Salicylic Acid Improves Boron Toxicity Tolerance by Modulating the Physio-Biochemical Characteristics of Maize (Zea mays L.) at an Early Growth Stage


 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Root and Shoot Length
2.2. Plant Biomass
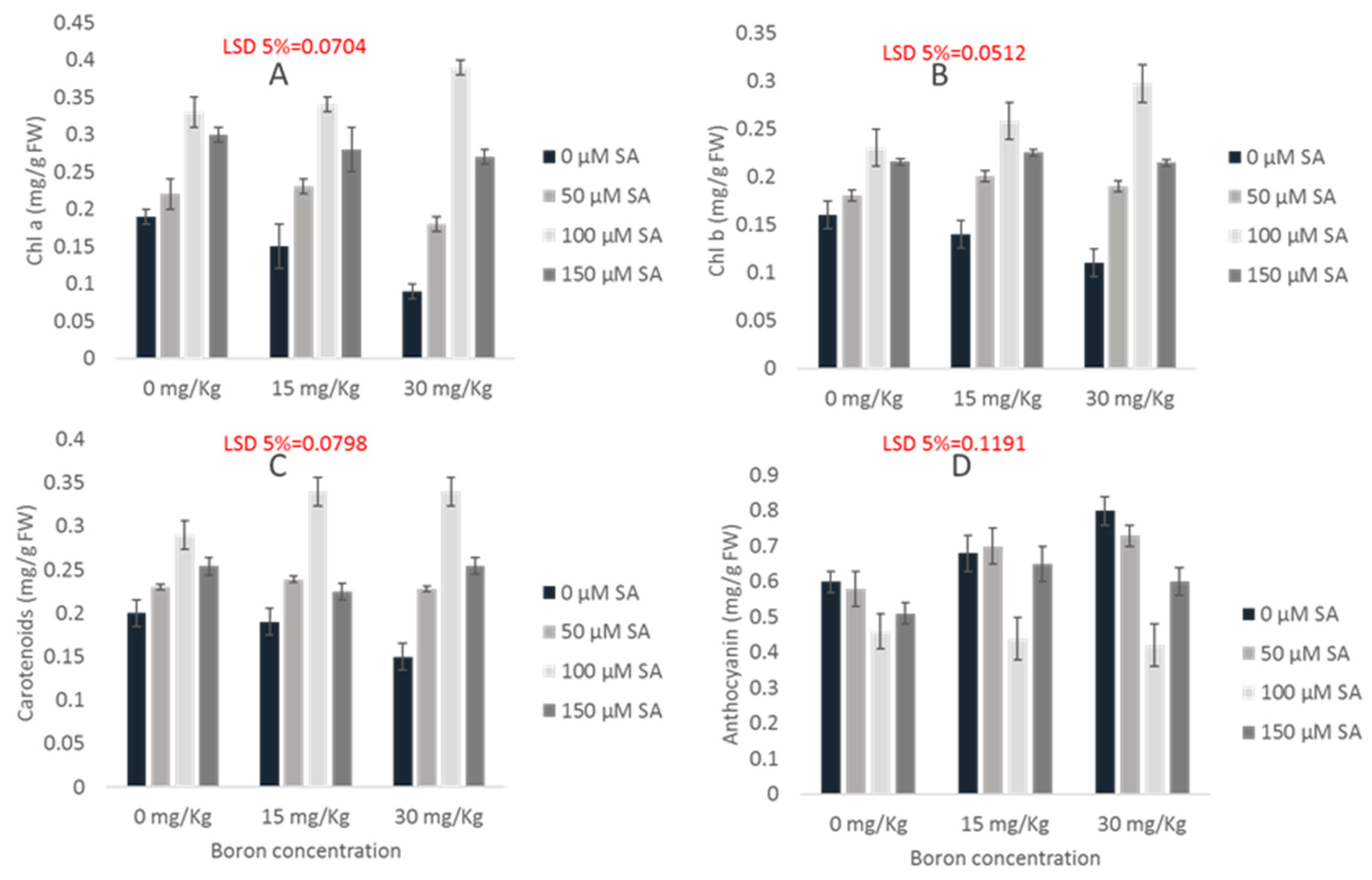
2.3. Photosynthetic Pigments
2.4. Anthocyanin
2.5. Ascorbic Acid
2.6. H2O2 Concentration
2.7. Proline Content
2.8. Glycine Betaine
2.9. Potassium Content
2.10. Calcium Content
2.11. Nitrate Concentration
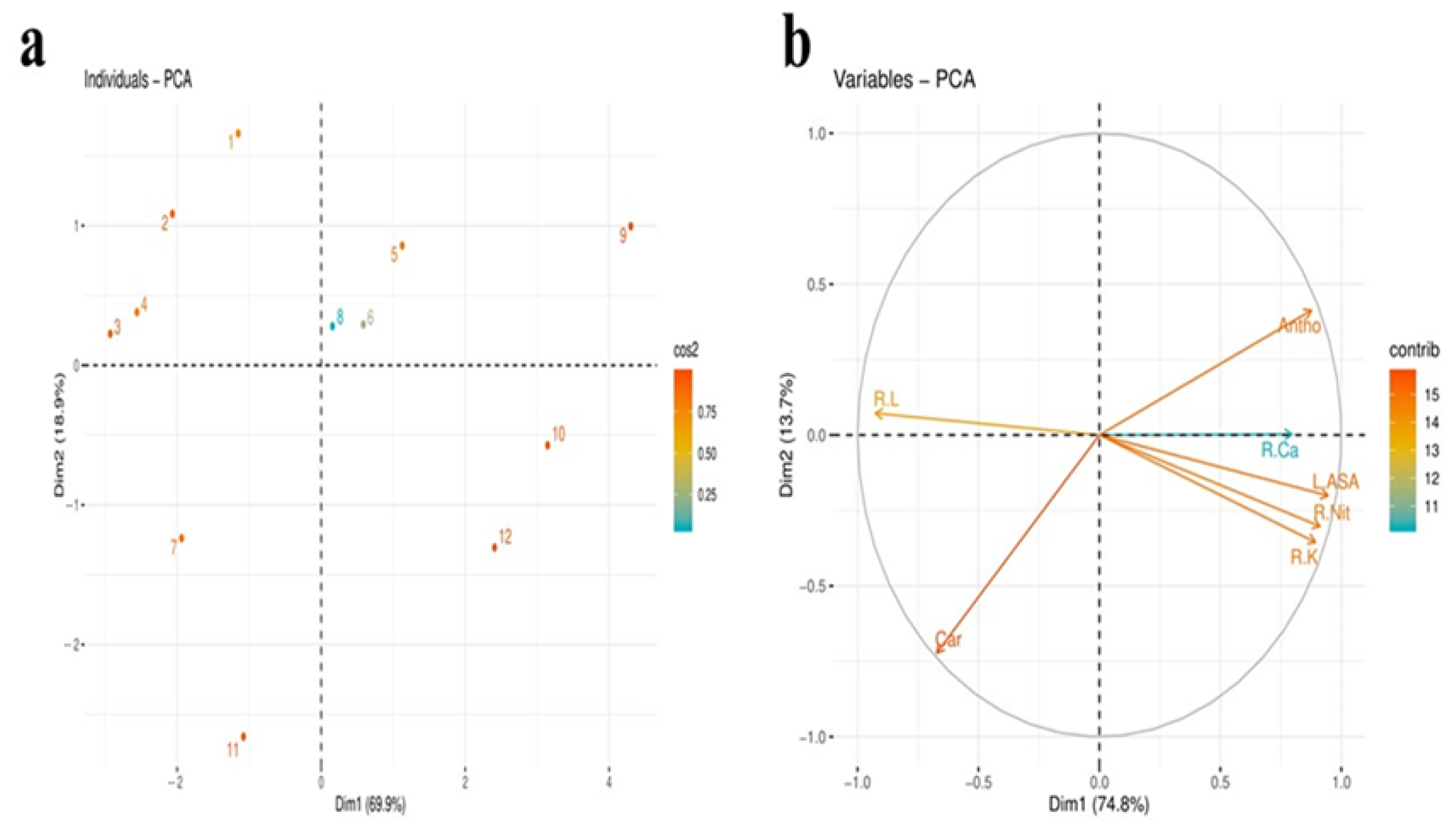
2.12. Principal Component Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Morphological Parameters and Plant Biomass
4.3. Physiological and Biochemical Analysis
4.3.1. Photosynthetic Pigments
4.3.2. Anthocyanin Content
4.3.3. Ascorbic Acid Content
4.3.4. H2O2 Content Determination
4.3.5. Potassium Content
4.3.6. Calcium Content
4.3.7. Nitrate Content
5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jaradat, A.; Simulated, A. Climate change deferentially impacts phenotypic plasticity and stoichiometric homeostasis in major food crops. Emir. J. Food Agr. 2018, 30, 429–442. [Google Scholar]
- Khan, N.; Ali, S.; Shahid, M.A.; Kharabian-Masouleh, A. Advances in detection of stress tolerance in plants through metabolomics approaches. Plant Omics 2017, 10, 153. [Google Scholar] [CrossRef]
- Mantri, N.; Patade, V.; Penna, S.; Ford, R.; Pang, E. Abiotic stress responses in plants: Present and future. In Abiotic Stress Responses in Plants; Springer: New York, NY, USA, 2012; pp. 1–19. [Google Scholar]
- Bakhsh, A. Engineering crop plants against abiotic stress: Current achievements and prospects. Emir. J. Food Agr. 2015, 27, 24–39. [Google Scholar] [CrossRef]
- Cheng, S. Heavy metal pollution in China: Origin, pattern and control. Environ. Sci. Pollut. Res. 2003, 10, 192–198. [Google Scholar] [CrossRef]
- Kelepertzis, E. Accumulation of heavy metals in agricultural soils of Mediterranean: Insights from Argolida basin, Peloponnese, Greece. Geoderma 2014, 221, 82–90. [Google Scholar] [CrossRef]
- Zhao, F.J.; Ma, Y.; Zhu, Y.G.; Tang, Z.; McGrath, S.P. Soil contamination in China: Current status and mitigation strategies. Environ. Sci. Technol. 2014, 49, 750–759. [Google Scholar] [CrossRef]
- Hussain, S.; Zhang, J.H.; Zhong, C.; Zhu, L.F.; Cao, X.C.; YU, S.M.; Jin, Q.Y. Effects of salt stress on rice growth, development characteristics and the regulating ways. J. Integr. Agric. 2017, 16, 2357–2374. [Google Scholar] [CrossRef]
