Abstract
Developing new crops adapted to arid conditions is a promising approach to meet the increasing demand for food production under expanding aridity. Tef [Eragrostis tef (Zucc.) Trotter] is a C4 cereal crop cultivated mainly in Ethiopia and the Horn of Africa, and known for its high resilience to stressful environments. Due to their tiny size, tef seeds are traditionally sown by broadcasting and lightly covering with soil. Under semiarid Mediterranean conditions, a deeper sowing may guarantee seedling establishment while saving on irrigation water. The objective of this study was, therefore, to determine the effects of sowing depth on tef emergence, development, lodging, and productivity. Tef seeds were sown at different depths in test tubes and pots, and in two field experiments. In tubes and pots, time from sowing to emergence increased about twofold and emergence rate decreased in the deepest sowing (3 cm) treatment compared to controls (0 cm). In the pot and field trials, deep sowing (3 cm) significantly reduced plant height, shoot and root biomass, and lodging. Sowing depths of 1–2 cm allowed successful plant establishment while not exacting penalties on plant emergence or development; hence this range appears to be optimal for sowing irrigated tef.
1. Introduction
Climate change toward higher temperatures, greater aridity, and more frequent erratic climate events is posing a major threat to agricultural sustainability [1]. The development of new crops adapted to arid conditions is among the promising approaches to meet the projected demand for food production under expanding aridity. The introduction of a crop into a new region requires comprehensive studies of the crop’s responses to environmental factors. The adoption of a crop from a traditional rain-fed system in an intensive, mechanized, and irrigated cropping system is particularly challenging, requiring examination and adjustment of management practices, from field preparation, sowing, irrigation, mineral nutrition, and plant protection, to harvesting techniques.
Tef [Eragrostis tef (Zucc.) Trotter] is a cereal crop that is cultivated mainly in Ethiopia and the Horn of Africa [2]. Tef has a thin culm, long narrow leaves, and a thousand-kernel weight of 200–400 mg, making it the smallest-grained cereal [3]. Recently, tef has been gaining popularity in many Western countries due to its dual potential—as a gluten-free “super food” grain [3], and as high-quality forage [4,5]. In Israel, the demand for tef grain is further increased by the Ethiopian descent citizens who continue to consume it for their traditional cuisine [6].
Tef has a C4 photosynthetic pathway, which allows for efficient utilization of high solar radiation. It is highly tolerant to various stressful environments, such as marginal soils, water-logging, salinity, and drought [7,8,9]. In addition to tef’s remarkable resilience, its ability to produce grain and/or fodder in a relatively short growing season (60–90 days) [6] enables drought escape and water savings. Thus, tef offers great advantages for arid region cultivation and holds promise for expansion of its production to currently uncultivated lands.
One of the basic details in mechanical farming is sowing depth. As a general rule of thumb, seeds should not be sown deeper than five times their width [10]. Tef’s tiny seed size, 0.4–0.7 mm width and 1–1.4 mm length, hence calls for a maximum sowing depth of 2–3 mm. Tef is traditionally sown in Ethiopia by manual seed broadcasting and lightly covering with soil to prevent drying out [11,12], probably due to limited access to agricultural machinery. In addition, local farmers believe that the seeds are so small that they will not emerge from greater depths [13]. However, under hot and dry conditions, deeper sowing is recommended to guarantee seedling establishment [14]. In Oregon, USA, the recommended sowing depth for tef is between 3 mm and 15 mm [5]. Sowing tef at 0 mm or at greater than 15 mm depth had negative effects on plant height and seedling emergence [13,14,15], with no seeds emerging from a sowing depth of 50 mm or greater [13].
Based on our previous study [16] and published data [4], tef is sown in Israel during the spring (March–April), at the onset of the hot and dry summer. Rainfall in Israel in March, and even more so in April, is very limited and scattered, and therefore irrigation is essential to securing proper germination and crop establishment. Shallow sowing would require up to two irrigation applications per day to avoid seed desiccation, which can sum to 150 mm during its 10–15 days of establishment, about half of the seasonal water requirement of the crop. Therefore, deeper sowing might be key to saving water during the crop’s establishment. Optimal sowing depth should provide a sufficiently thick soil layer to maintain the humidity required for germination, but not be so thick as to confer excessive mechanical resistance to the seedling’s emergence. Knowledge of the effects of sowing depth is also necessary for the development of advanced agricultural practices, including—but not limited to—the choice of sowing machinery.
