Climate Change and the Herbicide Paradigm: Visiting the Future

{kind=link}
{kind=link}
Abstract
1. Introduction
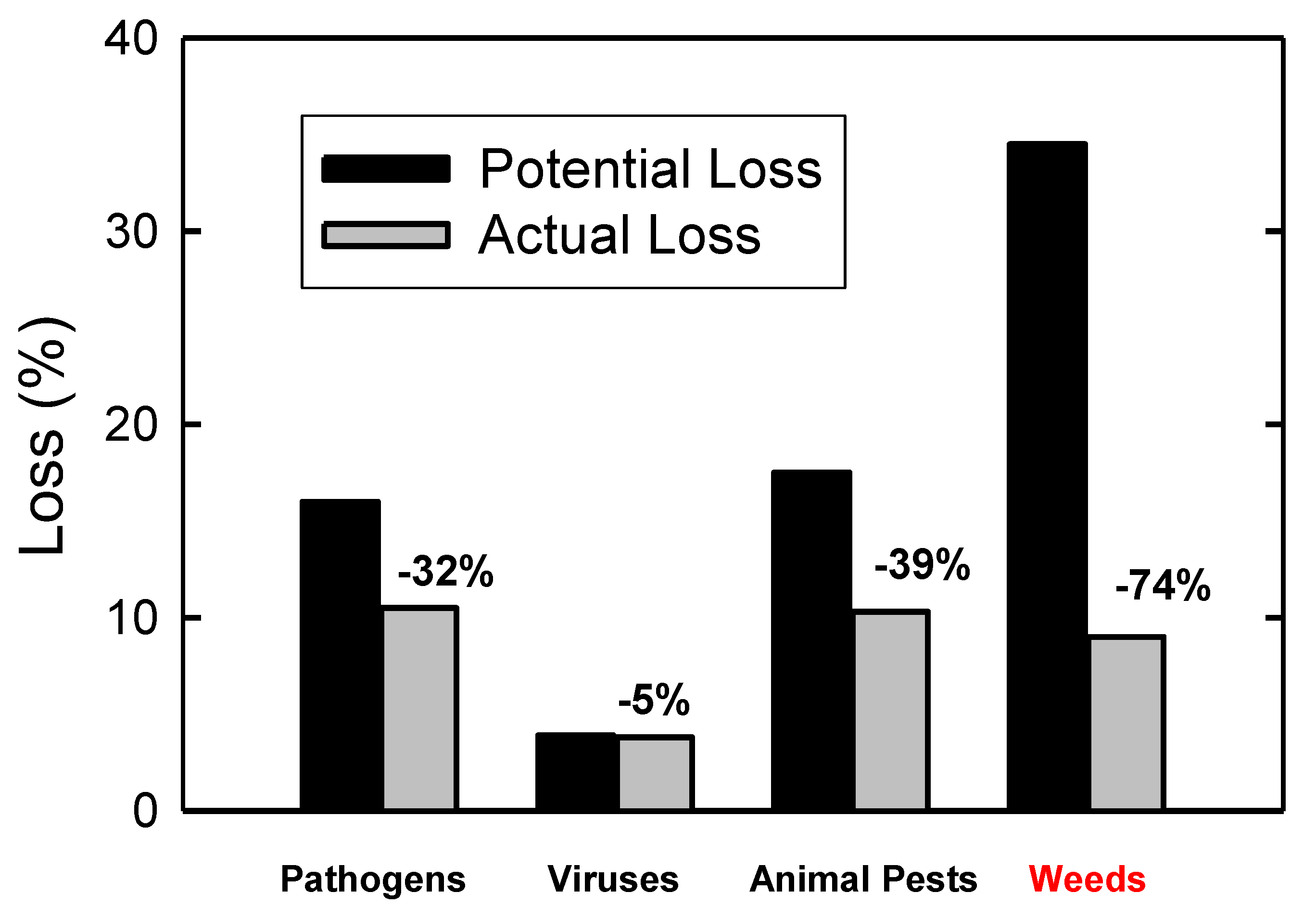
2. Weed Management
3. The Glyphosate Paradigm
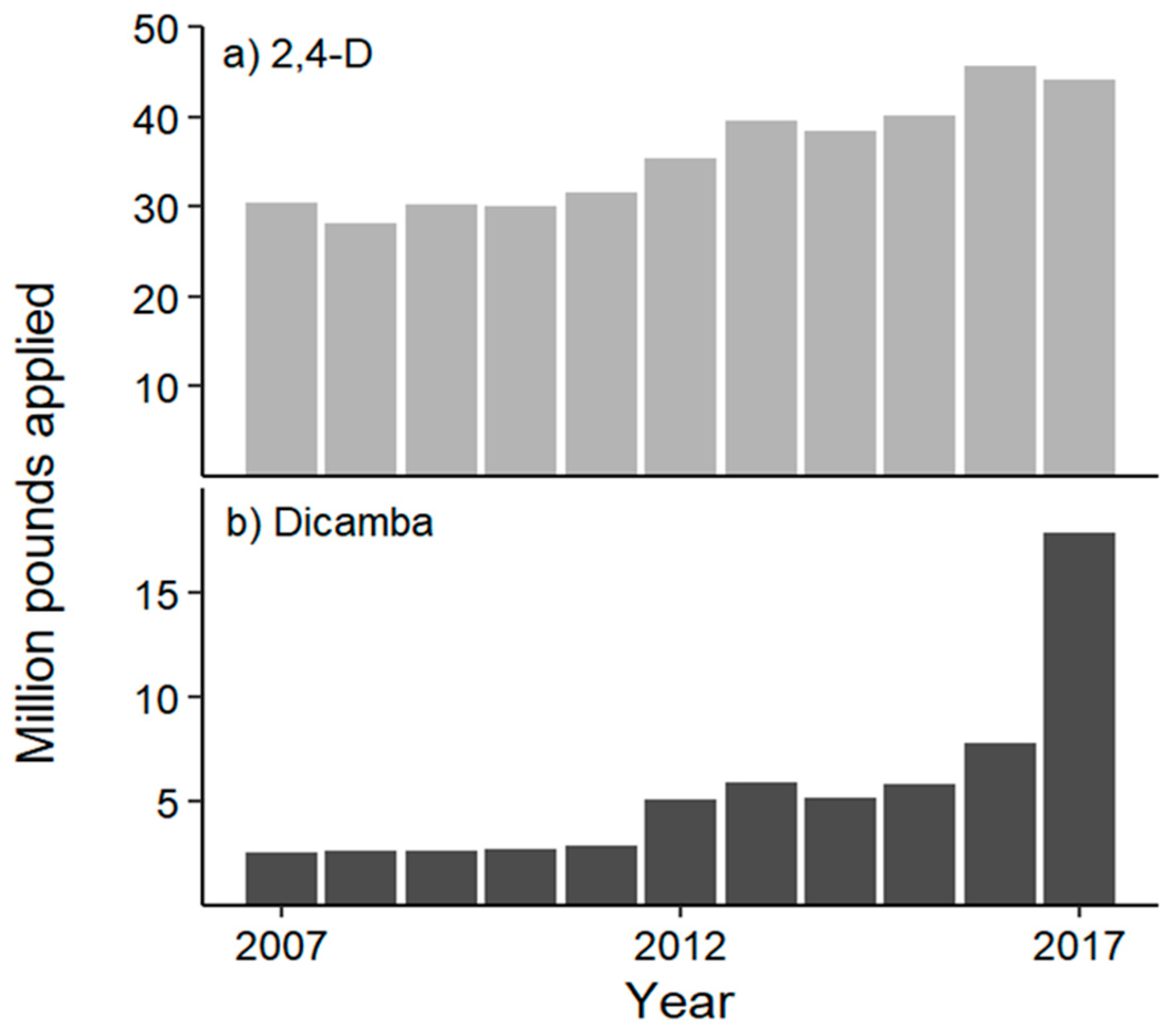
4. A Shifting Dynamic
5. CO2, Climate: New Evolutionary Pressures
6. A New Paradigm?
7. Further Challenges
8. Final Thoughts
Funding
Acknowledgments
Conflicts of Interest
References
- United Nations Population Estimates. Available online: https://www.un.org/development/desa/en/news/population/world-population-prospects-2019.html (accessed on 13 November 2020).