- Liu, P.; Yang, Y.S.; Xu, G.D.; Fang, Y.H.; Yang, Y.A.; Kalin, R.M. The effect of molybdenum and boron in soil on the growth and photosynthesis of three soybean varieties. Plant Soil Environ. 2005, 51, 197–205. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Babar, M.A. The stimulatory effects of plant growth promoting rhizobacteria and plant growth regulators on wheat physiology grown in sandy soil. Arch. Microbiol. 2019, 201, 769–785. [Google Scholar] [CrossRef]
- Ayvaz, M.; Avcı, M.K.; Yamaner, C.; Koyuncu, M.; Güven, A.; Fagerstedt, K. Does excess boron affect the malondialdehyde levels of potato cultivars? Eurasia J. Biosci. 2013, 7, 47–53. [Google Scholar] [CrossRef]
- Stiles, A.R.; Liu, C.; Kayama, Y.; Wong, J.; Doner, H.; Funston, R.; Terry, N. Evaluation of the boron tolerant grass, Puccinellia distans, as an initial vegetative cover for the phytorestoration of a boron-contaminated mining site in southern California. Environ. Sci. Technol. 2011, 45, 8922–8927. [Google Scholar] [CrossRef] [PubMed]
- Princi, M.P.; Lupini, A.; Araniti, F.; Longo, C.; Mauceri, A.; Sunseri, F.; Abenavoli, M.R. Boron toxicity and tolerance in plants: Recent advances and future perspectives. In Plant Metal Interaction; Elsevier: Amsterdam, The Netherlands, 2016; pp. 115–147. [Google Scholar]
- Kayıhan, C.; Öz, M.T.; Eyidogan, F.; Yucel, M.; Oktem, H.A. Physiological, biochemical and transcriptomic responses to boron toxicity in leaf and root tissues of contrasting wheat cultivars. Plant Mol. Biol. Rep. 2017, 35, 97–109. [Google Scholar] [CrossRef]
- Yermiyahu, U.; Ben-Gal, A.; Keren, R.; Reid, R.J. Combined effect of salinity and excess boron on plant growth and yield. Plant Soil 2008, 304, 73–87. [Google Scholar] [CrossRef]
- Diaz, F.J.; Grattan, S.R. Performance of tall wheatgrass (Thinopyrum ponticum cv. Jose) irrigated with saline-high boron drainage water: Implications on ruminant mineral nutrition. Agric. Ecosyst. Environ. 2009, 131, 128–136. [Google Scholar] [CrossRef]
- Zhang, B.; Chu, G.; Wei, C.; Ye, J.; Li, Z.; Liang, Y. The growth and antioxidant defense responses of wheat seedlings to omethoate stress. Pestic. Biochem. Phys. 2011, 100, 273–279. [Google Scholar] [CrossRef]
- Khan, N.; Zandi, P.; Ali, S.; Mehmood, A.; Adnan Shahid, M.; Yang, J. Impact of salicylic acid and PGPR on the drought tolerance and phytoremediation potential of Helianthus annus. Front. Microbiol. 2018, 9, 2507. [Google Scholar] [CrossRef]
- Tavallali, V. Interactive effects of zinc and boron on growth, photosynthesis and water relations in pistachio. J. Plant Nutr. 2017, 40, 1588–1603. [Google Scholar] [CrossRef]