Plant lodging, defined as the permanent displacement of the stem from the vertical, which is caused by environmental conditions as well as morphological plant traits [17], is the most significant yield-reducing problem in tef, accounting for up to 35% of yield losses [18,19]. While certain studies relate tef lodging to stem characteristics (stem lodging) [20], others [18], including our recently published paper [21], point to shoot–root junction traits (root lodging) as major contributors to tef lodging. In wheat, deeper sowing has been shown to induce a longer, deeper root crown, thus improving plant anchorage and reducing root lodging [17] (and references therein). Hence, reduced lodging could be an additional benefit of deeper sowing in tef. The objective of this study was, therefore, to determine the effects of sowing depth on tef plant emergence, development, lodging, and productivity.
2. Materials and Methods
2.1. Plant Material
Six tef genotypes representing the range of thousand-seed weights were selected for the current study from the diversity panel assembled in our laboratory [6]. These genotypes consisted of three seed-size groups: small, medium, and large, with each size group including a brown-seeded and a white-seeded genotype (Table 1).

Table 1.
Characteristics of genotypes used in this study.
2.2. Test-Tube Experiment
The effects of sowing depth on seedling emergence and establishment were studied in a test-tube experiment for all six genotypes. Seeds were sown in 50 mL test tubes filled with brown-red sandy loam soil composed of 76% sand, 8% silt, and 16% clay (Figure 1).

Figure 1.
Tef seedlings sown in 50 mL tubes at depths of (left to right) 0, 1, 2, 3 cm 144 h after sowing.
A hole was drilled at the bottom of each tube to allow drainage and covered with a thin layer of fabric to avoid soil leakage. A factorial experimental design was employed with the six genotypes, each sown at four depths: 0 cm (control) with 1–2 mm soil coverage, 1 cm, 2 cm, and 3 cm. Test tubes were filled with dry soil to the designated level, watered, and 20 seeds from a single genotype were scattered on the soil surface of each tube. To ensure an accurate sowing depth, preweighed soil was added to match a soil layer of 1, 2, and 3 cm in the 50 mL test tube, while the 0 cm treatment seeds were gently mixed into the top 1–2 mm of soil. After sowing, tubes were wrapped with aluminum foil to avoid root exposure to light. A total of 24 tubes—one tube for each of the six genotypes and four sowing depths—were sown in each of five growing cycles, referred to as experimental blocks.
The tubes were placed in a climate-controlled greenhouse at the Faculty of Agriculture, Food and Environment campus in Rehovot, Israel, under temperatures of 28/22 °C day/night, and were watered twice daily by microsprinklers. At the end of the growth cycle, after all plants reached the second leaf stage, plants were uprooted, and the soil was gently washed away to observe the root system.
The number of emerged seedlings and number of seedlings that reached the second leaf stage were recorded daily for each test tube. Time from sowing to emergence (TSE), time from sowing to fully expanded second leaf (TSL2), and time from emergence to second leaf (TEL2) were calculated as follows:
where n is the number of seedlings emerged at each counting, t is the time in hours from sowing to counting, and N is the total number of seedlings emerged.
where s is the number of seedlings with a fully unfolded second leaf at each counting, t is the time in hours from sowing to counting, and S is the total number of seedlings that reached the second leaf stage.
TEL2 = TSL2 − TSE
2.3. Pot Experiment
Three genotypes, RTC-119 (large seeds), RTC-400 (medium seeds), and RTC-19 (small seeds), were selected for the pot experiment (Table 1). To enable accurate sowing depth, transparent cylindrical pots (8.2 cm diameter, 18 cm height), made from plastic soft-drink bottles, were used for this experiment. Holes were drilled at the bottom of each pot to allow drainage and covered with a thin layer of fabric to avoid soil leakage. A factorial experimental design was employed with three genotypes, each sown at four depths (0 cm (control), 1 cm, 2 cm, and 3 cm), and six replicates (a total of 72 pots). The type of soil used, number of seeds per experimental unit, sowing methodology, and scoring of seedling emergence and second leaf stage were as in the test-tube experiment. Once all seedlings reached the second leaf stage, all but one randomly selected seedling were removed from each bottle and the remaining plant continued to grow until harvest at flowering onset. Just before harvest, the lodging angle (between the main culm and the vertical) was measured. The plants were then uprooted, the soil was washed away, and number of tillers, crown diameter, number and diameter of crown roots, and shoot and root dry weight were assessed following the protocols in Ben-Zeev et al. [21].
2.4. Field Experiments
Two field experiments were conducted in the summers of two consecutive years, 2019 and 2020, in two locations, the Hula Valley in Northern Israel (33°06′46.8” N, 35°35′03.7” E) and Revadim in Central Israel (31°46′04.8″ N, 34°49′02.8″ E), respectively. Soil type at the Hula experimental site was deep peat (52% sand, 43% silt, 5% clay, and ~10% organic matter) and at the Revadim site clay soil (clay 41%, sand 42%, silt 17%, and ~1% organic matter). A factorial block design was employed in both experiments with 2 genotypes (RTC-119 and 400) × 2 sowing depths × 12 replicates in Hula (2019), and 3 genotypes (RTC-19, 119 and 400) × 2 sowing depths × 5 replicates in Revadim (2020).