- Qian, B.; Zhang, X.; Smith, W.; Grant, B.; Jing, Q.; Cannon, A.J.; Neilsen, D.; McConkey, B.; Li, G.; Bonsal, B.; et al. Climate change impacts on Canadian yields of spring wheat, canola and maize for global warming levels of 1.5 C, 2.0 C, 2.5 C and 3.0 C. Environ. Res. Lett. 2019, 14, 074005. [Google Scholar] [CrossRef]
- Lobell, D.B.; Asseng, S. Comparing estimates of climate change impacts from process-based and statistical crop models. Environ. Res. Lett. 2017, 12, 015001. [Google Scholar] [CrossRef]
- Rosenzweig, C.; Parry, M.L. Potential impact of climate change on world food supply. Nature 2004, 367, 133–138. [Google Scholar] [CrossRef]
- Lobell, D.B.; Burke, M.B.; Tebaldi, C.; Mastrandrea, M.D.; Falcon, W.P.; Naylor, R.L. Prioritizing climate change adaptation needs for food security in 2030. Science 2008, 319, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, T.; Von Braun, J. Climate change impacts on global food security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Fujimori, S.; Hasegawa, T.; Krey, V.; Riahi, K.; Bertram, C.; Bodirsky, B.L.; Bosetti, V.; Callen, J.; Després, J.; Doelman, J.; et al. A multi-model assessment of food security implications of climate change mitigation. Nat. Sustain. 2019, 2, 386–396. [Google Scholar] [CrossRef]
- Calanca, P.P. Effects of abiotic stress in crop production. In Quantification of Climate Variability, Adaptation and Mitigation for Agricultural Sustainability; Springer: Cham, Germany, 2017; pp. 165–180. [Google Scholar]
- Gregory, P.J.; Johnson, S.N.; Newton, A.C.; Ingram, J.S. Integrating pests and pathogens into the climate change/food security debate. J. Exp. Bot. 2009, 60, 2827–2838. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.C. Climate change effects on insects: Implications for crop protection and food security. J. Crop Improv. 2014, 28, 229–259. [Google Scholar] [CrossRef]
- Garrett, K.A.; Nita, M.; De Wolf, E.D.; Esker, P.D.; Gomez-Montano, L.; Sparks, A.H. Plant pathogens as indicators of climate change. In Climate Change; Elsevier: Amsterdam, The Netherlands, 2016; pp. 325–338. [Google Scholar]
- Ziska, L.H.; Dukes, J.S. Weed Biology and Climate Change; Wiley-Blackwell: Hoboken, NJ, USA, 2011; p. 293. [Google Scholar]
- Boydston, R.A.; Mojtahedi, H.; Crosslin, J.M.; Brown, C.R.; Anderson, T. Effect of hairy nightshade (Solanum sarrachoides) presence on potato nematodes, diseases, and insect pests. Weed Sci. 2008, 56, 151–154. [Google Scholar] [CrossRef]
- Tofangsazi, N.; Hogg, B.N.; Portman, S.L.; Pratt, P.D. Tritrophic Interactions between an Invasive Weed (Lepidium latifolium), an Insect Herbivore (Bagrada hilaris), and a Plant Pathogenic Fungus (Albugo lepidii). Environ. Entom. 2019, 48, 1317–1322. [Google Scholar] [CrossRef] [PubMed]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–44. [Google Scholar] [CrossRef]
- Bunce, J.A.; Ziska, L.H. Crop ecosystem responses to climatic change: Crop/weed interactions. In Climate Change and Global Crop Productivity; Wiley and Sons: Hoboken, NJ, USA, 2000; pp. 333–348. [Google Scholar]
- Peters, K.; Breitsameter, L.; Gerowitt, B. Impact of climate change on weeds in agriculture: A review. Agron. Sustain. Dev. 2014, 34, 707–721. [Google Scholar] [CrossRef]
- Varanasi, A.; Prasad, P.V.; Jugulam, M. Impact of climate change factors on weeds and herbicide efficacy. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2016; pp. 107–146. [Google Scholar]
- Ruttledge, A.; Chauhan, B.S. Climate Change and Weeds of Cropping Systems. In Crop Protection under Changing Climate; Springer: Cham, Germany, 2020; pp. 57–84. [Google Scholar]
- Ziska, L.H. The role of climate change and increasing atmospheric carbon dioxide on weed management: Herbicide efficacy. Agric. Ecosyst. Environ. 2016, 231, 304–309. [Google Scholar] [CrossRef]
- Waryszak, P.; Lenz, T.I.; Leishman, M.R.; Downey, P.O. Herbicide effectiveness in controlling invasive plants under elevated CO2: Sufficient evidence to rethink weeds management. J. Environ. Manag. 2018, 226, 400–407. [Google Scholar] [CrossRef]
- Refatti, J.P.; de Avila, L.A.; Camargo, E.R.; Ziska, L.H.; Oliveira, C.; Salas-Perez, R.; Rouse, C.E.; Roma-Burgos, N. High [CO2] and temperature increase resistance to cyhalofop-butyl in multiple-resistant Echinochloa colona. Front. Plant Sci. 2019, 10, 529. [Google Scholar] [CrossRef]
- Gianessi, L.P. The increasing importance of herbicides in worldwide crop production. Pest Manag. Sci. 2016, 69, 1099–1105. [Google Scholar] [CrossRef]
- Duke, S.O. The history and current status of glyphosate. Pest Manag. Sci. 2018, 74, 1027–1034. [Google Scholar] [CrossRef]
- Nandula, V.K. Herbicide resistance: Definitions and concepts. Glyphosate Resistance in Crops and Weeds: History, Development, and Management; Wiley and Sons: Hoboken, NJ, USA, 2010; pp. 35–43. [Google Scholar]
- Baucom, R.S.; Mauricio, R. Fitness costs and benefits of novel herbicide tolerance in a noxious weed. Proc. Natl. Acad. Sci. USA 2004, 101, 13386–13390. [Google Scholar] [CrossRef]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef]
- United States Geological Survey. Available online: https://water.usgs.gov/nawqa/pnsp/usage/maps/show_map.php?year=2016&map=GLYPHOSATE&hilo=L (accessed on 13 November 2020).
