Linking Cover Crop Residue Quality and Tillage System to CO2-C Emission, Soil C and N Stocks and Crop Yield Based on a Long-Term Experiment

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Evaluations of CO2-C in Different Tillage and Cover Crop Combinations
2.2.1. Treatments and Practices
2.2.2. CO2-C Emission Measurements
2.2.3. Soil Temperature and Moisture Measurements and Supporting Weather Conditions
2.3. Cover Crops Biomass Input to Soil, Residue Decomposition in Litter Bags and Half-Life Time
2.4. Apparent C Balance Calculation
2.5. Statistical Analysis
3. Results and Discussion
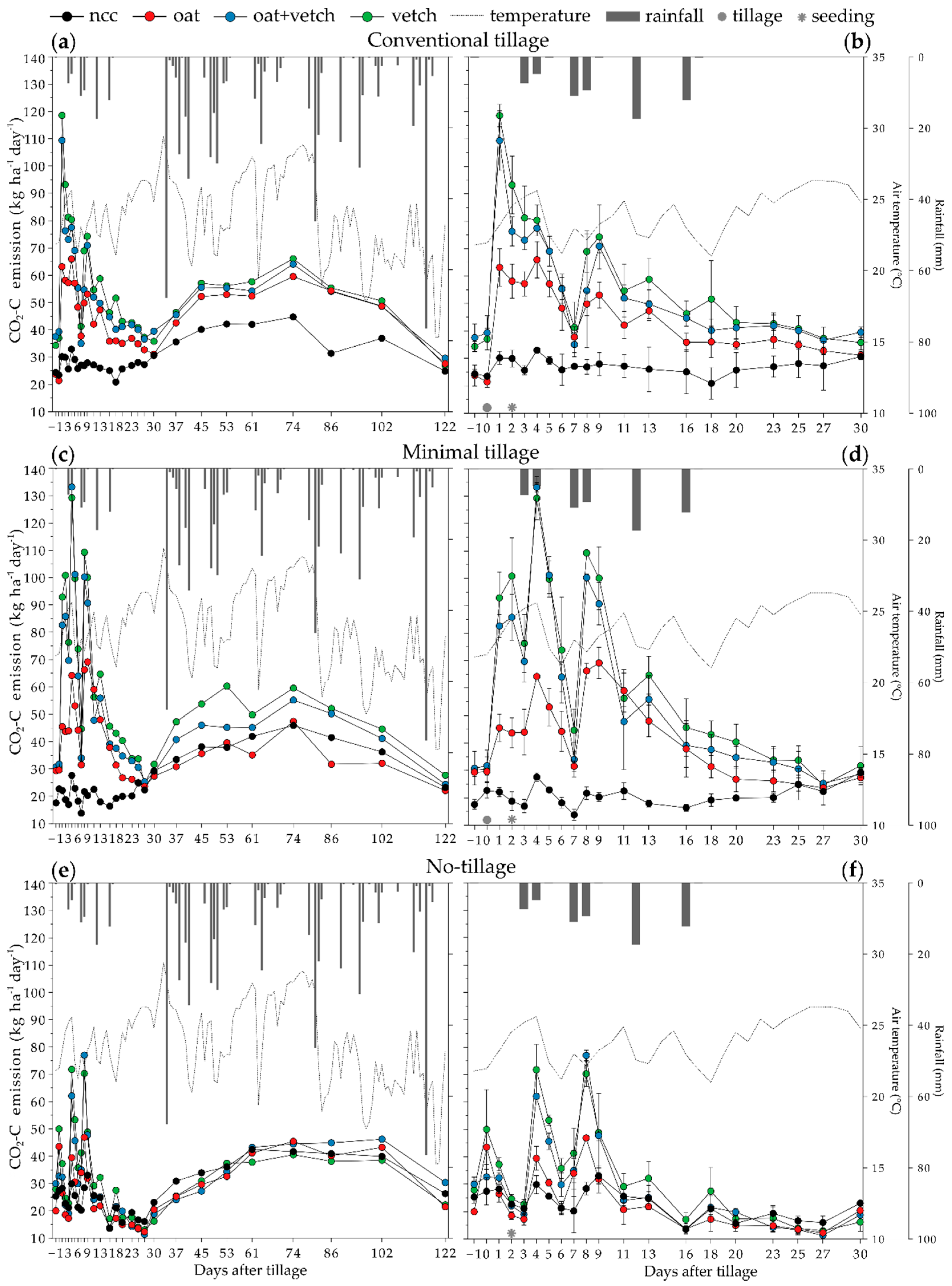
3.1. Weather Conditions, and Soil Temperature and Moisture Relation with Temporal CO2-C emission
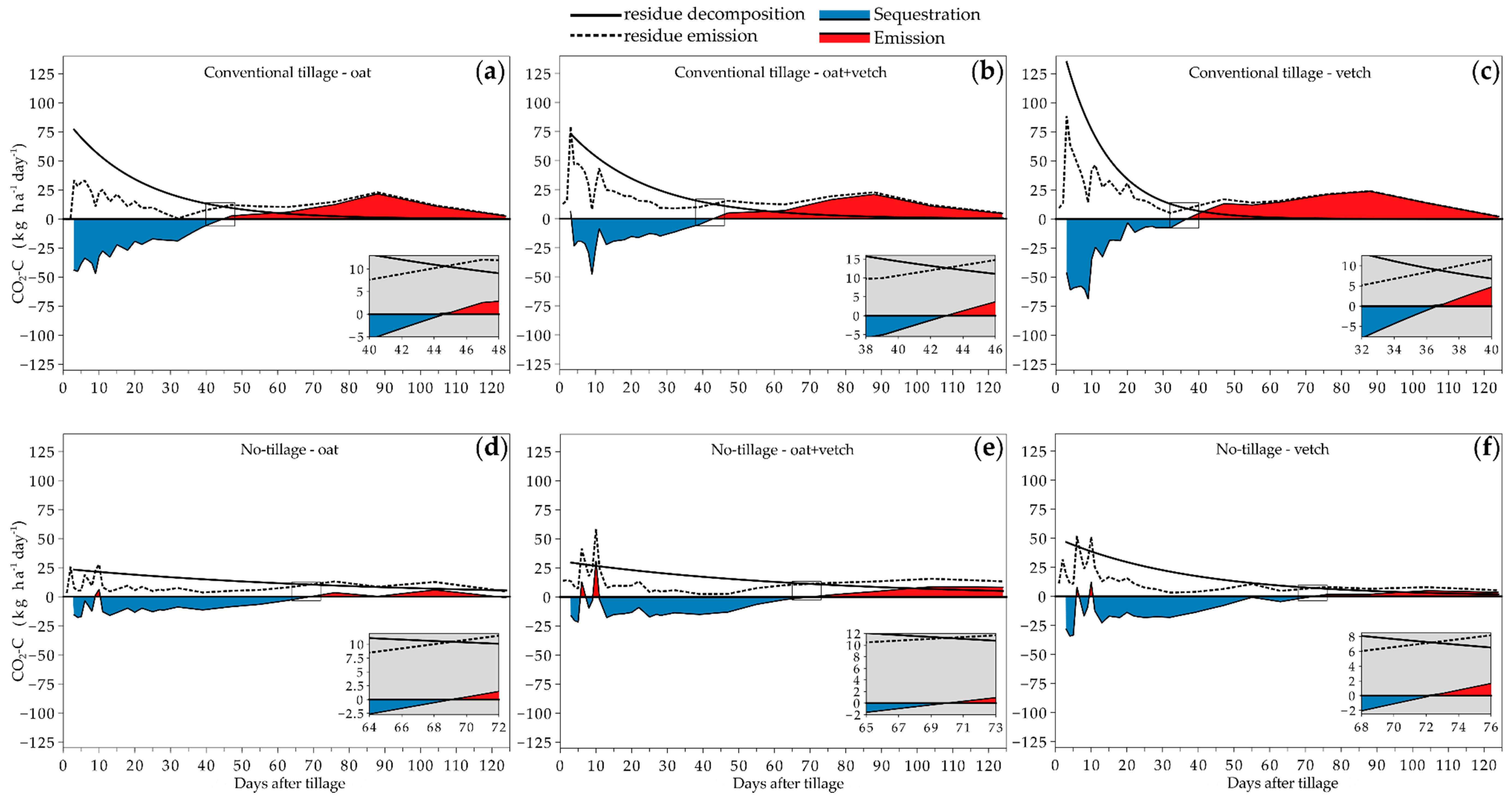
3.2. Temporal CO2-C Emission in Different Tillage and Cover Crop Combinations
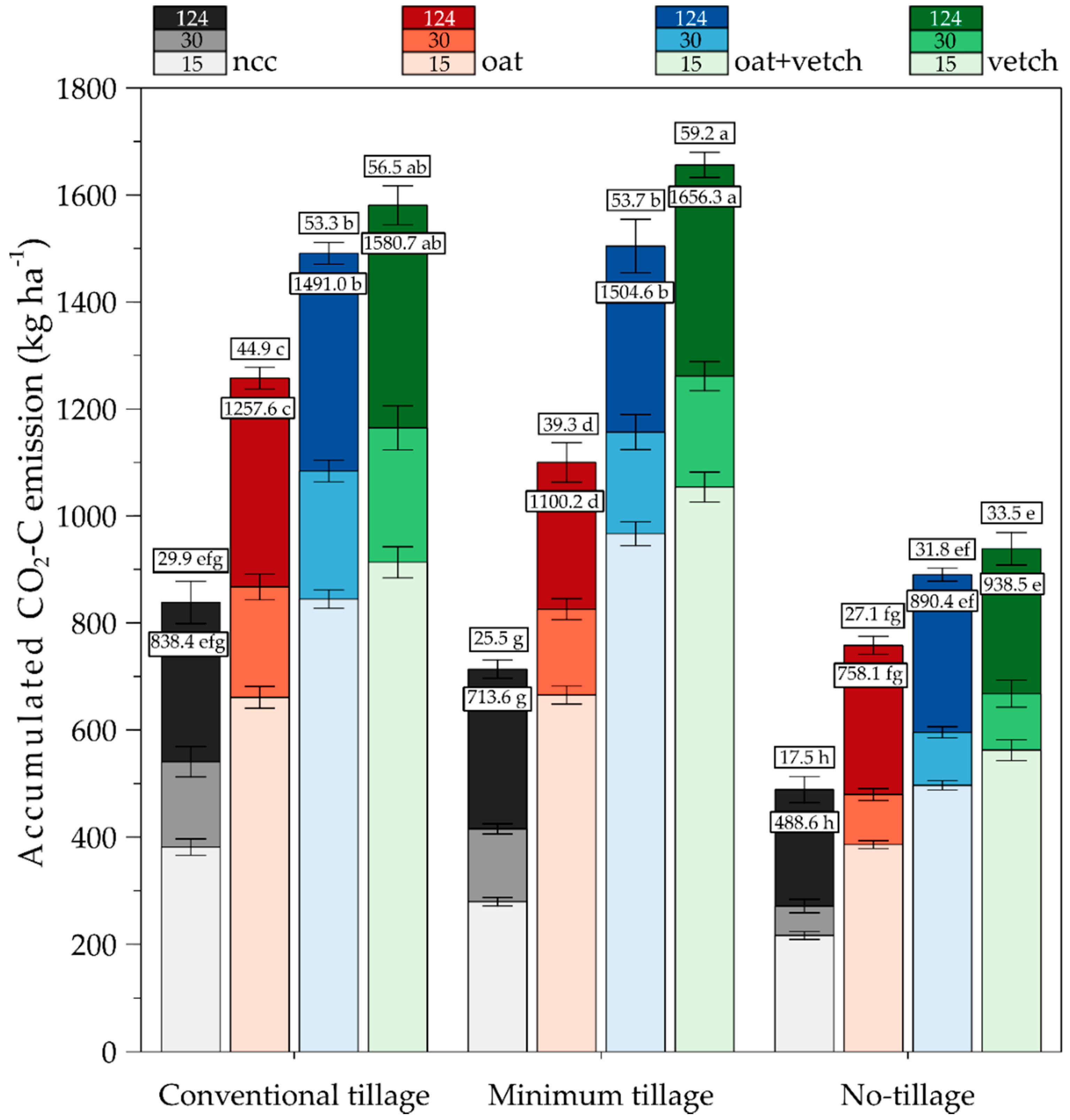
3.3. Daily Average and Accumulated CO2-C Emission in Different Tillage and Cover Crop Combinations
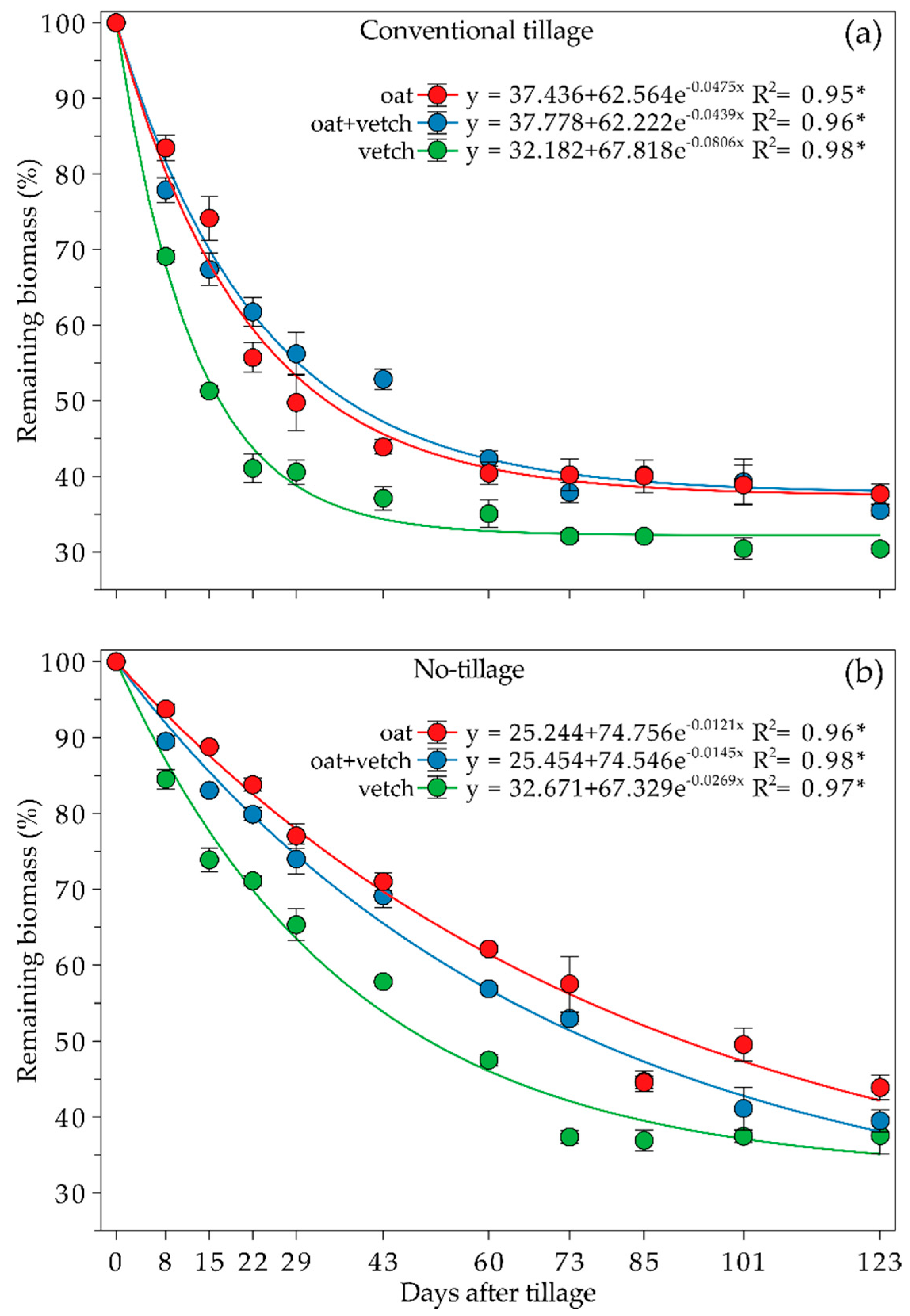
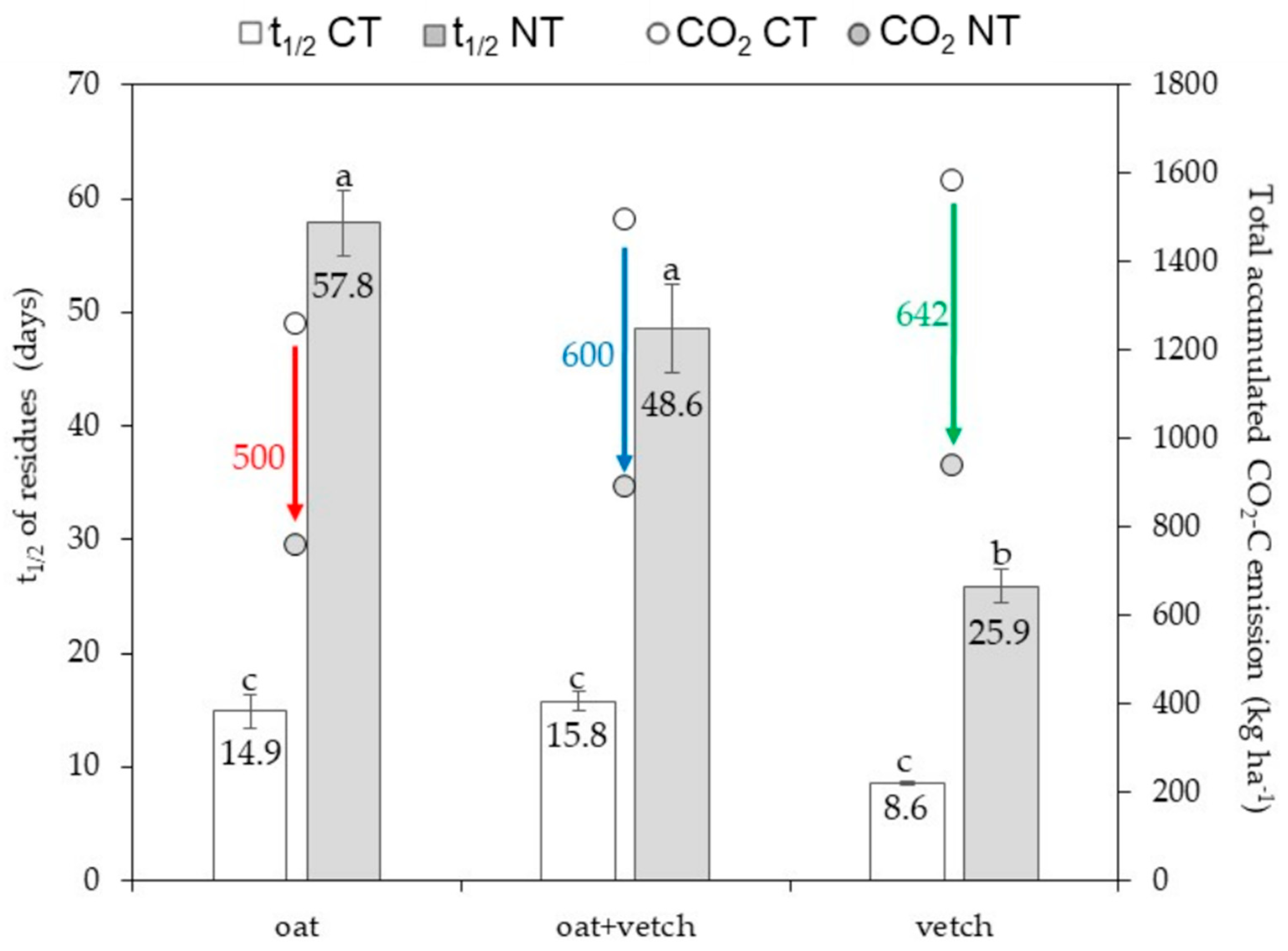
3.4. Cover Crops Biomass Input to Soil, Residue Decomposition in Litter Bags and Half-Life Time
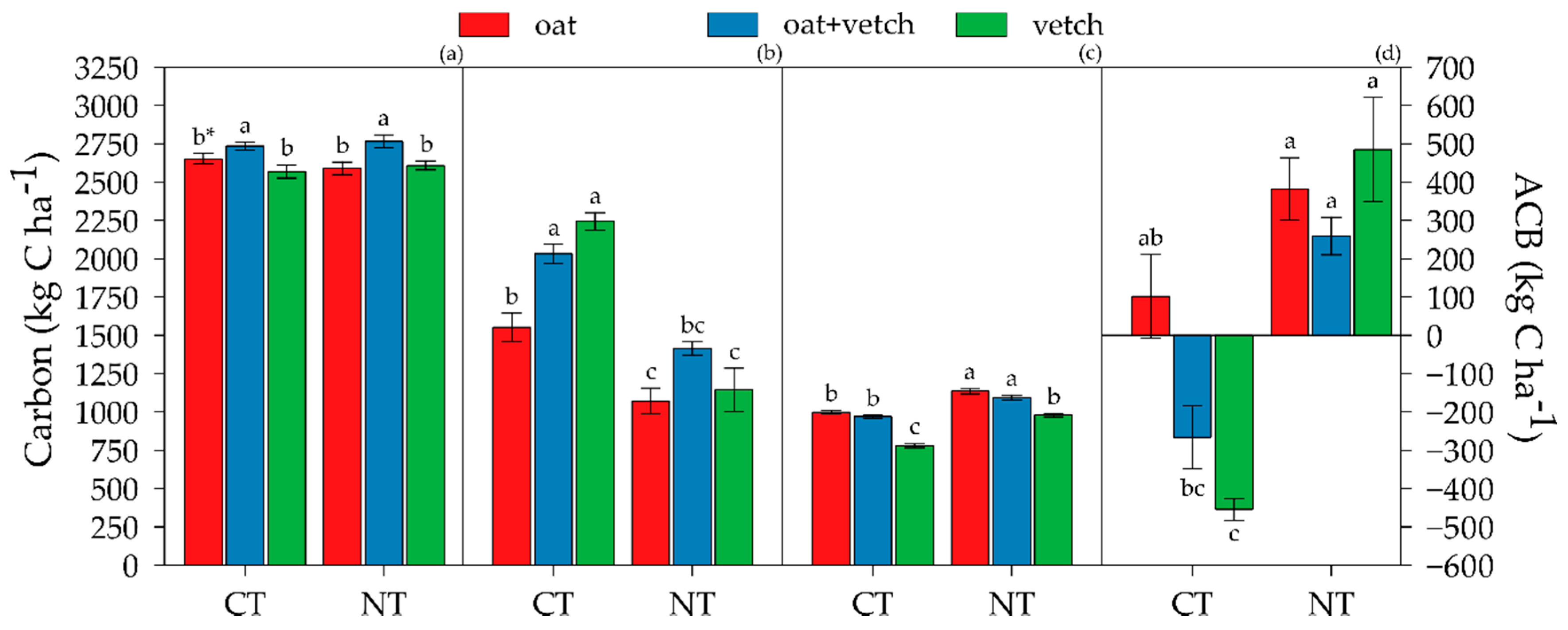
