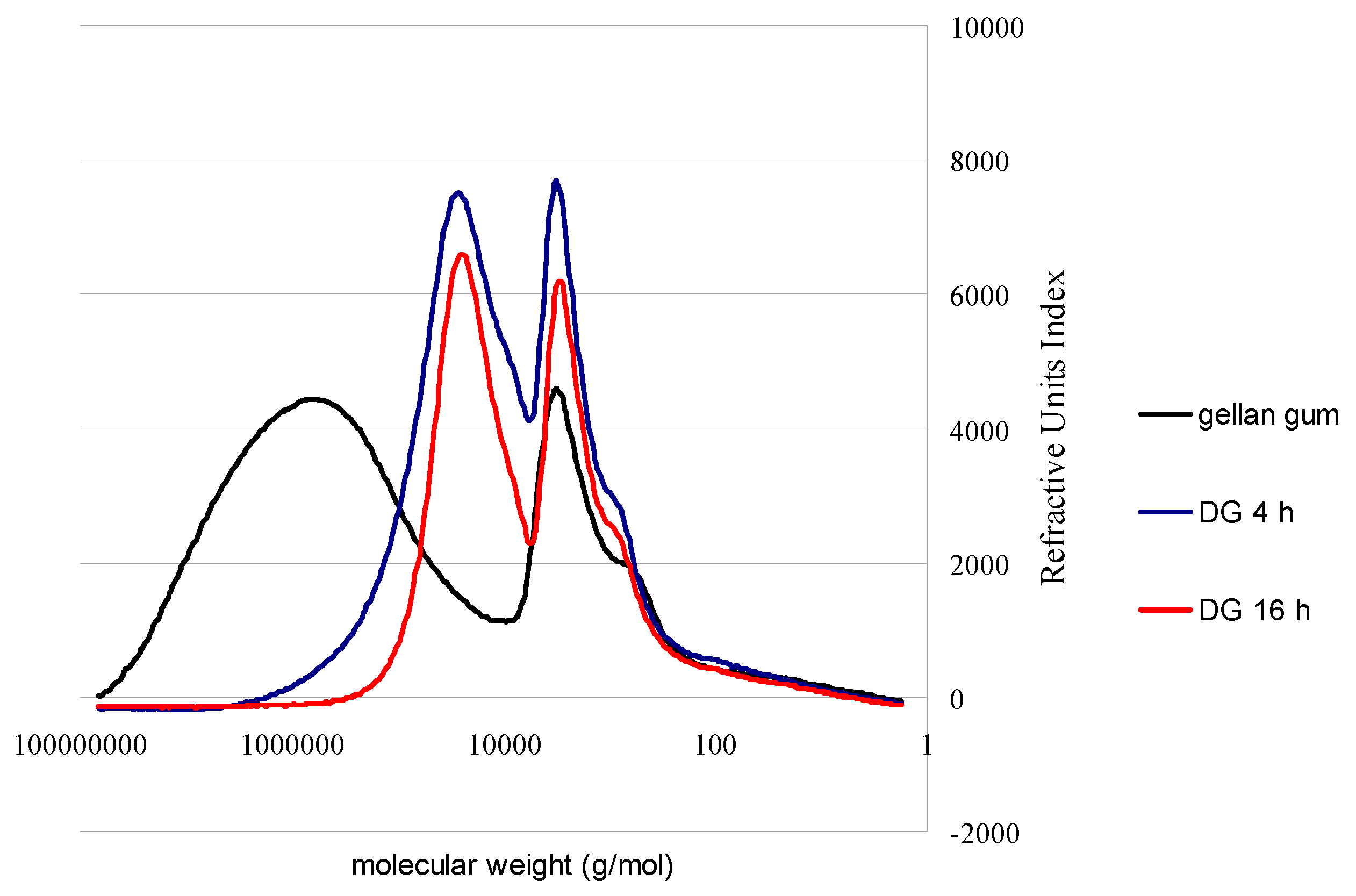{kind=link}
{kind=link}
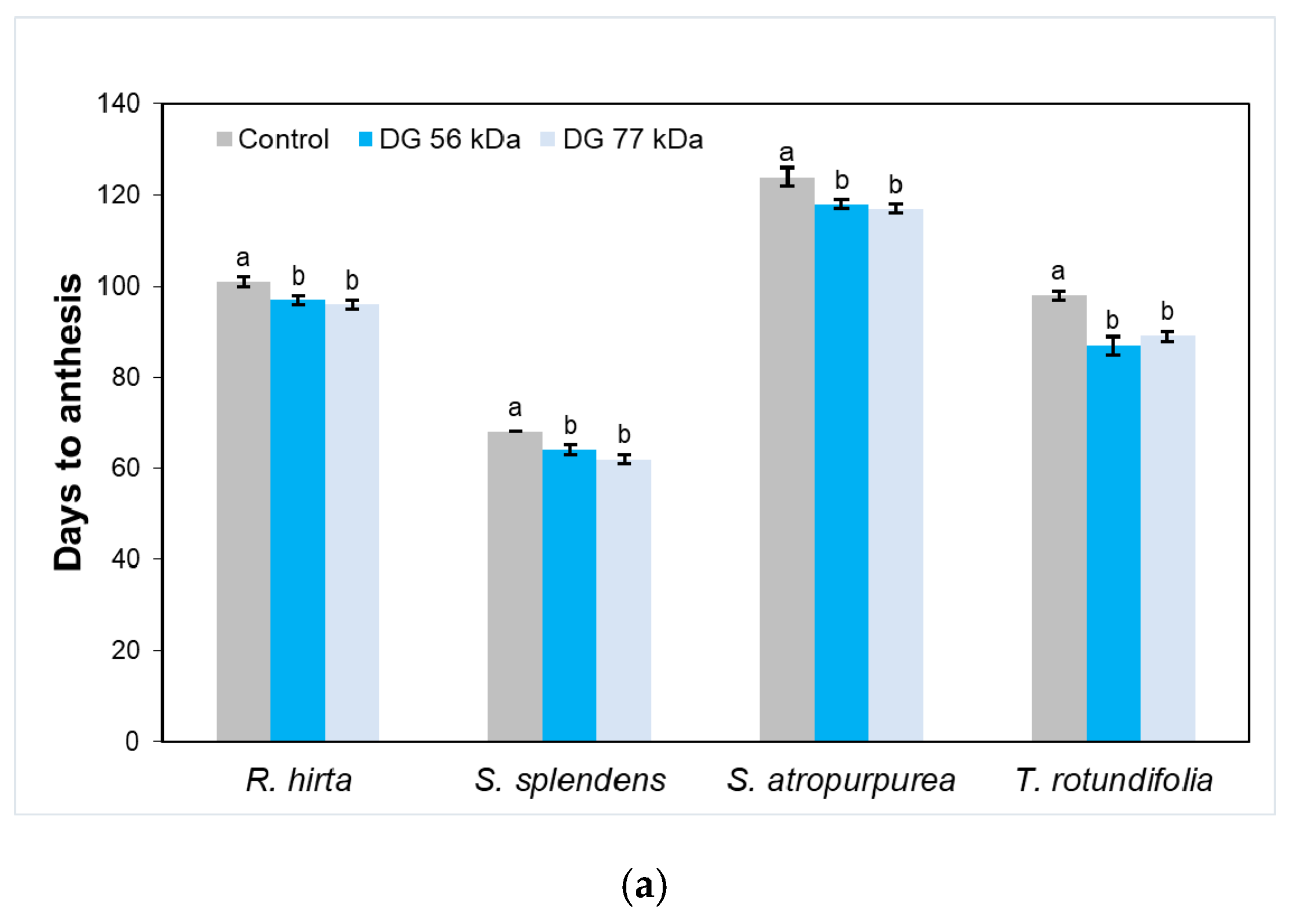{kind=link}
{kind=link}
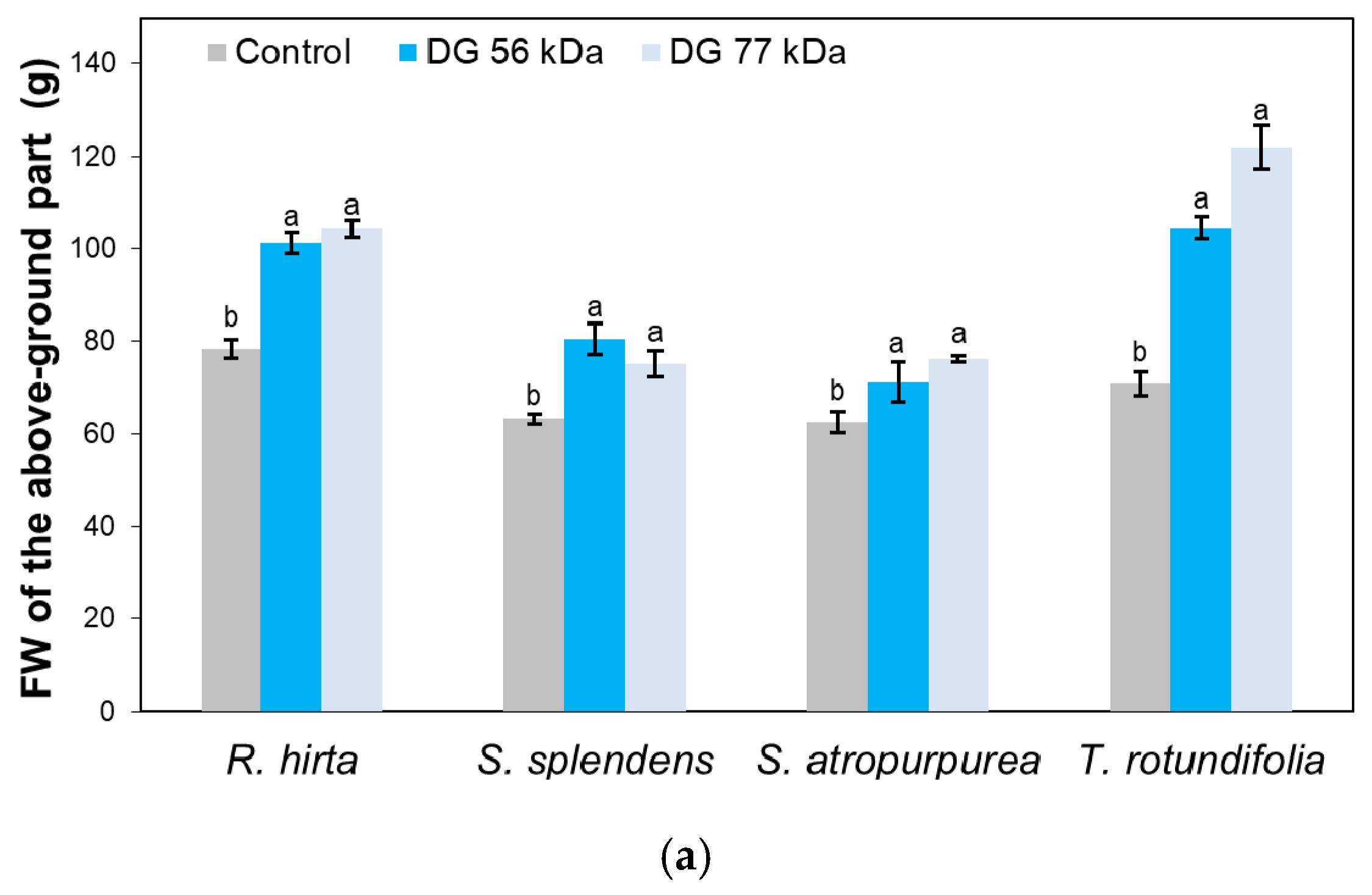{kind=link}
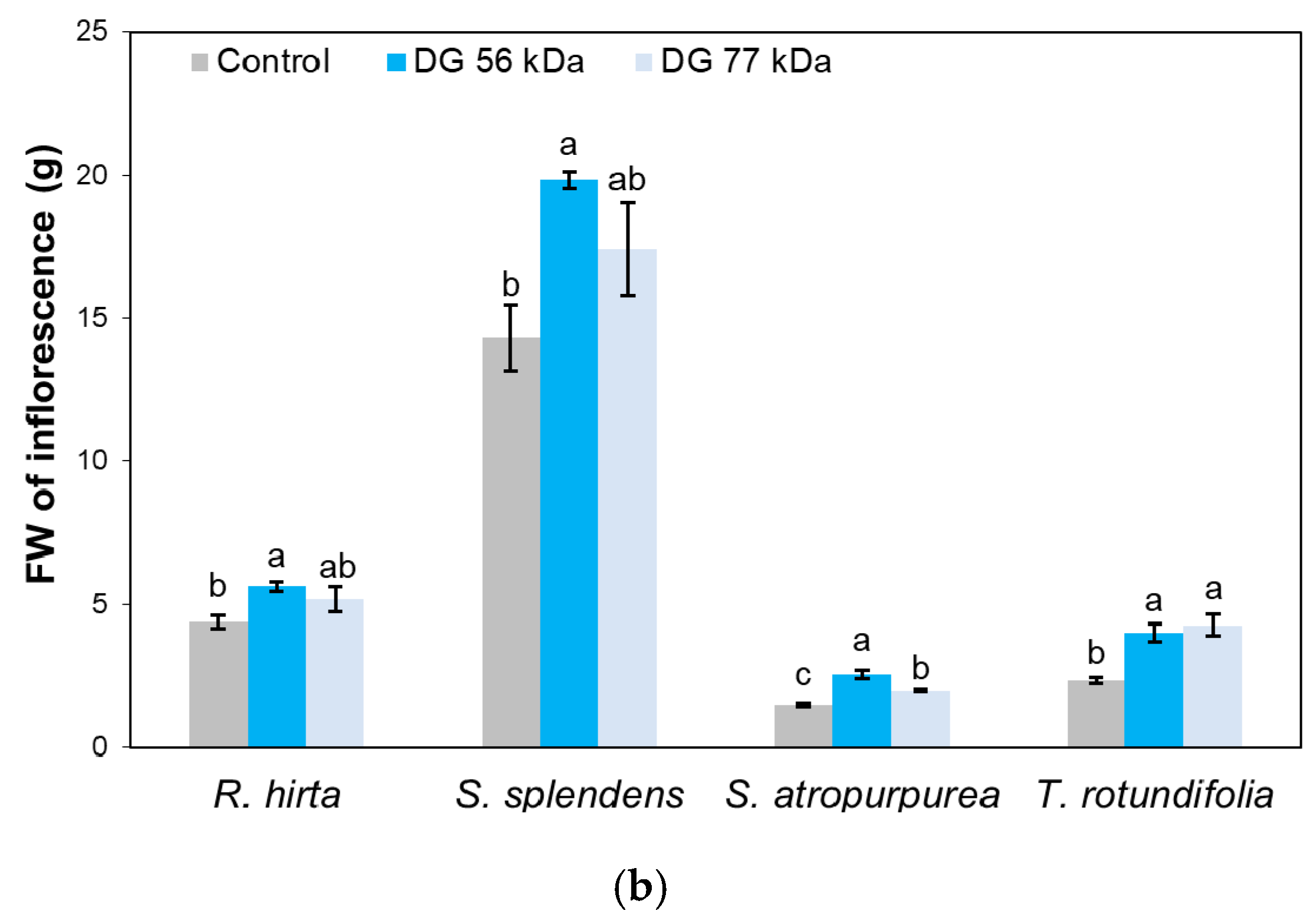{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation and Determination of Depolymerized Gellan
2.2. Plant Material and Experimental Design
2.3. Statistical Analysis of Experimental Data
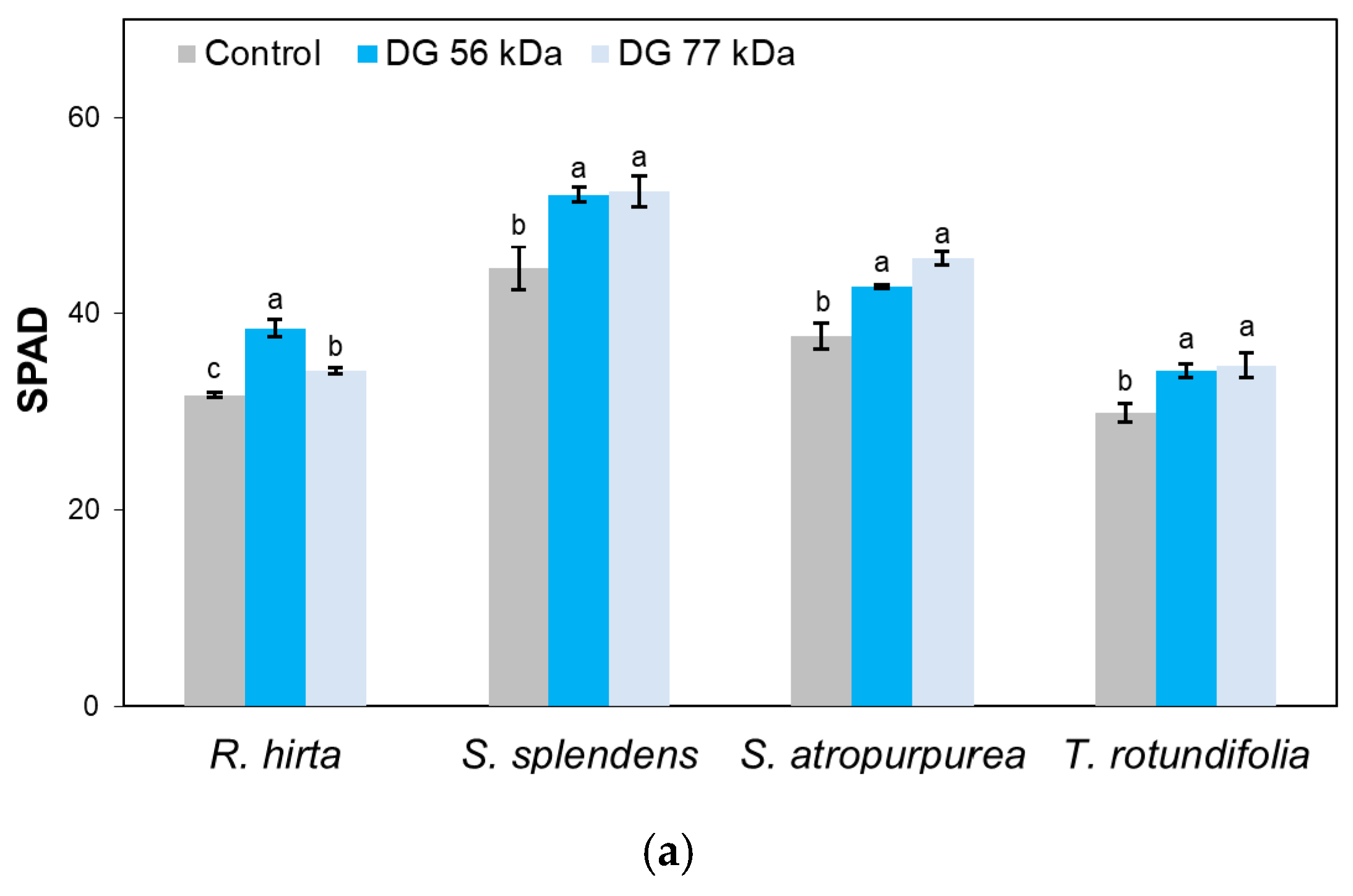
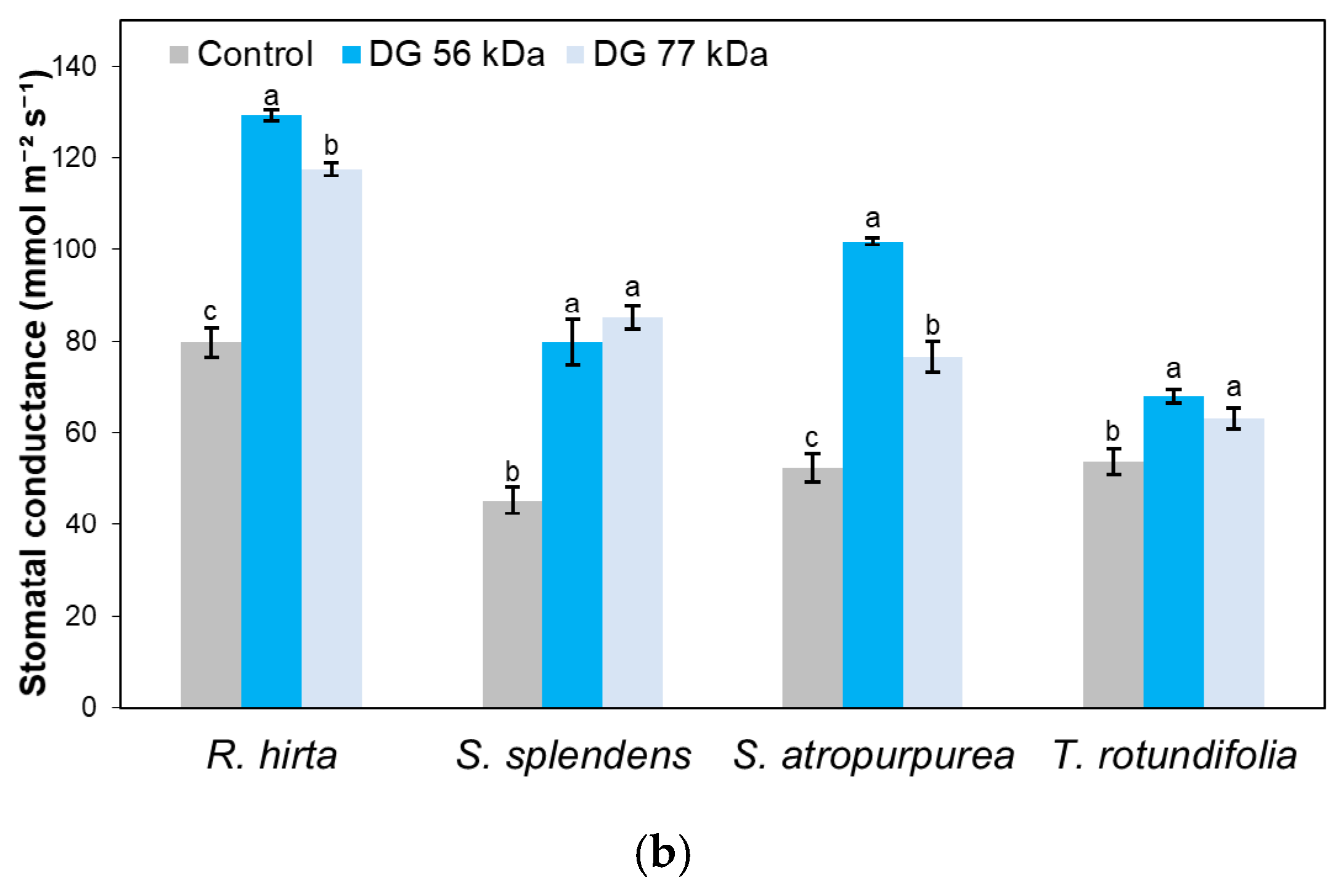
3. Results
4. Discussion
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Drew, J.; Yue, C.; Anderson, N.O.; Pardey, P.G. Premiums and discounts for plant patents and trademarks used on ornamental plant cultivars: a hedonic price analysis. HortScience 2015, 50, 879–887. [Google Scholar] [CrossRef]
- Dominguez, G.B.; Mibus-Schoppe, H.; Sparke, K. Evaluation of existing research concerning sustainability in the value chain of ornamental plants. Eur. J. Sustain. Dev. 2017, 6, 11–19. [Google Scholar] [CrossRef]
- Guo, Y.; Starman, T.; Hall, C. Growth, quality, and economic value responses of bedding plants to reduced water usage. HortScience 2019, 54, 856–864. [Google Scholar] [CrossRef]
- Hall, T.J.; Dennis, J.H.; Lopez, R.G.; Marshall, M.I. Factors affecting growers’ willingness to adopt sustainable floriculture practices. HortScience 2009, 44, 1346–1351. [Google Scholar] [CrossRef]