- Nasim, M.; Rengel, Z.; Aziz, T.; Regmi, B.D.; Saqib, M. Boron toxicity alleviation by zinc application in two barley cultivars differing in tolerance to boron toxicity. Pak. J. Agric. Sci. 2015, 52, 151–158. [Google Scholar]
- Samet, H.; Cikili, Y.; Dursun, S. The role of potassium in alleviating boron toxicity and combined effects on nutrient contents in pepper (Capsicum annuum L.). Bulg. J. Agric. Sci. 2015, 21, 64–70. [Google Scholar]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Sakran, A.M.; Ali, H.M.; Basalah, M.O.; Faisal, M.; Al-Amri, A.A. Calcium-induced amelioration of boron toxicity in radish. J. Plant Growth Regul. 2013, 32, 61–71. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Mohamed, H.I.; Mahmoud, A.; Elkelish, A.; Misra, A.N.; Guy, K.M.; Kamran, M.; Ai, S.; Zhang, M. Salicylic Acid Stimulates Antioxidant Defense and Osmolyte Metabolism to Alleviate Oxidative Stress in Watermelons under Excess Boron. Plants 2020, 9, 724. [Google Scholar] [CrossRef] [PubMed]
- Maghsoudia, K.; Arvinb, M.J. Salicylic acid and osmotic stress effects on seed germination and seedling growth of wheat (Triticum aestivum L.) cultivars. Plant Ecophysiol. 2010, 2, 7–11. [Google Scholar]
- El-Shazoly, R.M.; Metwally, A.A.; Hamada, A.H. Salicylic acid or thiamin increases tolerance to boron toxicity stress in wheat. J. Plant Nutr. 2019, 42, 702–722. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Zhang, Q.; Ashraf, U.; Anjum, S.A.; Ali, I.; Wang, L. Maize Tolerance against Drought and Chilling Stresses Varied with Root Morphology and Antioxidative Defense System. Plants 2020, 9, 720. [Google Scholar] [CrossRef] [PubMed]
- Aliyu, H.G.; Adamu, H.M. The potential of maize as phytoremediation tool of heavy metals. Eur. Sci. J. 2014, 6, 32–33. [Google Scholar]
- Wang, B.; Liu, C.; Zhang, D.; He, C.; Zhang, J.; Li, Z. Effects of maize organ-specific drought stress response on yields from transcriptome analysis. BMC Plant Biol. 2019, 19, 335. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. Effects of exogenously applied salicylic acid and putrescine alone and in combination with rhizobacteria on the phytoremediation of heavy metals and chickpea growth in sandy soil. Int. J. Phytoremediation 2018, 16, 405–414. [Google Scholar] [CrossRef]
- Brown, P.H.; Bellaloui, N.; Wimmer, M.A.; Bassil, E.S.; Ruiz, J.; Hu, H.; Pfeffer, H.; Dannel, F.; Römheld, V. Boron in plant biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Parks, J.L.; Edwards, M. Boron in the environment. Crit. Rev. Environ. Sci. Technol. 2005, 35, 81–114. [Google Scholar] [CrossRef]