3.5. Apparent C Balance
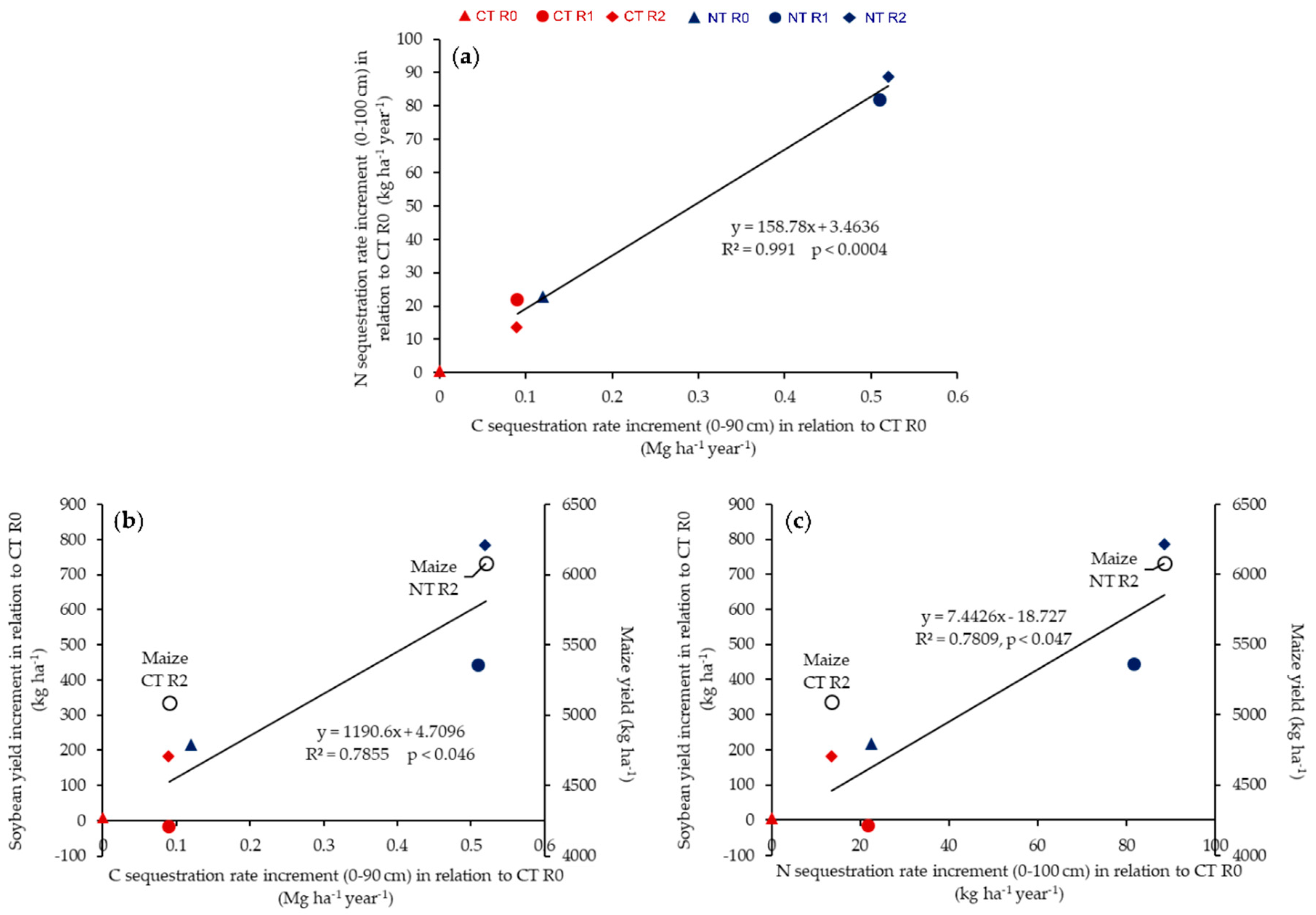
3.6. Linking CO2-C Emission and ACB to Long-Term Soil C and N Sequestration and Crop Yield
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wingeyer, A.B.; Amado, T.J.C.; Pérez-Bidegain, M.; Studdert, G.A.; Varela, C.H.P.; Garcia, F.O.; Karlen, D.L. Soil quality impacts of current South American agricultural practices. Sustainability 2015, 7, 2213–2242. [Google Scholar] [CrossRef]
- FAO. The State of the World’s Land and Water Resources for Food and Agriculture (SOLAW)—Managing Systems at Risks. Food and Agriculture Organization of the United Nations. 2011. Available online: http://www.fao.org/3/a-i1688e.pdf (accessed on 21 June 2018).
- De Figueiredo, E.B.; Panosso, A.R.; Reicosky, D.C.; La Scala, N. Short-term CO2-C emissions from soil prior to sugarcane (Saccharum spp.) replanting in southern Brazil. GCB Bioenergy 2015, 7, 316–327. [Google Scholar] [CrossRef]
- FAO; ITPS. Status of the World’s Soil Resources (SWSR)—Main Report. Food and Agriculture Organization of the United Nations and Intergovernmental Technical Panel on Soils. 2015. Available online: http://www.fao.org/3/i5199e/i5199e.pdf (accessed on 22 June 2018).