- Lazzerini, G.; Merante, P.; Lucchetti, S.; Nicese, F.P. Assessing environmental sustainability of ornamental plant production: a nursery level approach in Pistoia District, Italy. Agroecol. Sust. Food 2018, 42, 911–932. [Google Scholar] [CrossRef]
- Cirillo, C.; Rouphael, Y.; Pannico, A.; El-Nakhel, C.; Colla, G.; De Pascale, S. Application of protein hydrolysate-based biostimulant as new approach to improve performance of bedding plants. Acta Hortic. 2018, 1215, 443–447. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Massa, D.; Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulant applications in low input horticultural cultivation systems. Italus Hortus 2018, 25, 27–36. [Google Scholar]
- Parađiković, N.; Teklić, T.; Zeljković, S.; Lisjak, M.; Špoljarević, M. Biostimulants research in some horticultural plant species—A review. Food Energy Secur. 2019, 8, e00162. [Google Scholar]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: a review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef]
- Cabrera, J.C.; Wégria, G.; Onderwater, R.C.A.; González, G.; Nápoles, M.C.; Falcón-Rodríguez, A.B.; Costales, D.; Rogers, H.J.; Diosdado, E.; González, S.; et al. Practical use of oligosaccharins in agriculture. Acta Hortic. 2013, 1009, 195–212. [Google Scholar] [CrossRef]
- Merino, D.; Casalongué, C.; Alvarez, V.A. Polysaccharides as eco-nanomaterials for agricultural applications. In Handbook of Ecomaterials; Torres-Martínez, L.M., Kharissova, O.V., Kharisov, B.I., Eds.; Springer Nature: Basel, Switzerland, 2018; pp. 2709–2730. [Google Scholar]