- Blevins, D.G.; Lukaszewski, K.M. Boron in plant structure and function. Annu. Rev. Plant Biol. 1998, 49, 481–500. [Google Scholar] [CrossRef] [PubMed]
- Zafar-ul-Hye, M.; Munir, K.; Ahmad, M.; Imran, M. Influence of boron fertilization on growth and yield of wheat crop under salt stress environment. Soil Environ. 2016, 35, 181–186. [Google Scholar]
- Liu, D.; Jiang, W.; Zhang, L.; Li, L. Effects of boron ions on root growth and cell division of broad bean (Vicia Faba L.). Isr. J. Plant Sci. 2000, 48, 47–58. [Google Scholar] [CrossRef]
- Ghanati, F.; Morita, A.; Yokota, H. Deposition of suberin in roots of soybean induced by excess boron. Plant Sci. 2005, 168, 397–405. [Google Scholar] [CrossRef]
- Konuk, M.; Liman, R.; Cigerci, I.H. Determination of genotoxic effect of boron on Allium cepa root meristematic cells. Pak. J. Bot. 2007, 39, 73–79. [Google Scholar]
- Sarafi, E.; Tsouvaltzis, P.; Chatzissavvidis, C.; Siomos, A.; Therios, I. Melatonin and resveratrol reverse the toxic effect of high boron (B) and modulate biochemical parameters in pepper plants (Capsicum annuum L.). Plant Physiol. Biochem. 2017, 112, 173–182. [Google Scholar] [CrossRef]
- Eser, A.; Aydemir, T. The effect of kinetin on wheat seedlings exposed to boron. Plant Physiol. Biochem. 2016, 108, 158–164. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Ali, S.; Babar, M.A. Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stresses. Plant Growth Regul. 2020, 90, 189–203. [Google Scholar] [CrossRef]
- Chen, M.; Mishra, S.; Heckathorn, S.A.; Frantz, J.M.; Krause, C. Proteomic analysis of Arabidopsis thaliana leaves in response to acute boron deficiency and toxicity reveals effects on photosynthesis, carbohydrate metabolism and protein synthesis. J. Plant Physiol. 2013, 171, 235–242. [Google Scholar] [CrossRef]
- Turan, M.; Taban, N.; Taban, S. Effect of calcium on the alleviation of boron toxicity and localization of boron and calcium in cell wall of wheat. Not. Bot. Horti Agrobot. Cluj-Napoca 2009, 37, 99–103. [Google Scholar]
- Ghanati, F.; Morita, A.; Yokota, H. Induction of suberin and increase of lignin content by excess boron in tobacco cells. J. Plant Nutr. Soil Sci. 2002, 48, 357–364. [Google Scholar] [CrossRef]
- Sharma, A.; Sidhu, G.P.S.; Araniti, F.; Bali, A.S.; Shahzad, B.; Tripathi, D.K.; Brestic, M.; Skalicky, M.; Landi, M. The Role of Salicylic Acid in Plants Exposed to Heavy Metals. Molecules 2020, 25, 540. [Google Scholar] [CrossRef] [PubMed]