- Sanderman, J.; Hengl, T.; Fiske, G.J. Soil carbon debt of 12,000 years of human land use. Proc. Natl. Acad. Sci. USA 2017, 114, 9575–9580. [Google Scholar] [CrossRef] [PubMed]
- Paustian, K.; Lehmann, J.; Ogle, S.; Reay, D.; Robertson, G.P.; Smith, P. Climate-smart soils. Nature 2016, 532, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Hungria, M.; Franchini, J.C.; Brandão-Junior, O.; Kaschuk, G.; Souza, R.A. Soil microbial activity and crop sustainability in a long-term experiment with three soil-tillage and two crop-rotation systems. Appl. Soil Ecol. 2009, 42, 288–296. [Google Scholar] [CrossRef]
- Kassam, A.; Friedrich, T.; Derpsch, R. Global spread of Conservation Agriculture. Int. J. Environ. Stud. 2018, 76, 29–51. [Google Scholar] [CrossRef]
- Pittelkow, C.M.; Linquist, B.A.; Lundy, M.E.; Liang, X.; van Groenigen, K.J.; Lee, J.; van Gestel, N.; Six, J.; Venterea, R.T.; van Kessel, C. When does no-till yield more? A global meta-analysis. Field Crop. Res. 2015, 183, 156–168. [Google Scholar] [CrossRef]
- Amado, T.J.C.; Crusciol, C.A.C.; Costa, C.H.M.; Leal, O.A.; Pott, L.P. Rehabilitating degraded and abandoned agricultural lands with Conservation Agriculture systems. In Advances in