- Ahmed, K.B.M.; Khan, M.M.A.; Siddiqui, H.; Jahan, A. Chitosan and its oligosaccharides, a promising option for sustainable crop production-a review. Carbohydr. Polym. 2019, 17, 115331. [Google Scholar]
- Liu, J.; Yang, S.; Li, X.; Yan, Q.; Reaney, M.J.; Jiang, Z. Alginate oligosaccharides: Production, biological activities, and potential applications. Compr. Rev. Food Sci. F. 2019, 18, 1859–1881. [Google Scholar] [CrossRef]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Carrageenans from red seaweeds as promoters of growth and elicitors of defense response in plants. Front. Mar. Sci. 2016, 3, 81. [Google Scholar] [CrossRef]
- Ali, A.; Khan, M.M.A.; Uddin, M.; Naeem, M.; Idrees, M.; Hashmi, N.; Dar, T.A.; Varshney, L. Radiolytically depolymerized sodium alginate improves physiological activities, yield attributes and composition of essential oil of Eucalyptus citriodora Hook. Carbohydr. Polym. 2014, 112, 134–144. [Google Scholar] [CrossRef]
- El-Mohdy, H.L.A. Radiation-induced degradation of sodium alginate and its plant growth promotion effect. Arab. J. Chem. 2017, 10, 431–438. [Google Scholar] [CrossRef]
- Ahmad, B.; Jahan, A.; Sadiq, Y.; Shabbir, A.; Jaleel, H.; Khan, M.M.A. Radiation mediated molecular weight reduction and structural modification in carrageenan potentiates improved photosynthesis and secondary metabolism in peppermint (Mentha piperita L.). Int. J. Biol. Macromol. 2019, 124, 1069–1079. [Google Scholar] [CrossRef]