- Papadakis, I.; Dimassi, K.; Bosabalidis, A.; Therios, I.; Patakas, A.; Giannakoula, A. Boron toxicity in ‘Clementine’ mandarin plants grafted on two root stocks. Plant Sci. 2004, 166, 539–547. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M. Exogenous application of nitric oxide promotes growth and oxidative defense system in highly boron stressed tomato plants bearing fruit. Sci. Hortic. 2015, 185, 43–47. [Google Scholar] [CrossRef]
- Pardossi, A.; Romani, M.; Carmassi, G.; Guidi, L.; Landi, M.; Incrocci, L.; Maggini, R.; Puccinelli, M.; Vacca, W.; Ziliani, M. Boron accumulation and tolerance in sweet basil (Ocimum basilicum L.) with green or purple leaves. Plant Soil 2015, 395, 375–389. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef]
- Qian, H.F.; Peng, X.F.; Han, X.; Ren, J.; Zhan, K.Y.; Zhu, M. The stress factor, exogenous ascorbic acid, affects plant growth and the antioxidant system in Arabidopsis thaliana. Russ. J. Plant Physiol. 2014, 61, 467–475. [Google Scholar] [CrossRef]
- Karabal, E.; Yucel, M.; Huseyin, A.O. Antioxidant responses of tolerant and sensitive barley cultivars to boron toxicity. Plant Sci. 2003, 164, 925–933. [Google Scholar] [CrossRef]
- Gunes, A.; Soylemezoglu, G.; Inal, A.; Bagci, E.G.; Coban, S.; Sahin, O. Antioxidant and stomatal responses of grapevine (Vitis vinifera L.) to boron toxicity. Sci. Hortic. 2006, 110, 279–284. [Google Scholar] [CrossRef]
- Cervilla, L.M.; Blasco, B.; Rıos, R.; Romero, L.; Ruiz, J. Oxidative stress and antioxidants in tomato (Solanum lycopericum) plants subjected to boron toxicity. Ann. Bot. 2007, 100, 747–756. [Google Scholar] [CrossRef]
- Ardic, M.; Sekmen, A.H.; Tokur, S.; Ozdemir, F.; Turkan, I. Antioxidant responses of chickpea plants subjected to boron toxicity. Plant Biol. 2009, 11, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bano, A.; Curá, J.A. Role of Beneficial Microorganisms and Salicylic Acid in Improving Rainfed Agriculture and Future Food Safety. Microorganisms 2020, 8, 1018. [Google Scholar] [CrossRef] [PubMed]
- Shaki, F.; Maboud, H.E.; Niknam, V. Effects of salicylic acid on hormonal cross talk, fatty acids profile, and ions homeostasis from salt-stressed safflower. J. Plant Interact. 2019, 14, 340–346. [Google Scholar] [CrossRef]
- Ahmad, P.; Abdel Latef, A.A.; Hashem, A.; Abd-Allah, E.F.; Gucel, S.; Tran, L.S.P. Nitric oxide mitigates salt stress by regulating levels of osmolytes and antioxidant enzymes in chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hoque, M.A.; Burritt, D.J.; Fujita, M. Proline Protects Plants against Abiotic Oxidative Stress: Biochemical and Molecular Mechanisms. In Oxidative Damage to Plants; Academic Press: Cambridge, MA, USA, 2014; pp. 477–522. [Google Scholar]