Conservation Agriculture. Benefits and Practices, 1st ed.; Kassam, A., Farooq, M., Eds.; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2020; Volume 2, pp. 419–464. [Google Scholar]
- Ferreira, A.O.; Amado, T.J.C.; Nicoloso, R.S.; Sá, J.C.M.; Fiorin, J.E.; Hansel, D.S.S.; Menefee, D. Soil carbon stratification affected by long-term tillage and cropping systems in southern Brazil. Soil Tillage Res. 2013, 133, 65–74. [Google Scholar] [CrossRef]
- Ferreira, A.O.; Amado, T.J.C.; Rice, C.W.; Diaz, D.A.R.; Briedis, C.; Inagaki, T.M.; Gonçalves, D.R.P. Driving factors of soil carbon accumulation in Oxisols in long-termno-till systems of South Brazil. Sci. Total Environ. 2018, 622–623, 735–742. [Google Scholar] [CrossRef]
- Sisti, C.P.J.; Dos Santos, H.P.; Kohhann, R.; Alves, B.J.R.; Urquiaga, S.; Boddey, R.M. Change in carbon and nitrogen stocks in soil under 13 years of conventional or zero tillage in southern Brazil. Soil Tillage Res. 2004, 76, 39–58. [Google Scholar] [CrossRef]
- Brazil Ministry of Environment. Intended Nationally Determined Contributions, iNDC BRASIL. 2015. Available online: http://www.mma.gov.br/images/arquivo/80108/BRAZIL%20iNDC%20english%20FINAL.pdf (accessed on 12 May 2020).