- Ahmed, K.B.M.; Khan, M.M.A.; Jahan, A.; Siddiqui, H.; Uddin, M. Gamma rays induced acquisition of structural modification in chitosan boosts photosynthetic machinery, enzymatic activities and essential oil production in citronella grass (Cymbopogon winterianus Jowitt). Int. J. Biol. Macromol. 2020, 145, 372–389. [Google Scholar] [CrossRef]
- Naeem, M.; Nabi, A.; Aftab, T.; Khan, M.M.A. Oligomers of carrageenan regulate functional activities and artemisinin production in Artemisia annua L. exposed to arsenic stress. Protoplasma 2019. [Google Scholar] [CrossRef]
- Dzung, P.D.; Phu, D.V.; Du, B.D.; Ngoc, L.S.; Duy, N.N.; Hiet, H.D.; Hien, N.Q. Effect of foliar application of oligochitosan with different molecular weight on growth promotion and fruit yield enhancement of chili plant. Plant Prod. Sci. 2017, 20, 389–395. [Google Scholar] [CrossRef]
- Muley, A.B.; Shingote, P.R.; Patil, A.P.; Dalvi, S.G.; Suprasanna, P. Gamma radiation degradation of chitosan for application in growth promotion and induction of stress tolerance in potato (Solanum tuberosum L.). Carbohydr. Polym. 2019, 210, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Muthukumar, T.; Song, J.E.; Khang, G. Biological role of gellan gum in improving scaffold drug delivery, cell adhesion properties for tissue engineering applications. Molecules 2019, 24, 4514. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Hosokawa, K.; Oomiya, T.; Yamamura, S. Plant regeneration from protoplasts of Gentiana by embedding protoplasts in gellan gum. Plant Cell Tiss.Org. 1995, 41, 221–227. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Smith, D.R. Maturation of somatic embryos of Pinus strobus is promoted by a high concentration of gellan gum. Physiol. Plant. 1997, 100, 949–957. [Google Scholar] [CrossRef]
- Lelu-Walter, M.A.; Gautier, F.; Eliášová, K.; Sanchez, L.; Teyssier, C.; Lomenech, A.M.; Le Metté, C.; Hargreaves, C.; Trontin, J.F.; Reeves, C. High gellan gum concentration and secondary somatic embryogenesis: Two key factors to improve somatic embryo development in Pseudotsuga menziesii Mirb. Plant Cell Tiss. Org. 2017, 132, 137–155. [Google Scholar] [CrossRef]
- Salachna, P.; Mizielińska, M.; Soból, M. Exopolysaccharide gellan gum and derived oligo-gellan enhance growth and antimicrobial activity in Eucomis plants. Polymers 2018, 10, 242. [Google Scholar] [CrossRef]
- Ollerton, J.; Lack, A. Relationships between flowering phenology, plant size and reproductive success in shape Lotus corniculatus (Fabaceae). Plant Ecol. 1998, 139, 35–47. [Google Scholar] [CrossRef]
- Bolmgren, K.D. Cowan, P. Time–size tradeoffs: A phylogenetic comparative study of flowering time, plant height and seed mass in a north-temperate flora. Oikos 2008, 117, 424–429. [Google Scholar] [CrossRef]