Soil preparation included shallow tilling and flattening with a heavy-duty leveler and crumble roller tool to obtain a smooth and even seedbed. Seeds were tested to confirm uniform germination rates and were treated with Vitavax® fungicide (thiram + carboxin, Gadot Argo, Israel) at a rate of 100 µL/10 g seeds. Experimental plots were mechanically sown using a Plotseed S seeder (Wintersteiger, Ried, Austria). Ten rows were sown in each 8 m × 1.6 m plot at two sowing depths (1 or 3 cm) using a sowing rate of 6 kg/ha (Hula) or 4 kg/ha (Revadim).
Seedling density was assessed 10 days after emergence. A metal frame with inner dimensions of 25 cm × 100 cm was placed across the seedling rows in two locations per plot and the number of seedlings within this frame area was counted.
Lodging was assessed visually by two independent surveyors twice a week, starting from six weeks after emergence (WAE), using a scoring method adapted from Caldicott and Nuttall [22]. Lodging was scored on a 10-level severity scale (0 being a vertical nonlodging plant and 9 being a horizontal fully lodging plant), and lodging prevalence was determined (percentage of the entire plot area). Lodging index was calculated as the product of lodging severity × lodging score.
Plant height and panicle length assessments and harvest were performed at grain maturity, 12 WAE. Three plants, randomly selected from various parts of each plot, were measured from the soil surface to the bottom and top of the panicle and used to calculate the average culm length and panicle length, respectively. A metal frame of 25 cm × 100 cm was placed in the middle of the plot across six rows (excluding the two outer rows from each side) and all of the biomass bordered by the frame was manually harvested and collected into paper bags. Biomass samples were dried in a hot glasshouse (maximum temperature ~55 °C) for two weeks, weighed for total dry matter, and threshed using a LD350 thresher (Wintersteiger, Ried, Austria). Seed samples were cleaned of debris and weighed to determine grain yield (GY).
2.5. Statistical Analysis
Statistical analysis was conducted using JMP® Pro, Version 15, software (SAS Institute Inc., Cary, NC, USA, 1989–2019), and included two-way analysis of variance (ANOVA), Student t test, and Tukey HSD test (for factors showing a significant F ratio).
3. Results
3.1. Test-Tube Experiment: Seedling Emergence and Establishment
The six tef genotypes, sown at four depths (0, 1, 2, 3 cm) in test tubes, presented highly significant effects of both sowing depth and genotype on average TSE and emergence rate, with no significant interaction between the two (Table 2).
Table 2.
Test-tube experiment. ANOVA for average time from sowing to emergence (TSE), emergence rate, time from emergence to second leaf (TEL2), and time from sowing to second leaf (TSL2).
TSE increased twofold from 55 h at the shallowest sowing depth (0 cm) to 109 h at the deepest sowing depth (3 cm) (Figure 2A); the other treatments increased in similar increments across this range, with all treatments differing significantly from one another. TEL2 exhibited the opposite trend, decreasing from 134 h at the shallowest sowing depth to 104 h at the deepest sowing (Figure 2A). This trend compensated for a substantial part of the delay in emergence from deeper sowings, and therefore TSL2 ranged between 189 h for the shallowest treatment to 214 h for the deepest treatment. Emergence rates recorded at 0 and 1 cm depths were 47%, with no apparent difference between them. Emergence rate was significantly reduced to 33% in the 3 cm treatment, whereas the 2 cm depth exhibited intermediate values, not significantly different from the other treatments (Figure 2B). The delayed emergence and lower emergence rate induced by deeper sowing can be clearly visualized in Figure 1 and Figure 3.
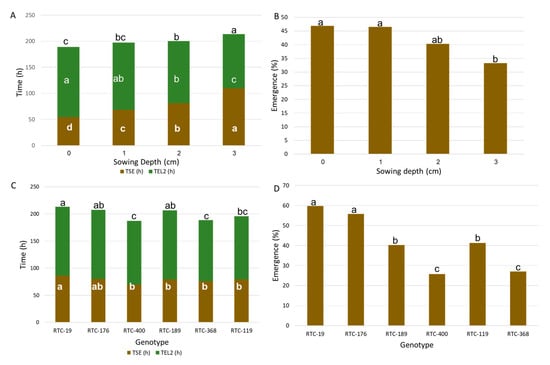
Figure 2.