- El-Esawi, M.A.; Elkelish, A.; Soliman, M.; Elansary, H.O.; Zaid, A.; Wani, S.H. Serratia marcescens BM1 Enhances Cadmium Stress Tolerance and Phytoremediation Potential of Soybean Through Modulation of Osmolytes, Leaf Gas Exchange, Antioxidant Machinery, and Stress-Responsive Genes Expression. Antioxidants 2020, 9, 43. [Google Scholar] [CrossRef]
- Abdelaal, K.A.; EL-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.I.; El-Banna, M.; El-Esawi, M.; Elkelish, A. Treatment of Sweet Pepper with Stress Tolerance-Inducing Compounds Alleviates Salinity Stress Oxidative Damage by Mediating the Physio-Biochemical Activities and Antioxidant Systems. Agronomy 2020, 10, 26. [Google Scholar] [CrossRef]
- Alhaithloul, H.A.; Soliman, M.H.; Ameta, K.L.; El-Esawi, M.A.; Elkelish, A. Changes in Ecophysiology, Osmolytes, and Secondary Metabolites of the Medicinal Plants of Mentha piperita and Catharanthus roseus Subjected to Drought and Heat Stress. Biomolecules 2020, 10, 43. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.M.; Ali, H.M.; Alayafi, A.A.; Ahmad, M. Serratia liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-Related Gene Expression. Int. J. Mol. Sci. 2018, 19, 3310. [Google Scholar] [CrossRef]
- Zafar-ul-Hye, M.; Naeem, M.; Danish, S.; Khan, M.J.; Fahad, S.; Datta, R.; Brtnicky, M.; Kintl, A.; Hussain, G.S.; El-Esawi, M.A. Effect of Cadmium-Tolerant Rhizobacteria on Growth Attributes and Chlorophyll Contents of Bitter Gourd under Cadmium Toxicity. Plants 2020, 9, 1386. [Google Scholar] [CrossRef]
- Naveed, M.; Bukhari, S.S.; Mustafa, A.; Ditta, A.; Alamri, S.; El-Esawi, M.A.; Rafique, M.; Ashraf, S.; Siddiqui, M.H. Mitigation of Nickel Toxicity and Growth Promotion in Sesame through the Application of a Bacterial Endophyte and Zeolite in Nickel Contaminated Soil. Int. J. Environ. Res. Public Health 2020, 17, 8859. [Google Scholar] [CrossRef]
- Imran, M.; Hussain, S.; El-Esawi, M.A.; Rana, M.S.; Saleem, M.H.; Riaz, M.; Ashraf, U.; Potcho, M.P.; Duan, M.; Rajput, I.A.; et al. Molybdenum Supply Alleviates the Cadmium Toxicity in Fragrant Rice by Modulating Oxidative Stress and Antioxidant Gene Expression. Biomolecules 2020, 10, 1582. [Google Scholar] [CrossRef] [PubMed]
- Ali, Q.; Shahid, S.; Ali, S.; El-Esawi, M.A.; Hussain, A.I.; Perveen, R.; Iqbal, N.; Rizwan, M.; Nasser Alyemeni, M.; El-Serehy, H.A.; et al. Fertigation of Ajwain (Trachyspermum ammi L.) with Fe-Glutamate Confers Better Plant Performance and Drought Tolerance in Comparison with FeSO4. Sustainability 2020, 12, 7119. [Google Scholar] [CrossRef]