- Zech, W.; Senesi, N.; Guggenberger, G.; Kaiser, K.; Lehmann, J.; Miano, T.M.; Miltner, A.; Schroth, G. Factors controlling humification and mineralization of soil organic matter in the tropics. Geoderma 1997, 79, 117–161. [Google Scholar] [CrossRef]
- Schlesinger, W.; Andrews, J. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Johnson, J.M.F.; Barbour, N.W.; Weyers, S.L. Chemical composition of crop biomass impacts its decomposition. Soil Sci. Soc. Am. J. 2007, 71, 155–162. [Google Scholar] [CrossRef]
- West, T.O.; Post, W.M. Soil organic carbon sequestration rates by tillage and crop rotation. Soil Sci. Soc. Am. J. 2002, 66, 1930–1946. [Google Scholar] [CrossRef]
- Lal, R. Farming carbon. Soil Tillage Res. 2007, 96, 1–5. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Cotrufo, M.F.; Ranalli, M.G.; Haddix, M.L.; Six, J.; Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 2019, 12, 989–994. [Google Scholar] [CrossRef]
- Veloso, M.G.; Angers, D.A.; Chantigny, M.H.; Bayer, C. Carbon accumulation and aggregation are mediated by fungi in a subtropical soil under conservation agriculture. Geoderma 2020, 363, 114159. [Google Scholar] [CrossRef]
- White, P.M.; Rice, C.W. Tillage effects on microbial and carbon dynamics during plant residue decomposition. Soil Sci. Soc. Am. J. 2009, 73, 138–145. [Google Scholar] [CrossRef]
- Wilson, G.W.T.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Kopttike, P.M.; Hernandez-Soriano, M.C.; Dalal, R.C.; Finn, D.; Menzies, N.W.; Hoeschen, C.; Mueller, C.W. Nitrogen-rich microbial products provide new organo-mineral associations for the stabilization of soil organic matter. Glob. Chang. Biol. 2018, 24, 1762–1770. [Google Scholar] [CrossRef] [PubMed]
- Arachchige, P.S.P.; Hettiarachchi, G.M.; Rice, C.W.; Dynes, J.J.; Maumann, L.; Wang, J.; Karunakaran, C.; Kilcoyne, A.L.D.; Attanayake, C.P.; Amado, T.J.C.; et al. Sub-micron level investigation reveals the inaccessibility of stabilized carbon in soil microaggregates. Sci. Rep. 2018, 8, 16810. [Google Scholar] [CrossRef] [PubMed]
- Veloso, M.G.; Angers, D.A.; Tiecher, T.; Giacomini, S.; Dieckow, J.; Bayer, C. High carbon storage in a previously degraded subtropical soil under no-tillage with legume cover crops. Agric. Ecosyst. Environ. 2018, 268, 15–23. [Google Scholar] [CrossRef]
- Santos, N.Z.D.; Dieckow, J.; Bayer, C.; Molin, R.; Favaretto, N.; Pauletti, V.; Piva, J.T. Forages, cover crops and related shoot and root additions in no-till rotations to C sequestration in a subtropical Ferralsol. Soil Tillage Res. 2011, 111, 208–218. [Google Scholar] [CrossRef]
- Boddey, R.M.; Jantalia, C.P.; Conceição, P.C.; Zanatta, J.A.; Bayer, C.; Mielniczuk, J.; Dieckow, J.; Dos Santos, H.P.; Denardin, J.; Aita, C.; et al. Carbon accumulation at depth in Ferralsols under zero-till subtropical agriculture. Glob. Chang. Biol. 2010, 16, 784–795. [Google Scholar] [CrossRef]
- Ebelhar, S.A.; Frye, W.W.; Blevins, R.L. Nitrogen from legume cover crops for no-tillage corn. Agron. J. 1984, 76, 51–55. [Google Scholar] [CrossRef]
- Holderbaum, J.F.; Decker, A.M.; Meisinger, J.J.; Mulford, F.R.; Vough, L.R. Fall-seeded legume cover crops for no-tillage corn in the humid east. Agron. J. 1990, 82, 117–124. [Google Scholar] [CrossRef]
- Seo, J.; Meisinger, J.J.; Lee, H. Recovery of nitrogen-15–labeled hairy vetch and fertilizer applied to corn. Agron. J. 2006, 98, 245–254. [Google Scholar] [CrossRef]
- Bayer, C.; Dieckow, J.; Amado, T.J.C.; Eltz, F.L.F.; Vieira, F.C.B. Cover crop effects increasing carbon storage in a subtropical no-till sandy Acrisol. Comm. Soil Sci. Plant Anal. 2009, 40, 1499–1511. [Google Scholar] [CrossRef]
- Conceição, P.C.; Dieckow, J.; Bayer, C. Combined role of no-tillage and cropping systems in soil carbon stocks and stabilization. Soil Tillage Res. 2013, 129, 40–47. [Google Scholar] [CrossRef]
- Chenu, C.; Angers, D.A.; Barré, P.; Derrien, D.; Arrouays, D.; Balesdent, J. Increasing organic stocks in agricultural soils: Knowledge gaps and potential innovations. Soil Tillage Res. 2018, 188, 41–52. [Google Scholar] [CrossRef]
- Campos, B.C.; Reinert, D.J.; Nicolidi, R.; Ruedell, J.; Petrere, C. Soil structural stability of a Dar-Red Latosol after seven years under crop rotation and management systems. Rev. Bras. Ciênc. Solo 1995, 19, 121–126. [Google Scholar]
- Nicoloso, R.S. Soil Organic Carbon Stocks and Stabilization Mechanisms on Temperate and Sub-Tropical Climate Agroecosystems. Ph.D. Thesis, Universidade Federal de Santa Maria, Rio Grande do Sul, Brazil, 21 July 2009. [Google Scholar]
- Campos, B.C.; Carneiro, T.J.C.; Bayer, C.; Nicoloso, R.D.S.; Fiorin, J.E. Carbon stock and its compartments in a subtropical Oxisol under long-term tillage and crop rotation systems. Rev. Bras. Cienc. Solo 2011, 35, 805–817. [Google Scholar] [CrossRef]
- Campos, B.C.; Amado, T.J.C.; Tornquist, C.G.; Nicoloso, R.S.; Fiorin, J.E. Long-term C-CO2 emissions and carbon crop residue mineralization in an Oxisol under different tillage and crop rotation systems. Rev. Bras. Ciênc. Solo 2011, 35, 819–832. [Google Scholar] [CrossRef]
- Nicoloso, R.S.; Amado, T.J.C.; Rice, C.W.; Pires, C.A.B.; Fiorin, J.E. A rotação de culturas aumenta os estoques de carbono e nitrogênio no solo sob sistema plantio direto. In Resultados Comparativos de 32 Anos dos Sistemas Plantio Direto e Convencional, 1st ed.; Fiorin, J.E., Ruedell, J., Fernandes, A.M.F., Eds.; Sescoop/RS: Rio Grande do Sul, Brazil, 2019; Volume 1, pp. 143–155. ISBN 978-85-63500-43-4. [Google Scholar]
- Nicoloso, R.S.; Amado, T.J.C.; Rice, C.W. Assessing strategies to enhance soil carbon sequestration with the DSSAT-CENTURY model. Eur. J. Soil Sci. 2020, 71, 1034–1049. [Google Scholar] [CrossRef]
- Fabrizzi, K.P.; Rice, C.W.; Amado, T.J.C.; Fiorin, J.E.; Barbagelata, P.; Melchiori, M. Protection of soil organic C and N in temperate and tropical soils: Effect of native and agroecosystems. Biogeochemistry 2009, 92, 129–143. [Google Scholar] [CrossRef]
- Pires, C.A.B.; Amado, T.J.C.; Reimche, G.; Schwalbert, R.; Sarto, M.V.M.; Nicoloso, R.S.; Fiorin, J.E.; Rice, C.W. Diversified crop rotation with no-till changes microbial distribution with depth and enhances activity in a subtropical Oxisol. Eur. J. Soil Sci. 2020, 71, 1173–1187. [Google Scholar] [CrossRef]
- Fiorin, J.E.; Ruedell, J.; Fernandes, A.M.F. Efeito dos sistemas de manejo de solo e da rotação de culturas sobre o rendimento de grãos de milho, soja e trigo. In Resultados Comparativos de 32 Anos dos Sistemas Plantio Direto e Convencional, 1st ed.; Fiorin, J.E., Ruedell, J., Fernandes, A.M.F., Eds.; Sescoop/RS: Rio Grande do Sul, Brazil, 2019; Volume 1, pp. 65–82. ISBN 978-85-63500-43-4. [Google Scholar]
- Pes, L.Z.; Amado, T.J.C.; La Scala, N.; Bayer, C.; Fiorin, J.E. The primary sources of carbon loss during the crop-establishment period in a subtropical Oxisol under contrasting tillage systems. Soil Tillage Res. 2011, 117, 163–171. [Google Scholar] [CrossRef]
- Ferreira, A.O.; Amado, T.J.C.; Rice, C.W.; Diaz, D.A.R.; Keller, C.; Inagaki, T.M. Can no-till grain production restore soil organic carbon to levels natural grass in a subtropical Oxisol? Agric. Ecosyst. Environ. 2016, 229, 13–20. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; United States Department of Agriculture Natural Resources Conservation Service: Washington, DC, USA, 2014; pp. 257–271.