- Powles, S.B.; Lorraine-Colwill, D.F.; Dellow, J.J.; Preston, C. Evolved resistance to glyphosate in rigid ryegrass (Lolium rigidum) in Australia. Weed Sci. 1998, 46, 604–607. [Google Scholar] [CrossRef]
- VanGessel, M.J. Glyphosate-resistant horseweed from Delaware. Weed Sci. 2001, 49, 703–705. [Google Scholar] [CrossRef]
- WSSA (Weed Science Society of America). Available online: http://www.weedscience.org/Home.aspx (accessed on 20 November 2020).
- Strom, S.A.; Gonzini, L.C.; Mitsdarfer, C.; Davis, A.S.; Riechers, D.E.; Hager, A.G. Characterization of multiple herbicide–resistant waterhemp (Amaranthus tuberculatus) populations from Illinois to VLCFA-inhibiting herbicides. Weed Sci. 2019, 67, 369–379. [Google Scholar] [CrossRef]
- Investigate Midwest. Available online: https://investigatemidwest.org/2020/06/16/weve-got-it-everywhere-dicamba-damaging-trees-across-midwest-and-south/ (accessed on 14 November 2020).
- Egan, J.F.; Barlow, K.M.; Mortensen, D.A. A meta-analysis on the effects of 2, 4-D and dicamba drift on soybean and cotton. Weed Sci. 2014, 62, 193–206. [Google Scholar] [CrossRef]
- Hopes, T. Regulatory News. In Rothamsted Repository Download; Rothamsted Publishing: Rothamsted, UK, 2019. [Google Scholar]
- Perry, J.; Waglechner, N.; Wright, G. The prehistory of antibiotic resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025197. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. 2017. Available online: www.weedscience.org (accessed on 12 November 2020).
- Von Bubnoff, A. Seeking new antibiotics in nature’s backyard. Cell 2006, 127, 867–869. [Google Scholar] [CrossRef]
- Leftwich, P.T.; Bolton, M.; Chapman, T. Evolutionary biology and genetic techniques for insect control. Evol. Appl. 2016, 9, 212–230. [Google Scholar] [CrossRef]
- Westwood, J.H.; Charudattan, R.; Duke, S.O.; Fennimore, S.A.; Marrone, P.; Slaughter, D.C.; Swanton, C.; Zollinger, R. Weed management in 2050: Perspectives on the future of weed science. Weed Sci. 2018, 66, 275–285. [Google Scholar] [CrossRef]
- Thompson, J.N. Rapid evolution as an ecological process. Trends Ecol. Evol. 1998, 13, 329–332. [Google Scholar] [CrossRef]
- Neve, P.; Vila-Aiub, M.; Roux, F. Evolutionary-thinking in agricultural weed management. New Phytol. 2009, 184, 783–793. [Google Scholar] [CrossRef]
- Matzrafi, M.; Brunharo, C.; Tehranchian, P.; Hanson, B.D.; Jasieniuk, M. Increased temperatures and elevated CO2 levels reduce the sensitivity of Conyza canadensis and Chenopodium album to glyphosate. Sci. Rep. 2019, 9, 2228. [Google Scholar] [CrossRef] [PubMed]
- Ziska, L.H.; Blumenthal, D.M.; Franks, S.J. Understanding the nexus of rising CO2, climate change, and evolution in weed biology. Invasive Plant Sci. Manag. 2019, 12, 79–88. [Google Scholar] [CrossRef]
- Matzrafi, M. Climate change exacerbates pest damage through reduced pesticide efficacy. Pest Manag. Sci. 2019, 75, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Ziska, L.H.; Goins, E.W. Elevated atmospheric carbon dioxide and weed populations in glyphosate treated soybean. Crop Sci. 2006, 46, 1354–1359. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Wang, H.; Chauhan, B.S.; Adkins, S.W. Effect of elevated carbon dioxide concentration on growth, productivity and glyphosate response of parthenium weed (Parthenium hysterophorus L.). Pest Manag. Sci. 2019, 75, 2934–2941. [Google Scholar] [CrossRef]