- Marfà, V.; Gollin, D.J.; Eberhard, S.; Mohnen, D.; Dan/ill, A.; Albersheim, P. Oligogalacturonides are able to induce flowers to form on tobacco explants. Plant J. 1991, 1, 217–225. [Google Scholar] [CrossRef]
- Darvill, A.; Augur, C.; Bergmann, C.; Carlson, R.W.; Cheong, J.J.; Eberhard, S.; Hahn, M.G.; Lo, V.M.; Marfa, V.; Meyer, B.; et al. Oligosaccharins- oligosaccharides that regulate growth, development and defence responses in plants. Glycobiology 1992, 2, 181–198. [Google Scholar] [CrossRef]
- John, M.; Röhrig, H.; Schmidt, J.; Walden, R.; Schell, J. Cell signalling by oligosaccharides. Trends Plant Sci. 1997, 2, 111–115. [Google Scholar] [CrossRef]
- Salachna, P.; Grzeszczuk, M.; Meller, E.; Mizielińska, M. Effects of gellan oligosaccharide and NaCl stress on growth, photosynthetic pigments, mineral composition, antioxidant capacity and antimicrobial activity in red perilla. Molecules 2019, 24, 3925. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.R.; Das, V.S.R. Correlation between biomass production and net photosynthetic rates and kinetic properties of RuBP carboxylase in certain C3 plants. Biomass 1986, 10, 157–164. [Google Scholar] [CrossRef]
- Chen, Z.; Tao, X.; Khan, A.; Tan, D.K.; Luo, H. Biomass accumulation, photosynthetic traits and root development of cotton as affected by irrigation and nitrogen-fertilization. Front. Plant Sci. 2018, 9, 173. [Google Scholar] [CrossRef] [PubMed]
- Roche, D. Stomatal conductance is essential for higher yield potential of C3 crops. Crit. Rev. Plant Sci. 2015, 34, 429–453. [Google Scholar] [CrossRef]
