Effect of Nutrient Solution Concentration on the Growth of Hydroponic Sweetpotato


Abstract
1. Introduction
2. Materials and Methods
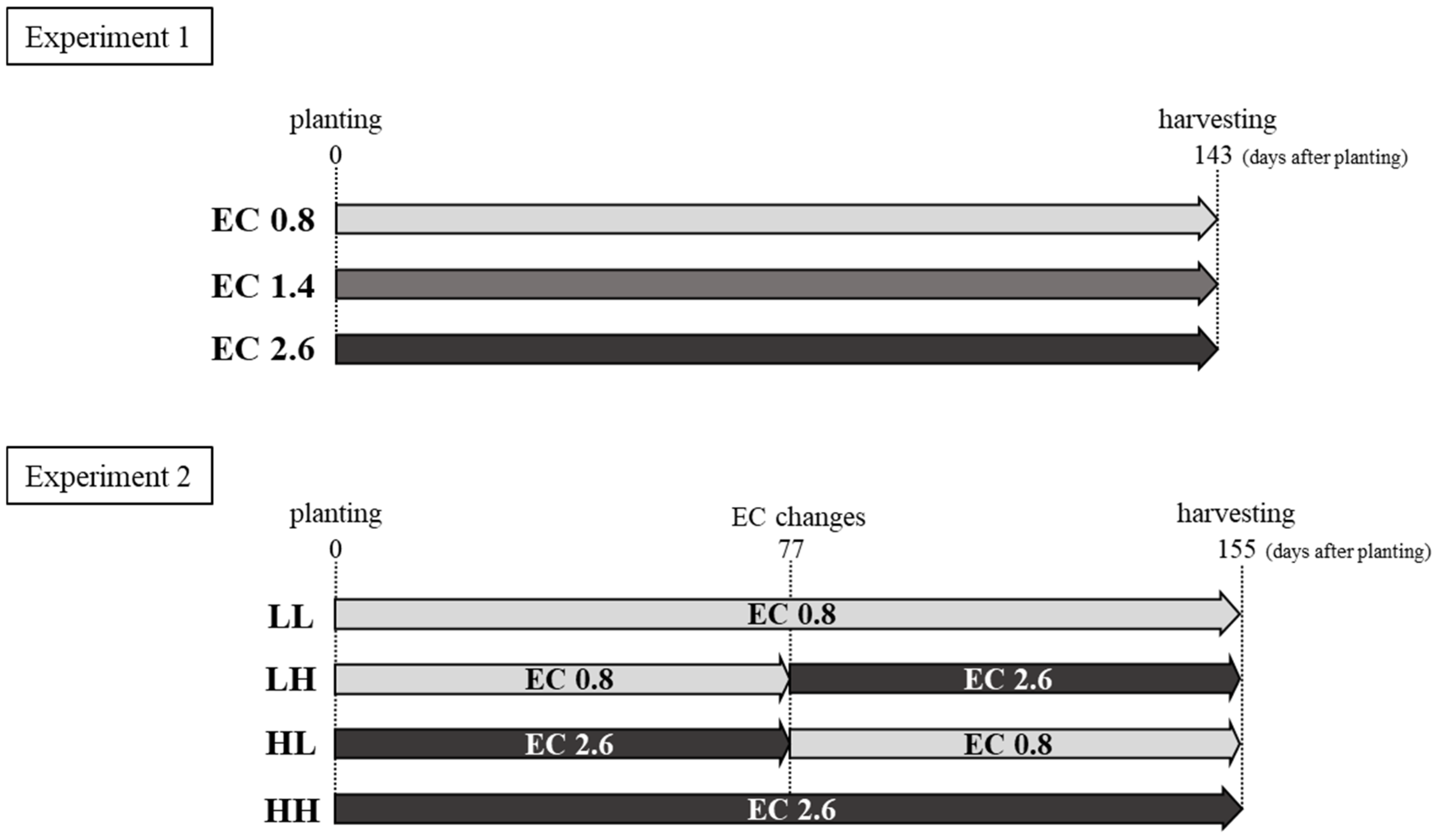