Test-tube experiment. Effects of sowing depth (A) and genotype (C) on average time from sowing to emergence (TSE, brown segment), time from emergence to second leaf (TEL2, green segment), and time from sowing to second leaf (TSL2, both segments combined). Effects of sowing depth (B) and genotype (D) on average emergence rate. Different letters indicate significant differences (p < 0.05).

Figure 3.
Tef seedlings sown at different depths (0, 1, 2, 3 cm). Seed locations (S) and mesocotyl (M) are marked by white arrows. Picture was taken upon appearance of fully expanded second leaf on the plant sown at 3 cm depth.
The six genotypes tested in this study presented a rather uniform TSE, with only RTC-19 showing a significantly higher value (slower emergence) compared to most other genotypes (Figure 2C). There were no significant differences between genotypes for TEL2, hence TSL2 of the various genotypes exhibited a trend that was rather similar to that of TSE. An interesting trend was observed with the emergence rates: the small-seeded genotypes (RTC-176 and 19) exhibited significantly higher emergence rates (56–60%) (Figure 2D) than the medium- and large-seeded genotypes (26–42%), with no consistent trend among the latter. However, the number of genotypes tested in this study is too small to extrapolate from these data to a relationship between seed size and emergence rate.
3.2. Pot Experiment: Emergence and Plant Performance
The three tef genotypes sown in pots at 0, 1, 2, and 3 cm depths exhibited significant effects of both sowing depth and genotype on TSE and emergence rate (Table 3).
Table 3.
Pot experiment. ANOVA for average time from sowing to emergence (TSE), emergence rate, plant height, and shoot, root and total biomass.
TSE increased significantly with every increase in sowing depth from 0 to 3 cm (Figure 4A), similar to the test-tube experiment. Emergence rate was significantly reduced at the deepest sowing depth compared to the other depths (Figure 4B). Genotype-by-sowing depth interaction was of low significance for TSE and nonsignificant for emergence rate (Table 3); whereas TSE for all three genotypes responded similarly to increased sowing depth (Figure 4C), emergence rate of RTC-19 and 119 decreased with sowing depth, while no effect was recorded for RTC-400 (Figure 4D).
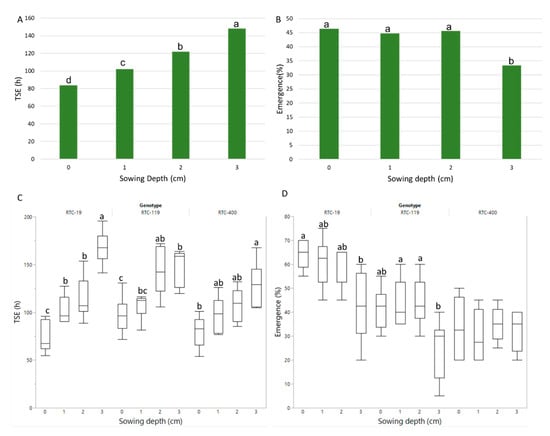
Figure 4.
Pot experiment. Effects of sowing depth on (A) average time from sowing to emergence (TSE) and (B) emergence rate. TSE (C) and emergence rate (D) for each genotype and sowing depth. Different letters indicate significant differences (p < 0.05) between sowing depths (on average and within each genotype).
Sowing depth significantly affected plant height and shoot and root biomass, and thus also the root-to-shoot weight ratio. Plant height ranged from 45.5 to 65.5 cm for RTC-19 at 3 cm and RTC-400 at 0 cm, respectively, and was generally reduced as sowing depth increased, with the exception of RTC-119 (Figure 5). Root and shoot biomass (range 0.28–0.6 g and 0.2–0.39 g, respectively) decreased with increasing sowing depth. Root-to-shoot ratio generally increased with increasing sowing depth, with RTC-119 as the only exception (data not shown).
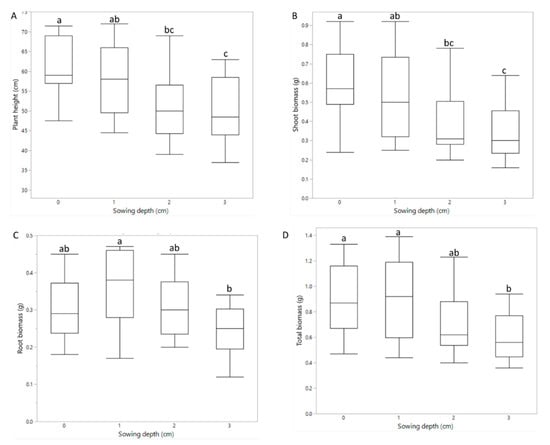
Figure 5.
Pot experiment. Effects of sowing depth on (A) plant height, (B) shoot biomass, (C) root biomass, and (D) total biomass. Different letters indicate significant differences (p < 0.05).