- Soliman, M.; Alhaithloul, H.A.; Hakeem, K.R.; Alharbi, B.M.; El-Esawi, M.; Elkelish, A. Exogenous Nitric Oxide Mitigates Nickel-Induced Oxidative Damage in Eggplant by Upregulating Antioxidants, Osmolyte Metabolism, and Glyoxalase Systems. Plants 2019, 8, 562. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Stark, D.; Wray, V. Anthocyanins. In Methods in Plant Biology; Volume 1: Plant Phenolics; Harborne, J.B., Ed.; Academic Press: London, UK; Harcourt Brace Jovanovich: London, UK, 1989; pp. 325–356. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Root Length (cm) | ||||
| 0 mg kg−1 B | 15 mg kg−1 B | 30 mg kg−1 B | ||
| SA | 0 µM | 27.1 ± 0.89 c | 24.3 ± 1.05 b | 21.2 ± 0.88 c |
| 50 µM | 27.8 ± 1.01 b | 24.2 ± 0.97 b | 21.34 ± 1.03 c | |
| 100 µM | 29 ± 0.98 a | 28.2 ± 0.87 a | 26.4 ± 0.77 a | |
| 150 µM | 26.4 ± 1.12 d | 23 ± 1.24 c | 21.8 ± 1.02 b | |
| Shoot Length (cm) | ||||
| SA | 0 µM | 30.3 ± 0.69 b | 28.5 ± 0.85 b | 22.65 ± 1.25 c |
| 50 µM | 30.2 ± 1.13 b | 28.7 ± 0.77 b | 22.10 ± 1.02 d | |
| 100 µM | 32.0 ± 0.99 a | 30.5 ± 0.98 a | 29.00 ± 0.84 a | |
| 150 µM | 28.0 ± 1.21 c | 27.4 ± 0.66 c | 27.00 ± 0.96 b | |
| Treatment | Root Fresh Weight (g) | Root Dry Weight (g) | Shoot Fresh Weight (g) | Shoot Dry Weight (g) |
|---|---|---|---|---|
| 00 µM SA + 00 mg/g−1 B | 1.00 ± 0.89 abc | 0.75 ± 0.21 bc | 3.80 ± 0.33 abc | 2.53 ± 1.01 abcd |
| 00 µM SA + 15 mg kg−1 B | 0.89 ± 0.95 bc | 0.65 ± 0.34 d | 3.10 ± 0.41 abc | 2.07 ± 0.85 abcd |
| 00 µM SA + 30 mg kg−1 B | 0.70 ± 0.55 c | 0.5 ± 0.21 d | 2.55 ± 0.55 c | 1.70 ± 0.33 d |
| 50 µM SA + 00 mg kg−1 B | 1.25 ± 0.45 ab | 1.02 ± 0.35 ab | 3.85 ± 1.01 abc | 2.57 ± 0.65 abcd |
| 50 µM SA + 15 mg kg−1 B | 1.15 ± 0.75 abc | 0.95 ± 0.34 ab | 3.00 ± 0.95 abc | 2.00 ± 0.35 abcd |
| 50 µM SA + 30 mg kg−1 B | 0.85 ± 0.65 bc | 0.62 ± 0.32 d | 2.70 ± 0.55 bc | 1.80 ± 0.45 cd |
| 100 µM SA + 00 mg kg−1 B | 1.50 ± 0.76 ab | 1.26 ± 0.22 a | 4.30 ± 0.25 a | 2.87 ± 0.27 a |
| 100 µM SA + 15 mg kg−1 B | 1.35 ± 0.55 ab | 1.09 ± 0.36 abc | 3.60 ± 0.97 abc | 2.40 ± 0.85 abc |
| 100 µM SA + 30 mg kg−1 B | 1.15 ± 0.75 abc | 0.9 ± 0.23 abc | 3.00 ± 0.85 abc | 2.00 ± 0.33 abcd |
| 150 µM SA + 00 mg kg−1B | 1.25 ± 0.82 ab | 0.99 ± 0.45 abc | 3.90 ± 0.21 abc | 2.60 ± 0.43 abc |
| 150 µM SA + 15 mg kg−1 B | 1.00 ± 071 abc | 0.75 ± 0.35 bc | 3.25 + 0.85 abc | 2.17 ± 0.55 abcd |
| 150 µM SA + 30 mg kg−1 B | 0.95 ± 0.66 bc | 0.71 ± 0.32 c | 2.85 ± 0.79 abc | 1.90 ± 0.65 bcd |
| LSD 5% | 0.51 | 0.49 | 1.46 | 0.98 |