2.1. Experimental Conditions
2.2. Measurement of Plant Growth and Yield
2.3. Measurement of Chlorophyll Contents
2.4. Data Analysis
3. Results
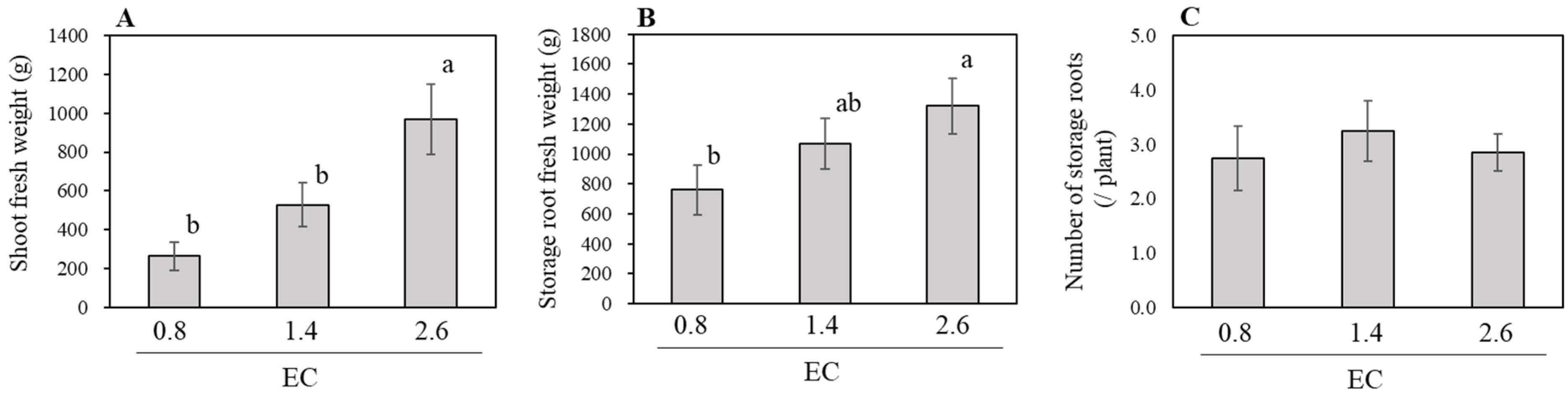