3.3. Pot Experiment: Lodging-Related Traits
Lodging-related traits, which included lodging angle, number of internodes and tillers, crown diameter, and crown root diameter and number, were assessed 12 weeks after sowing. ANOVA for lodging-related traits revealed significant effects of both sowing depth and genotype on the number of internodes, and significant effects of genotype on root diameter, whereas sowing depth-by-genotype interactions were not significant (Table 4). Crown root number and tiller number were not affected by sowing depth, genotype, or their interaction.
Table 4.
Pot experiment. ANOVA and main effect of sowing depth and genotypes on lodging and related traits.
Sowing depth had a moderate effect on lodging angle (angle between the main culm and the vertical), which fell below the statistical threshold by ANOVA, but did show significant differences between treatments by Tukey HSD test. The average lodging angle for the 3 cm treatment (40°) was significantly smaller (lower lodging) than for the 0 cm treatment (62°) (Table 4). Another variable affected by sowing depth was the number of internodes, which decreased incrementally with the increase in sowing depth, resulting in a significant difference between the 0 cm treatment and both 2 and 3 cm treatments (Table 4).
The tested genotypes did not have a significant effect on lodging angle. Significant differences between genotypes were manifested in three other lodging-related traits, with RTC-400 showing a significantly higher number of internodes and significantly lower crown and root diameters, compared to the two other genotypes, which did not differ from one another (Table 4).
3.4. Field Experiments: Emergence and Plant Performance
Two and three genotypes were mechanically sown in the field to depths of 1 or 3 cm in the 2019 and 2020 seasons, respectively. Sowing depth had a significant effect on number of seedlings/m2 and plant height in both years (Table 5), whereas panicle length and GY were only affected in 2019. The genotypes had significant effects on GY in both years, and on the number of seedlings/m2 in 2020.
Table 5.
ANOVA and main effects of sowing depth and genotypes on agronomic traits measured in two field experiments in 2019 (top) and 2020 (bottom).
Deeper sowing reduced seedling density by about 15% in both years (Table 5), and reduced plant height by 25% in 2019 and 7% in 2020. In 2019, panicle length and GY under the 3 cm sowing treatment were 21% and 39% lower, respectively, than for the 1 cm depth treatment, whereas no such effects were evident in 2020.
Seedling number did not differ between genotypes in 2019; however, in 2020, RTC-19 (not included in the 2019 trial) had a significantly greater number of seedlings than the other two genotypes (Table 5). This result corresponded with the greater germination rate of RTC-19 in our test-tube experiment (Figure 2). GY was significantly affected by genotype, with no consistent trends across years; RTC-119 exhibited the highest GY in 2019 and the lowest in 2020 (Table 5).
3.5. Field Experiments: Lodging
The weekly lodging index value, based on two evaluations per week, was subjected to ANOVA. In 2019, lodging indices at 7 and 8 WAE were significantly affected by sowing depth; the genotype effect was significant throughout 7–10 WAE and depth-by-genotype interactions were not significant (Table 6). In 2020, lodging index was not significantly affected by sowing depth, genotype effect was significant only at 8 WAE, and depth-by-genotype interactions were significant at 7 and 10 WAE.
Table 6.
ANOVA and main effects of sowing depth and genotypes on lodging indices (LI) assessed in two field experiments in 2019 (top) and 2020 (bottom) during weeks 7–10.
The whole-season analysis showed a significant effect of time (Week) and sowing depth in both years, a significant effect of genotype in 2019, and significant depth-by-genotype interactions in 2020 (Table 6). A plot of lodging indices vs. WAE showed a similar trend in both years, with the deep sowing treatment exhibiting lower lodging 7 and 8 WAE, whereas at 9 and 10 WAE, this advantage was diminished (Figure 6).
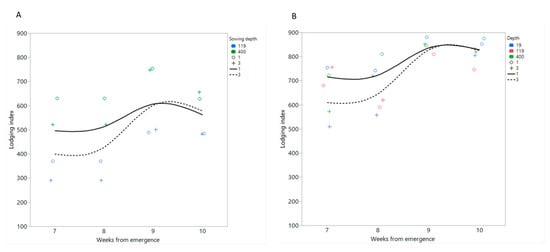
Figure 6.
Field experiment. Effects of sowing depth (+, dashed line: 3 cm; o, full line: 1 cm) on seasonal trends of lodging index of three tef genotypes (RTC119–red, RTC 400—green, and RTC-19—blue) in (A) 2019 and (B) 2020.