- Inda, A.V., Jr.; Klamt, E.; Nascimento, P.C. Composição da fase sólida mineral do solo. In Fundamentos de Química Do Solo, 2nd ed.; Meurer, E.J., Ed.; Gênesis: Rio Grande do Sul, Brazil, 2004; pp. 35–71. [Google Scholar]
- Albuquerque, J.A.; Reinert, D.J.; Fiorin, J.E.; Ruedell, J.; Petrere, C.; Fontinelli, F. Crop rotation and soil management systems: Effects on soil structure form after seven years. Rev. Bras. Ciênc. Solo 1995, 19, 115–119. [Google Scholar]
- Ruedell, J.; Fiorin, J.E.; Wyzykowski, T.; Fernandes, A.M.F.; Campos, B.C. Descrição do experimento de longa duração: 32 anos. In Resultados Comparativos de 32 Anos Dos Sistemas Plantio Direto e Convencional, 1st ed.; Fiorin, J.E., Ruedell, J., Fernandes, A.M.F., Eds.; Sescoop/RS: Rio Grande do Sul, Brazil, 2019; Volume 1, pp. 51–64. ISBN 978-85-63500-43-4. [Google Scholar]
- La Scala, N.; Lopes, A.; Marques, J.; Pereira, G.T. Carbon dioxide emissions after application of tillage systems for a dark red latosol in southern Brazil. Soil Tillage Res. 2001, 62, 163–166. [Google Scholar] [CrossRef]
- Walkley, A.; Black, A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Yeomans, J.C.; Bremner, J.M. A rapid and precise method for routine determination of organic carbon in soil. Commun. Soil Sci. Plan. 1998, 19, 1467–1476. [Google Scholar] [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen-total. In Methods of Soil Analysis. Chemical and Microbiological Properties, 2nd ed.; Klute, A., Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; Volume 9, pp. 595–624. [Google Scholar]
- Wieder, R.K.; Lang, G.E. A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology 1982, 63, 1636–1642. [Google Scholar] [CrossRef]
- Plant, A.F.; Parton, W.J. The dynamics of soil organic matter and nutrient cycling. In Soil Microbiology, Ecology and Biochemistry, 3rd ed.; Paul, E.A., Ed.; Academic Press: San Diego, CA, USA, 2007; pp. 433–467. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models; R Package Version 3.1-145. 2020. Available online: https://cran.r-project.org/web/packages/nlme/index.html (accessed on 18 November 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 20 November 2019).
- Campos, B.C. Carbon Dynamics on a Rhodic Hapludox on Soil Tillage and Crop Systems. Ph.D. Thesis, Universidade Federal de Santa Maria, Rio Grande do Sul, Brazil, 27 April 2006. [Google Scholar]
- Bortolotto, R.P.; Amado, T.J.C.; Nora, D.D.; Keller, C.; Roberti, D.; Fiorin, J.E.; Reichardt, K.; Zamberlan, J.F.; Pasini, M.P.B.; Nicoloso, R.S. Soil carbon dioxide flux in a no-tillage winter system. Afri. J. Agr. Res. 2015, 10, 450–457. [Google Scholar] [CrossRef]
- O’Connell, A.M. Microbial decomposition (respiration) of litter in eucalypt forests of South-Western Australia: An empirical model based on laboratory incubations. Soil Biol. Biochem. 1990, 22, 153–160. [Google Scholar] [CrossRef]
- Paustian, K.; Andrén, O.; Janzen, H.H.; Lal, R.; Smith, P.; Tian, G.; Tiessen, H.; Noordwijk, M.; Woomer, P.L. Agricultural soils as a sink to mitigate CO2 emissions. Soil Use Manag. 1997, 13, 230–244. [Google Scholar] [CrossRef]
- Linn, D.M.; Doran, J.W. Effect of water-filled pore space on carbon dioxide and nitrous oxide production in tilled and nontilled soils. Soil Sci. Soc. Am. J. 1984, 48, 1267–1272. [Google Scholar] [CrossRef]
- Quincke, J.A.; Wortmann, C.S.; Mamo, M.; Franti, T.; Drijber, R.A. Occasional tillage of no-till systems: Carbon dioxide flux and changes in total and labile soil organic carbon. Agron. J. 2007, 99, 1158–1168. [Google Scholar] [CrossRef]
- Aita, C.; Chiapinotto, I.C.; Giacomini, S.J.; Hübner, A.P.; Marques, M.G. Decomposition of black oat straw and pig slurry in a soil under no-tillage. Rev. Bras. Ciênc. Solo 2006, 30, 149–161. (In Portuguese) [Google Scholar] [CrossRef]
- Hendry, M.J.; Wassenaar, L.I.; Birkham, T.K. Microbial respiration and diffusive transport of O2, 16O2, and 18O16O in unsaturated soils: A mesocosm experiment. Geochim. Cosmochim. Acta 2002, 66, 3367–3374. [Google Scholar] [CrossRef]
- Risk, D.; Kellman, L.; Beltrami, H. Carbon dioxide in soil profiles: Production and temperature dependence. Geophys. Res. Lett. 2002, 29, 11–14. [Google Scholar] [CrossRef]
- La Scala, N.; Lopes, A.; Spokas, K.; Archer, D.W.; Reicoscky, D.C. Short-term temporal changes of bare soil CO2 fluxes after tillage described by first-order decay models. Eur. J. Soil Sci. 2009, 60, 258–264. [Google Scholar] [CrossRef]
- Reicosky, D.C.; Lindstrom, M.J. Fall tillage method: Effect on short-term carbon dioxide flux from soil. Agron. J. 1993, 85, 1237–1243. [Google Scholar] [CrossRef]
- La Scala, N.; Lopes, A.; Panosso, A.R.; Camara, F.T.; Pereira, G.T. Soil CO2 efflux following rotary tillage of a tropical soil. Soil Tillage Res. 2005, 84, 222–225. [Google Scholar] [CrossRef]
- Teixeira, L.G.; Corradi, M.M.; Fukuda, A.; Panosso, A.R.; Reicoscky, D.; Lopes, A.; La Scala, N. Soil and crop residue CO2-C emission under tillage systems in sugarcane-producing areas of southern Brazil. Sci. Agric. 2013, 70, 327–335. [Google Scholar] [CrossRef]
- Rigon, J.P.G.; Calonego, J.C.; Rosolem, C.A.; La Scala, N. Cover crop rotations in no-till system: Short-term CO2 emissions and soybean yield. Sci. Agric. 2018, 75, 18–26. [Google Scholar] [CrossRef]
- Reicosky, D.C.; Archer, D.W. Moldboard plow tillage depth and short-term carbon dioxide release. Soil Tillage Res. 2007, 94, 109–121. [Google Scholar] [CrossRef]
- Reicosky, D.C.; Dugas, W.A.; Torbert, H.A. Tillage-induced soil carbon dioxide loss from different cropping systems. Soil Tillage Res. 1997, 41, 105–118. [Google Scholar] [CrossRef]
- Aita, C.; Giacomini, S.J. Crop residue decomposition and nitrogen release in single and mixed cover crops. Rev. Bras. Ciênc. Solo 2003, 27, 601–612. (In Portuguese) [Google Scholar] [CrossRef]
- Abdalla, M.; Hastings, A.; Helmy, M.; Prescher, A.; Osborne, B.; Lanigan, G.; Forristal, D.; Killi, D.; Maratha, P.; Williams, M.; et al. Assessing the combined use of reduced tillage and cover crops for mitigating greenhouse gas emissions from arable ecosystem. Geoderma 2014, 223–225, 9–20. [Google Scholar] [CrossRef]