| Treatment | Leaf ASA (µmoles/g FW) | Leaf H2O2 (mg/g FW) | Leaf Proline (µMole/g FW) | Leaf GB (µg/g FW) |
|---|---|---|---|---|
| 00 µM SA + 00 mg kg−1 B | 210 ± 1.53 | 0.80 ± 0.15 | 32.00 ± 0.5 | 1.60 ± 0.05 |
| 00 µM SA + 15 mg kg−1 B | 320 ± 1.15 | 1.80 ± 0.06 | 36.50 ± 0.5 | 1.80 ± 0.03 |
| 00 µM SA + 30 mg kg−1 B | 375 ± 0.58 | 2.50 ± 0.10 | 46.00 ± 0.5 | 1.90 ± 0.03 |
| 50 µM SA + 00 mg kg−1 B | 209 ± 1.00 | 0.70 ± 0.05 | 33.00 ± 0.5 | 1.70 ± 0.05 |
| 50 µM SA + 15 mg kg−1 B | 300 ± 0.58 | 1.34 ± 0.03 | 37.00 ± 0.58 | 2.50 ± 0.06 |
| 50 µM SA + 30 mg kg−1 B | 360 ± 0.58 | 2.40 ± 0.03 | 47.00 ± 0.50 | 2.50 ± 0.08 |
| 100 µM SA + 00 mg kg−1 B | 200 ± 1.00 | 0.64 ± 0.02 | 33.00 ± 0.29 | 1.80 ± 0.05 |
| 100 µM SA + 15 mg kg−1 B | 260 ± 0.58 | 1.00 ± 0.03 | 46.00 ± 0.76 | 2.00 ± 0.05 |
| 100 µM SA + 30 mg kg−1 B | 240 ± 0.58 | 1.90 ± 0.03 | 58.00 ± 0.29 | 2.80 ± 0.05 |
| 150 µM SA + 00 mg kg−1 B | 211 ± 1.15 | 0.78 ± 0.02 | 37.00 ± 0.29 | 1.90 ± 0.05 |
| 150 µM SA + 15 mg kg−1 B | 276 ± 0.76 | 1.45 ± 0.05 | 37.00 ± 0.58 | 2.50 ± 0.09 |
| 150 µM SA + 30 mg kg−1 B | 335 ± 0.29 | 2.20 ± 0.20 | 48.00 ± 0.29 | 2.40 ± 0.08 |
| LSD 5% | 0.51 | 0.49 | 1.46 | 0.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawaz, M.; Ishaq, S.; Ishaq, H.; Khan, N.; Iqbal, N.; Ali, S.; Rizwan, M.; Alsahli, A.A.; Alyemeni, M.N. Salicylic Acid Improves Boron Toxicity Tolerance by Modulating the Physio-Biochemical Characteristics of Maize (Zea mays L.) at an Early Growth Stage. Agronomy 2020, 10, 2013. https://doi.org/10.3390/agronomy10122013
Nawaz M, Ishaq S, Ishaq H, Khan N, Iqbal N, Ali S, Rizwan M, Alsahli AA, Alyemeni MN. Salicylic Acid Improves Boron Toxicity Tolerance by Modulating the Physio-Biochemical Characteristics of Maize (Zea mays L.) at an Early Growth Stage. Agronomy. 2020; 10(12):2013. https://doi.org/10.3390/agronomy10122013
Chicago/Turabian StyleNawaz, Muhammad, Sabtain Ishaq, Hasnain Ishaq, Naeem Khan, Naeem Iqbal, Shafaqat Ali, Muhammad Rizwan, Abdulaziz Abdullah Alsahli, and Mohammed Nasser Alyemeni. 2020. "Salicylic Acid Improves Boron Toxicity Tolerance by Modulating the Physio-Biochemical Characteristics of Maize (Zea mays L.) at an Early Growth Stage" Agronomy 10, no. 12: 2013. https://doi.org/10.3390/agronomy10122013
APA StyleNawaz, M., Ishaq, S., Ishaq, H., Khan, N., Iqbal, N., Ali, S., Rizwan, M., Alsahli, A. A., & Alyemeni, M. N. (2020). Salicylic Acid Improves Boron Toxicity Tolerance by Modulating the Physio-Biochemical Characteristics of Maize (Zea mays L.) at an Early Growth Stage. Agronomy, 10(12), 2013. https://doi.org/10.3390/agronomy10122013