4. Discussion
4.1. Emergence Time and Rate
All three experimental setups presented similar trends: deeper sowing of tef resulted in later emergence and lower emergence rates in test tubes (Figure 1, Figure 2A,B and Figure 3), pots (Figure 4A,B), and field trials (Table 5). A sowing depth of 1 cm led to a slight but significant increase in TSE in both test tubes and pots. Larger effects on germination were recorded at a sowing depth of 2 cm, and the greatest effects were evident at a sowing depth of 3 cm (Figure 2A and Figure 4A). In both the test-tube and pot experiments, TSE increased incrementally with increasing sowing depth, reflecting the longer distance from the seed to the soil surface (Figure 2A and Figure 4A). TEL2, measured only in the test-tube experiment, exhibited a slight decrease with increasing sowing depth, thus partially compensating for the effect of sowing depth on TSE (Figure 2A). A rather clear breaking point can be observed with respect to germination rate, with a significant reduction for the 3 cm depth treatment in both test tubes (Figure 2B) and pots (Figure 4B), which was confirmed in the two field trials (Table 5). These trends were, for the most part, uniform across genotypes, in both the test-tube experiment (data not shown) and pot experiments (Figure 4C,D).
These results suggest that sowing tef at 1 or 2 cm depth has no or only minor effects on TSE or TSL2, whereas sowing at 3 cm depth has significant negative effects on these variables. In wheat (Triticum spp.) and barley (Hordeum vulgare), increasing sowing depths along the ranges of 0.8–14.3 cm [23] and 5–20 cm [24], respectively, reduced emergence rate and increased TSE. Previous studies on tef, all conducted in pots, have reported a reduction in the rate of and time to germination for sowing depths of 0 or greater than 2 cm [5,13,14,15,18]. Our test-tube and pot experiments are generally in agreement with these studies, although we did not observe a reduction in the 0 cm sowing treatment and did observe an increase in TSE at 2 cm sowing. These results were further validated in our two field experiments in two different locations with different soil types. We are not aware of any prior study on tef sowing depth conducted under field conditions.
4.2. Plant Development and Performance
In other tropical grasses, such as maize (Zea mays L.) and sorghum (Sorghum bicolor L.), the mesocotyl elongates until the coleoptile is exposed to light [25], thus locating the apical meristem at a depth corresponding to the coleoptile length, which is associated with seed size [26]. Being the smallest-seeded cereal [3], tef has a short coleoptile which locates the apical meristem just below the soil surface. Indeed, regardless of sowing depth, the root–shoot junction of tef seedlings for all treatments was located, upon emergence, just below the soil surface (Figure 3). Nevertheless, plants sown at 2 and 3 cm depths developed into shorter, lower biomass plants in both pots and field trials, with shorter panicles and a lower GY (2019 field trial) than plants in the shallower sowing treatments (Figure 5, Table 3 and Table 5). These differences in shoot traits, manifested at a later plant age, can be attributed to a penalty incurred from the deeper sowing. Similar penalties have been reported for wheat and barley [23,24], including a reduction in number of leaves and internode lengths, plant height, tiller number (no significant effect found in the present study), biomass, and GY. Confounding trends have been reported for sorghum, with a nonsignificant effect on plant biomass and a significant increase in plant height at deeper sowing depth [27].
4.3. Effect of Sowing Depth on Lodging and Related Traits
Lodging is a major yield-restricting factor in tef [18,19]. Deeper sowing has been shown to reduce root lodging in wheat [17], possibly due to the formation of a deeper root plate [28], which improves plant anchoring to the soil. In our pot experiment, deeper sowing of tef induced a significant effect on number of internodes, whereas its effects on lodging angle, crown diameter, and crown root diameter fell below the statistical threshold (Table 4). In our previous study, lodging of four genotypes grown in the field under various plant densities was associated with crown diameter, crown root diameter, and crown root number [21]; in the pot experiment, crown diameter and crown root diameter showed similar effects.
In both field trials, sowing tef seeds at 3 cm depth significantly reduced the seasonal average lodging index compared to 1 cm sowing depth (Table 6), with a particularly pronounced effect at 7–8 WAE (Figure 6). The effect of sowing depth on tef lodging has not been reported in previous studies. Recently, we reported that lower seedling density reduces lodging index [21]. Published studies on tef [20,29] and other cereals [28] have shown an association between lower plant height and reduced lodging. Both seedling density and plant height were reduced by the deeper sowing in the field (Table 5). Therefore, we cannot rule out the possibility that these two factors contributed to the reduced lodging observed in our field trials. However, the significant increase in root-to-shoot ratio (not presented) and reduced number of internodes, as well as the nearly significant modifications in crown and crown root diameters in the pot experiment, suggest that additional mechanisms contribute to reduced lodging under the deeper sowing treatment in the field.