- Ziska, L.H.; Gealy, D.R.; Tomecek, M.B.; Jackson, A.K.; Black, H.L. Recent and projected increases in atmospheric CO2 concentration can enhance gene flow between wild and genetically altered rice (Oryza sativa). PLoS ONE 2012, 7, e37522. [Google Scholar] [CrossRef]
- Raja, R.; Nguyen, T.T.; Slaughter, D.C.; Fennimore, S.A. Real-time robotic weed knife control system for tomato and lettuce based on geometric appearance of plant labels. Biosyst. Eng. 2020, 194, 152–164. [Google Scholar] [CrossRef]
- Mirsky, S.B.; Curran, W.S.; Mortenseny, D.M.; Ryany, M.R.; Shumway, D.L. Timing of cover-crop management effects on weed suppression in no-till planted soybean using a roller-crimper. Weed Sci. 2011, 59, 380–389. [Google Scholar] [CrossRef]
- Tang, X.; Lowder, L.G.; Zhang, T.; Malzahn, A.A.; Zheng, X.; Voytas, D.F.; Zhong, Z.; Chen, Y.; Ren, Q.; Li, Q.; et al. A CRISPR–Cpf1 system for efficient genome editing and transcriptional repression in plants. Nat. Plants 2017, 3, 17018. [Google Scholar] [CrossRef]
- Colas, F.; Cordeau, S.; Granger, S.; Jeuffroy, M.H.; Pointurier, O.; Queyrel, W.; Rodriguez, A.; Villerd, J.; Colbach, N. Co-development of a decision support system for integrated weed management: Contribution from future users. Eur. J. Agron. 2020, 114, 126010. [Google Scholar] [CrossRef]
- Korres, N.E.; Burgos, N.R.; Travlos, I.; Vurro, M.; Gitsopoulos, T.K.; Varanasi, V.K.; Duke, S.O.; Kudsk, P.; Brabham, C.; Rouse, C.E.; et al. New directions for integrated weed management: Modern technologies, tools and knowledge discovery. Adv. Agron. 2019, 155, 243–319. [Google Scholar]
- Shaner, D.L.; Beckie, H.J. The future for weed control and technology. Pest Manag. Sci. 2014, 70, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Matzrafi, M.; Seiwert, B.; Reemtsma, T.; Rubin, B.; Peleg, Z. Climate change increases the risk of herbicide-resistant weeds due to enhanced detoxification. Planta 2016, 244, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, K.; Matloob, A.; Aslam, F.; Florentine, S.K.; Chauhan, B.S. Weeds in a changing climate: Vulnerabilities, consequences, and implications for future weed management. Front. Plant Sci. 2017, 8, 95. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E. Current status and future prospects in herbicide discovery. Plants 2019, 8, 341. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziska, L.H. Climate Change and the Herbicide Paradigm: Visiting the Future. Agronomy 2020, 10, 1953. https://doi.org/10.3390/agronomy10121953
Ziska LH. Climate Change and the Herbicide Paradigm: Visiting the Future. Agronomy. 2020; 10(12):1953. https://doi.org/10.3390/agronomy10121953
Chicago/Turabian StyleZiska, Lewis H. 2020. "Climate Change and the Herbicide Paradigm: Visiting the Future" Agronomy 10, no. 12: 1953. https://doi.org/10.3390/agronomy10121953
APA StyleZiska, L. H. (2020). Climate Change and the Herbicide Paradigm: Visiting the Future. Agronomy, 10(12), 1953. https://doi.org/10.3390/agronomy10121953