- Sun, L.; Wang, C.; Shi, Q.; Ma, C. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Yang, X.; Wang, J.; Li, Y.; Yu, H.; Zhang, Y.; Liu, G. Advances in characterisation and biological activities of chitosan and chitosan oligosaccharides. Food Chem. 2016, 190, 1174–1181. [Google Scholar] [CrossRef]
- Zhang, X.; Li, K.; Liu, S.; Zou, P.; Xing, R.; Yu, H.; Li, P. Relationship between the degree of polymerization of chitooligomers and their activity affecting the growth of wheat seedlings under salt stress. J. Agr. Food Chem. 2017, 65, 501–509. [Google Scholar] [CrossRef]
- Salachna, P.; Grzeszczuk, M.; Meller, E.; Soból, M. Oligo-alginate with low molecular mass improves growth and physiological activity of Eucomis autumnalis under salinity stress. Molecules 2018, 23, 812. [Google Scholar] [CrossRef]
- Zheng, F.; Chen, L.; Zhang, P.; Zhou, J.; Lu, X.; Tian, W. Carbohydrate polymers exhibit great potential as effective elicitors in organic agriculture: A review. Carbohydr. Polym. 2020, 230, 115637. [Google Scholar] [CrossRef]
- Boller, T.; Felix, G. A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Loo, E.P.I. Plant immunity in signal integration between biotic and abiotic stress responses. New Phytol. 2020, 225, 87–104. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liu, Y.; Liu, R. Sphingomonas sp.: An important microbial resource for biopolymer synthesis. Acta Microbiol. Sin. 2009, 49, 560–566. [Google Scholar]