5. Conclusions
This study was motivated by the need to reduce the amount of irrigation water required for tef establishment under semiarid Mediterranean conditions. An optimal sowing depth should allow sufficient soil above the seed to maintain the required moisture for germination, while not inflicting penalties on seedling emergence or plant development. Despite the tiny size of tef seeds, which was thought to require a very shallow sowing depth, we showed that tef can successfully emerge from a depth of 1–2 cm without any apparent penalty. Tef even emerged from 3 cm depth, while showing reduced lodging, but this was coupled with a penalty on plant development and productivity. Therefore, we conclude that a 1–2 cm depth is optimal for sowing irrigated tef. Further research combining optimal sowing depth, sowing rate, and irrigation regimes with a wide genotypic range to evaluate plant development, lodging and productivity, could lead to advances in tef production in Mediterranean and other similar regions around the globe.
Author Contributions
Conceptualization, S.B.-Z. and Y.S.; formal analysis, S.B.-Z.; funding acquisition, Y.S.; investigation, S.B.-Z., S.K., O.R.; project administration, Y.S.; supervision, Y.S.; writing—original draft, S.B.-Z.; writing—review and editing, S.K., O.R., and Y.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by The Israel Ministry of Agriculture and Rural Development, Chief Scientist Foundation (grant nos. 21-01-0045 and 12-01-0032).
Acknowledgments
We thank Vered Barak, Neta Levinson, Noa Kirby, Ori Harash, and Eran Dagan for their technical assistance in the field and laboratory. We also thank the Tsabar Kama agriculture partnership for hosting the field experiments. S.B.-Z. is indebted to the Robert H. Smith Foundation for a doctoral fellowship award. Y.S. is the incumbent of the Haim Gvati Chair in Agriculture.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sillmann, J.; Kharin, V.V.; Zwiers, F.W.; Zhang, X.; Bronaugh, D. Climate extremes indices in the CMIP5 multimodel ensemble: Part 2. Future climate projections. J. Geophys. Res. Atmos. 2013, 118, 2473–2493. [Google Scholar] [CrossRef]
- Blösch, R.; Plaza-Wüthrich, S.; Barbier de Reuille, P.; Weichert, A.; Routier-Kierzkowska, A.-L.; Cannarozzi, G.; Robinson, S.; Tadele, Z. Panicle angle is an important factor in tef lodging tolerance. Front. Plant Sci. 2020, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Tatham, A.S.; Fido, R.J.; Moore, C.M.; Kasarda, D.D.; Kuzmicky, D.D.; Keen, J.N.; Shewry, P.R. Characterisation of the major prolamins of tef (Eragrostis tef) and finger millet (Eleusine coracana). J. Cereal Sci. 1996, 24, 65–71. [Google Scholar] [CrossRef]
- Miller, D. Teff Grass: Crop Overview and Forage Production Guide. Available online: https://kingsagriseeds.com/wp-content/uploads/2014/12/Teff-Grass-Management-Guide.pdf (accessed on 18 November 2020).
- Norberg, S.; Roseberg, R.; Charlton, B.; Shock, C. Teff: A New Warm-season Annual Grass for Oregon. Oregon State University Extension Catalogue 2009. Available online: https://catalog.extension.oregonstate.edu/sites/catalog/files/project/pdf/em8970.pdf (accessed on 18 November 2020).
- Ben-Zeev, S.; Bimro, J.; Barak, V.; Saranga, Y. Phenotypic diversity and heritability in Eragrostis tef under irrigated Mediterranean conditions. Isr. J. Plant Sci. 2018, 65, 222–231. [Google Scholar] [CrossRef]
- Ketema, S. Tef (Eragrostis tef): Breeding, Genetic Resources, Agronomy, Utilization and Role in Ethiopian Agriculture; Institute of Agricultural Research: Adis Ababa, Ethopia, 1993.
- Stallknecht, G.F.; Gilbertson, K.M.; Eckhoff, J.L. Teff: Food crop for humans and animals. In New Crops; Janick, J., Simon, J.E., Eds.; Wiley: New York, NY, USA, 1993; pp. 231–234. [Google Scholar]
- Gelaw, A.M.; Qureshi, A.S. Tef (Eragrostis tef): A superfood grain from Ethiopia with great potential as an alternative crop for marginal environments. In Emerging Research in Alternative Crops; Hirich, A., Choukr-Allah, R., Ragab, R., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 265–278. [Google Scholar]
- Government of Western Australia. Monitoring Sowing Depth. Available online: https://www.agric.wa.gov.au/mycrop/monitoring-sowing-depth (accessed on 18 November 2020).