- Rochette, P.; Angers, D.A. Soil surface carbon dioxide fluxes induced by spring, summer, and fall moldboard plowing in a sandy loam. Soil Sci. Soc. Am. J. 1999, 63, 621–628. [Google Scholar] [CrossRef]
- La Scala, N.; Lopes, A.; Spokas, K.; Bolonhezi, D.; Archer, D.W.; Reicosky, D.C. Short-term temporal changes of soil carbon losses after tillage described by a first-order decay model. Soil Tillage Res. 2008, 99, 108–118. [Google Scholar] [CrossRef]
- Alvarez, R.; Alvarez, C.R.; Lorenzo, G. Carbon dioxide fluxes following tillage from a mollisol in the Argentine Rolling Pampa. Eur. J. Soil Biol. 2001, 37, 161–166. [Google Scholar] [CrossRef]
- Prior, S.A.; Rogers, H.H.; Runion, G.B.; Torbert, H.A.; Reicosky, D.C. Carbon dioxide-enriched agroecosystems: Influence of tillage on short-term soil carbon dioxide efflux. J. Environ. Qual. 1997, 26, 244–252. [Google Scholar] [CrossRef]
- Prior, S.A.; Reicosky, D.C.; Reeves, D.W.; Runion, G.B.; Raper, R.L. Residue and tillage effects on planting implement-induced short-term CO2 and water loss from a loamy sand soil in Alabama. Soil Tillage Res. 2000, 54, 197–199. [Google Scholar] [CrossRef]
- Reicosky, D.C.; Lindstrom, M.J.; Schumacher, T.E.; Lobb, D.E.; Malo, D.D. Tillage-induced CO2 loss across an eroded landscape. Soil Tillage Res. 2005, 81, 183–194. [Google Scholar] [CrossRef]
- La Scala, N.; Bolonhezi, D.; Pereira, G.T. Short-term soil CO2 emission after conventional and reduced tillage of a no-till sugar cane area in southern Brazil. Soil Tillage Res. 2006, 91, 244–248. [Google Scholar] [CrossRef]
- Costa, F.S.; Bayer, C.; Zanatta, J.A.; Mielniczuk, J. Carbon stock and carbon dioxide emissions as affected by soil management systems in southern Brazil. Rev. Bras. Ciênc. Solo 2008, 32, 323–332. (In Portuguese) [Google Scholar] [CrossRef]
- Sainju, U.M.; Singh, B.P.; Whitehead, W.F. Long-term effects of tillage, cover crops, and nitrogen fertilization on organic carbon and nitrogen concentrations in sandy loam soils in Georgia, USA. Soil Tillage Res. 2002, 63, 167–179. [Google Scholar] [CrossRef]
- Al-Kaisi, M.M.; Yin, X. Tillage and crop residue effects on soil carbon and carbon dioxide emission in corn–soybean rotations. J. Environ. Qual. 2005, 34, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Li, L.J.; Han, X.Z.; You, M.Y.; Yuan, Y.R.; Ding, X.L.; Qiao, Y.F. Carbon and nitrogen mineralization patterns of two contrasting crop residues in a Mollisol: Effects of residue type and placement in soils. Eur. J. Soil Biol. 2013, 54, 1–6. [Google Scholar] [CrossRef]
- Sievers, T.; Cook, R.L. Aboveground and root decomposition of cereal rye and hairy vetch cover crops. Soil Sci. Soc. Am. J. 2018, 82, 147–155. [Google Scholar] [CrossRef]
- Acosta, J.A.A.; Amado, T.J.C.; Neergaard, A.; Vinther, M.; Silva, L.S.; Nicoloso, R.S. Effect of 15N-labeled hairy vetch and nitrogen fertilization on maize nutrition and yield under no-tillage. Rev. Bras. Ciênc. Solo 2011, 35, 1337–1345. [Google Scholar] [CrossRef]
- Jackson, L.E.; Calderon, F.J.; Steenwerth, K.L.; Scow, K.M.; Rolston, D.E. Responses of soil microbial processes and community structure to tillage events and implications for soil quality. Geoderma 2003, 114, 305–317. [Google Scholar] [CrossRef]
- Kramberger, B.; Gselman, A.; Janzekovic, M.; Kaligaric, M.; Bracko, B. Effects of cover crops on soil mineral nitrogen and on the yield and nitrogen content of maize. Eur. J. Agron. 2009, 31, 103–109. [Google Scholar] [CrossRef]
- Veloso, M.G.; Dick, D.P.; Costa, J.B.; Bayer, C. Cropping systems including legume cover crops favour mineral–organic associations enriched with microbial metabolites in no-till soil. Soil Res. 2019, 57, 851–858. [Google Scholar] [CrossRef]
- Poffenbarger, H.J.; Mirsky, S.B.; Weil, R.R.; Kramer, M.; Spargo, J.T.; Cavigelli, M.A. Legume proportion, poultry litter, and tillage effects on cover crop decomposition. Agron. J. 2015, 107, 2083–2096. [Google Scholar] [CrossRef]
- Teasdale, J.R.; Mohler, C.L. Light transmittance, soil temperature, and soil moisture under residue of hairy vetch and rye. Agron. J. 1993, 85, 673–680. [Google Scholar] [CrossRef]
- Manzoni, S.; Jackson, R.B.; Trofymow, J.A.; Porporato, A. The global stoichiometry of litter nitrogen mineralization. Science 2008, 321, 684–686. [Google Scholar] [CrossRef]
- Kallenbach, C.M.; Frey, S.D.; Grandy, A.S. Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 2016, 7, 13630. [Google Scholar] [CrossRef]
- Maron, P.A.; Sarr, A.; Kaisermann, A.; Lévêque, J.; Mathieu, O.; Guigue, J.; Karimi, B.; Bernard, L.; Dequiedt, S.; Terrat, S.; et al. High microbial diversity promotes soil ecosystem functioning. Appl. Environ. Microbiol. 2018, 84, 1–13. [Google Scholar] [CrossRef]
- Mikha, M.M.; Rice, C.W. Tillage and manure effects on soil and aggregate- associated carbon and nitrogen. Soil Sci. Soc. Am. J. 2004, 68, 809–816. [Google Scholar] [CrossRef]
- Dieckow, J.; Mielniczuk, J.; Knicer, H.; Bayer, H.; Dick, D.P.; Kögel-Knaber, I. Soil C and N stocks as affected by cropping systems and nitrogen fertilisation in a southern Brazil Acrisol managed under no-tillage for 17 years. Soil Tillage Res. 2005, 81, 87–95. [Google Scholar] [CrossRef]
- Totsche, K.U.; Amelung, W.; Gerbazek, M.; Guggenberger, G.; Klumpp, E.; Knief, C.; Lehndorff, E.; Mikutta, R.; Peth, S.; Pretchel, A.; et al. Microaggregates in soils. J. Plant Nutr. Soil Sci. 2017, 181, 104–136. [Google Scholar] [CrossRef]
- Ferreira, A.O.; Sá, J.C.M.; Lal, R.; Tivet, F.; Briedis, C.; Inagaki, T.M.; Gonçalves, D.R.P.; Romaniw, J. Macroaggregation and soil organic carbon restoration in a highly weathered Brazilian Oxisol after two decades under no-till. Sci. Total Environ. 2018, 621, 1559–1567. [Google Scholar] [CrossRef]
- Dieckow, J.; Bayer, C.; Conceição, P.C.; Zanatta, J.A.; Martin-Neto, L.; Milori, D.M.M.; Salton, J.C.; Macedo, M.M.; Mielniczuk, J.; Hernani, L.C. Land use, tillage, texture and organic matter stock and composition in tropical and subtropical Brazilian soils. Eur. J. Soil Sci. 2009, 60, 249. [Google Scholar] [CrossRef]
- Reis, C.E.S.; Dick, D.P.; Caldas, J.S.; Bayer, C. Carbon sequestration in clay and silt fractions of Brazilian soils under conventional and no-till systems. Sci. Agric. 2014, 71, 292–301. [Google Scholar] [CrossRef]