- Luo, Y.; Wang, F.; Zhou, M.; Sheng, H.M. Sphingomonas sp. Cra20 increases plant growth rate and alters rhizosphere microbial community structure of Arabidopsis thaliana under drought stress. Front. Microbiol. 2019, 10, 1221. [Google Scholar] [CrossRef]
- Khan, A.L.; Waqas, M.; Kang, S.M.; Al-Harrasi, A.; Hussain, J.; Al-Rawahi, A.; Al-Khiziri, S.; Ullah, I.; Ali, L.; Jung, H.Y.; et al. Bacterial endophyte Sphingomonas sp. LK11 produces gibberellins and IAA and promotes tomato plant growth. J. Microbiol. 2014, 52, 689–695. [Google Scholar] [CrossRef]








© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salachna, P. Effects of Depolymerized Gellan with Different Molecular Weights on the Growth of Four Bedding Plant Species. Agronomy 2020, 10, 169. https://doi.org/10.3390/agronomy10020169
Salachna P. Effects of Depolymerized Gellan with Different Molecular Weights on the Growth of Four Bedding Plant Species. Agronomy. 2020; 10(2):169. https://doi.org/10.3390/agronomy10020169
Chicago/Turabian StyleSalachna, Piotr. 2020. "Effects of Depolymerized Gellan with Different Molecular Weights on the Growth of Four Bedding Plant Species" Agronomy 10, no. 2: 169. https://doi.org/10.3390/agronomy10020169
APA StyleSalachna, P. (2020). Effects of Depolymerized Gellan with Different Molecular Weights on the Growth of Four Bedding Plant Species. Agronomy, 10(2), 169. https://doi.org/10.3390/agronomy10020169