- Ketema, S. Eragrostis tef (Zucc.) Trotter: Promoting the Conservation and Use of Underutilized and Neglected Crops; Institute of Plant Genetics and Crop Plant Research: Gatersleben, Germany; International Plant Genetic Resources Institute: Rome, Italy, 1997; ISBN 9290433043. [Google Scholar]
- Mihretie, F.; Tsunekawa, A.; Bitew, Y.; Cheklie, G.; Derebe, B.; Getahun, W.; Beshir, O.; Tadesse, Z.; Asfaw, M. Teff [Eragrostis tef (Zucc.)] rain-fed yield response to planting method, seeding density, and row spacing. Agron. J. 2020. [Google Scholar] [CrossRef]
- Evert, S.; Staggenborg, S.; Olson, B.L.S. Soil temperature and planting depth effects on tef emergence. J. Agron. Crop Sci. 2009, 195, 232–236. [Google Scholar] [CrossRef]
- Debelo, A. Germination, Yield, and Yield Components of Tef (Eragrostis tef (Zucc.) Trotter) as Affected by Environment, Tillage and Weed Control Practices; Oklahoma State University: Stillwater, OK, USA, 1992. [Google Scholar]
- Paff, K.; Asseng, S. A review of tef physiology for developing a tef crop model. Eur. J. Agron. 2018, 94, 54–66. [Google Scholar] [CrossRef]
- Bimro, J. Agronomic Aspects of Teff (Eragrostis tef) Cultivation in Israel; The Hebrew University of Jerusalem: Rehovot, Israel, 2016. [Google Scholar]
- Pinthus, M.J. Lodging in wheat, barley, and oats: The phenomenon, its causes, and preventive measures. Adv. Agron. 1974, 25, 209–263. [Google Scholar]
- Van Delden, S.H.; Vos, J.; Ennos, A.R.; Stomph, T.J. Analysing lodging of the panicle bearing cereal teff (Eragrostis tef). New Phytol. 2010, 186, 696–707. [Google Scholar] [CrossRef] [PubMed]
- Ketema, S. Studies of Lodging Floral Biology and Breeding Techniques in Tef (Eragrostis tef); University of London: London, UK, 1983. [Google Scholar]
- Bayable, M.; Tsunekawa, A.; Haregeweyn, N.; Ishii, T.; Alemayehu, G.; Tsubo, M.; Adgo, E.; Tassew, A.; Tsuji, W.; Asaregew, F.; et al. Biomechanical properties and agro-morphological traits for improved lodging resistance in Ethiopian teff (Eragrostis tef (Zucc.) Trottor) accessions. Agronomy 2020, 10, 1012. [Google Scholar] [CrossRef]
- Ben-Zeev, S.; Rabinovitz, O.; Orlov-Levin, V.; Chen, A.; Graff, N.; Goldwasser, Y.; Saranga, Y. Less Is more: Lower sowing rate of irrigated tef (Eragrostis tef) alters plant morphology and reduces lodging. Agronomy 2020, 10, 570. [Google Scholar] [CrossRef]
- Caldicott, J.J.B.; Nuttall, A.M. A method for the assessment of lodging in cereal crops. J. Natl. Inst. Agric. Bot. 1979, 15, 88–91. [Google Scholar]
- Kirby, E.J.M. Effect of sowing depth on seedling emergence, growth and development in barley and wheat. Field Crops Res. 1993, 35, 101–111. [Google Scholar] [CrossRef]
- Photiades, I.; Hadjichristodoulou, A. Sowing date, sowing depth, seed rate and row spacing of wheat and barley under dryland conditions. Field Crops Res. 1984, 9, 151–162. [Google Scholar] [CrossRef]
- Hanway, J. How a Corn Plant Develops; Special Report 48; Iowa State University: Ames, IA, USA, 1966; Volume 48, pp. 1–18. [Google Scholar]
- Cornish, P.S.; Hindmarsh, S. Seed size influences the coleoptile length of wheat. Aust. J. Exp. Agric. 1988, 28, 521–523. [Google Scholar] [CrossRef]
- Harris, D. The effects of manure, genotype, seed priming, depth and date of sowing on the emergence and early growth Sorghum bicolor (L.) Moench in semi-arid Botswana. Soil Tillage Res. 1996, 40, 73–88. [Google Scholar]
- Berry, P.M.; Sterling, M.; Spink, J.H.; Baker, C.J.; Sylvester-Bradley, R.; Mooney, S.J.; Tams, A.R.; Ennos, A.R. Understanding and reducing lodging in cereals. Adv. Agron. 2004, 84, 217–271. [Google Scholar]
- Josst, M.; Esfeld, K.; Burian, A.; Cannarozzi, G.; Chanyalew, S.; Kuhlemeier, C.; Assefa, K.; Tadele, Z. Semi-dwarfism and lodging tolerance in tef (Eragrostis tef) is linked to a mutation in the alpha-Tubulin 1 gene. J. Exp. Bot. 2015, 66, 933–944. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).