- Giacomini, S.J.; Recous, S.; Mary, B.; Aita, C. Simulating the effects of N availability, straw particle size and location in soil on C and N mineralization. Plant Soil 2007, 301, 289–301. [Google Scholar] [CrossRef]
- Nevins, C.J.; Nakatsu, C.; Armstrong, S. Characterization of microbial community response to cover crop residue decomposition. Soil Biol. Biochem. 2018, 127, 39–49. [Google Scholar] [CrossRef]
- Abdalla, K.; Chivenge, P.; Ciais, P.; Chaplot, V. No-tillage lessens soil CO2 emissions the most under arid and sandy soil conditions: Results from a meta-analysis. Biogeosciences 2016, 13, 3619–3633. [Google Scholar] [CrossRef]
- Schwen, A.; Jeitler, E.; Böttcher, J. Spatial and temporal variability of soil gas diffusivity, its scaling and relevance for soil respiration under different tillage. Geoderma 2015, 259–260, 323–336. [Google Scholar] [CrossRef]
- Schmatz, R.; Recous, S.; Aita, C.; Tahir, M.M.; Schu, A.L.; Chaves, B.; Giacomini, S.J. Crop residue quality and soil type influence the priming effect but not the fate of crop residue C. Plant Soil 2017, 414, 229–245. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Domanski, G. Carbon input by plants into the soil. Rev. J. Plant Nutr. Soil Sci. 2000, 163, 421–431. [Google Scholar] [CrossRef]
- Castellano, M.J.; Mueller, K.E.; Olk, D.C.; Sawyer, J.E.; Six, J. Integrating plant litter quality, soil organic matter stabilization, and the carbon saturation concept. Glob. Chang. Biol. 2015, 21, 3200–3209. [Google Scholar] [CrossRef]
- Gale, W.; Cambardella, C.A. Carbon dynamics of surface residue- and root-derived organic matter under simulated no-till. Soil Sci. Soc. Am. J. 2000, 64, 190–195. [Google Scholar] [CrossRef]
- Zanatta, J.A.; Bayer, C.; Dieckow, J.; Vieira, F.C.B.; Mielniczuk, J. Soil organic carbon accumulation and carbon costs related to tillage, cropping systems and nitrogen fertilization in a subtropical Acrisol. Soil Tillage Res. 2007, 94, 510–519. [Google Scholar] [CrossRef]
- Kleber, M.; Sollins, P.; Sutton, R. A conceptual model of organo-mineral interactions in soils: Self-assembly of organic molecular fragments into zonal structures on mineral surfaces. Biogeochemistry 2007, 85, 9–24. [Google Scholar] [CrossRef]
- Torabian, S.; Farhangi-Abriz, S.; Denton, M.D. Do tillage systems influence nitrogen fixation in legumes? A review. Soil Tillage Res. 2019, 185, 113–121. [Google Scholar] [CrossRef]
- Bayer, C.; Mielniczuk, J.; Amado, T.J.C.; Martin-Neto, L.; Fernandes, S.V. Organic matter storage in a sandy clay loam acrisol affected by tillage and cropping systems in southern Brazil. Soil Tillage Res. 2000, 54, 101–109. [Google Scholar] [CrossRef]
- Bayer, C.; Lovato, T.; Dieckow, J.; Zanatta, J.A.; Mielniczuk, J. A method for estimating coefficients of soil organic matter dynamics based on long-term experiments. Soil Tillage Res. 2006, 91, 217–226. [Google Scholar] [CrossRef]
- Amado, T.J.C.; Bayer, C.; Conceição, P.C.; Spagnollo, E.; Campos, B.H.C.; Veiga, M. Potential of carbon accumulation in no-till soils with intensive use and cover crops in southern Brazil. J. Environ. Qual. 2006, 35, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Sanchez, E.J.; Veroz-Gonzalez, O.; Conway, G.; Moreno-Garcia, M.; Kassam, A.; Mkomwa, S.; Ordoñez-Fernandez, R.; Triviño-Tarradas, P.; Carbonell-Bojollo, R. Meta-analysis on carbon sequestration through conservation agriculture in Africa. Soil Tillage Res. 2019, 190, 22–30. [Google Scholar] [CrossRef]
- Calzarano, F.; Stagnari, F.; D’Egidio, S.; Pagnani, G.; Galieni, A.; Di Marco, S.; Metruccio, E.G.; Pisante, M. Durum wheat quality, yield and sanitary status under conservation agriculture. Agriculture 2018, 8, 140. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tillage System | Cover Crop | Dry Biomass | Carbon | Nitrogen | C/N |
|---|---|---|---|---|---|
| -------------------------kg ha−1------------------------ | |||||
| CT 1 | oat | 6598 ± 75.6 | 2651 ± 30.4 | 78 ± 4.5 | 34.0 |
| oat + vetch | 6780 ± 67.4 | 2735 ± 33.0 | 116 ± 6.3 | 23.6 | |
| vetch | 6401 ± 60.5 | 2569 ± 21.6 | 190 ± 6.6 | 13.5 | |
| MT 1 | oat | 6398 ± 52.4 | 2571 ± 35.7 | 75 ± 5.8 | 34.3 |
| oat + vetch | 6903 ± 72.1 | 2784 ± 28.9 | 118 ± 7.7 | 23.6 | |
| vetch | 6525 ± 83.6 | 2619 ± 42.0 | 194 ± 9.4 | 13.5 | |
| NT 1 | oat | 6442 ± 69.7 | 2589 ± 26.9 | 76 ± 3.7 | 34.1 |
| oat + vetch | 6858 ± 81.1 | 2766 ± 19.4 | 117 ± 7.1 | 23.4 | |
| vetch | 6498 ± 87.8 | 2608 ± 72.0 | 193 ± 8.6 | 13.5 | |
| Mean (CT and NT) 2 | oat | 6521 ± 47.7 ab | 2620 ± 19,1 b | 77.0 ± 0.60 c | - |
| oat + vetch | 6819 ± 73.6 a | 2751 ± 29.7 a | 117.0 ± 1.27 b | - | |
| vetch | 6450 ± 11.4 b | 2589 ± 47.1 b | 192.0 ± 3.49 a | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leal, O.A.; Amado, T.J.C.; Fiorin, J.E.; Keller, C.; Reimche, G.B.; Rice, C.W.; Nicoloso, R.S.; Bortolotto, R.P.; Schwalbert, R. Linking Cover Crop Residue Quality and Tillage System to CO2-C Emission, Soil C and N Stocks and Crop Yield Based on a Long-Term Experiment. Agronomy 2020, 10, 1848. https://doi.org/10.3390/agronomy10121848
Leal OA, Amado TJC, Fiorin JE, Keller C, Reimche GB, Rice CW, Nicoloso RS, Bortolotto RP, Schwalbert R. Linking Cover Crop Residue Quality and Tillage System to CO2-C Emission, Soil C and N Stocks and Crop Yield Based on a Long-Term Experiment. Agronomy. 2020; 10(12):1848. https://doi.org/10.3390/agronomy10121848
Chicago/Turabian StyleLeal, Otávio A., Telmo J. C. Amado, Jackson E. Fiorin, Cristiano Keller, Geovane B. Reimche, Charles W. Rice, Rodrigo S. Nicoloso, Rafael P. Bortolotto, and Rai Schwalbert. 2020. "Linking Cover Crop Residue Quality and Tillage System to CO2-C Emission, Soil C and N Stocks and Crop Yield Based on a Long-Term Experiment" Agronomy 10, no. 12: 1848. https://doi.org/10.3390/agronomy10121848
APA StyleLeal, O. A., Amado, T. J. C., Fiorin, J. E., Keller, C., Reimche, G. B., Rice, C. W., Nicoloso, R. S., Bortolotto, R. P., & Schwalbert, R. (2020). Linking Cover Crop Residue Quality and Tillage System to CO2-C Emission, Soil C and N Stocks and Crop Yield Based on a Long-Term Experiment. Agronomy, 10(12), 1848. https://doi.org/10.3390/agronomy10121848