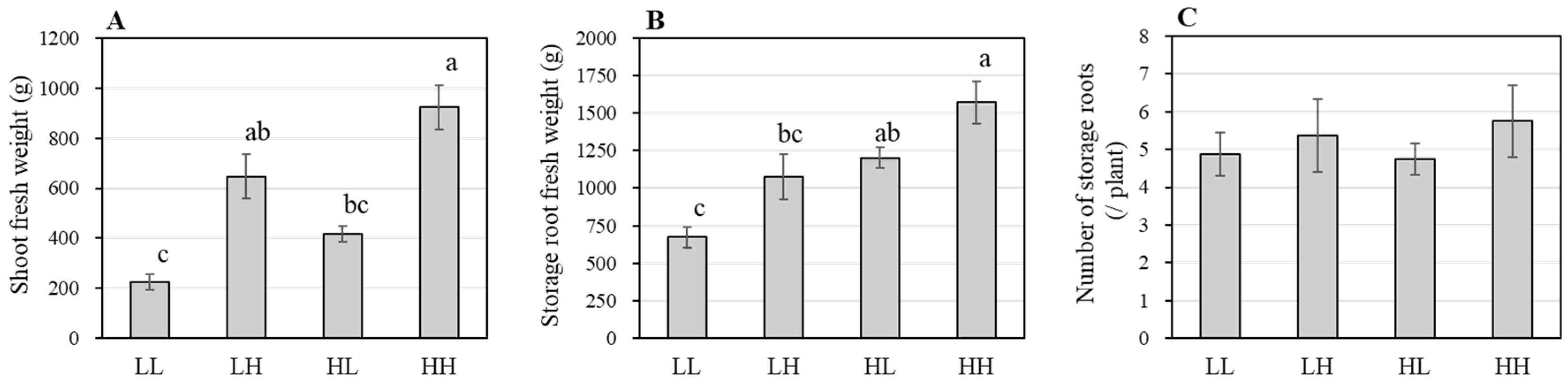
3.1. Effect of NSC on the Growth of Hydroponic Sweetpotato (Experiment One)
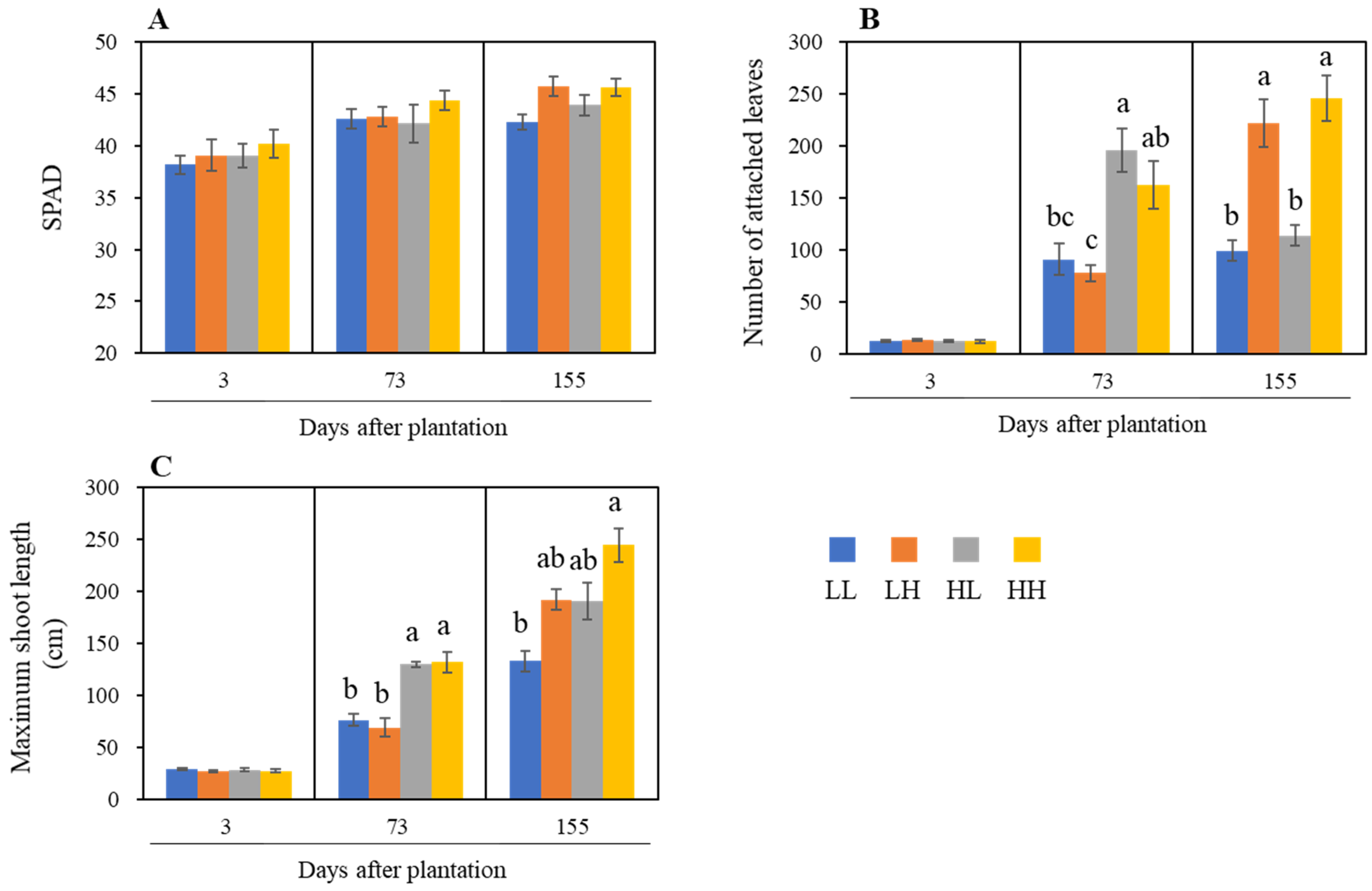
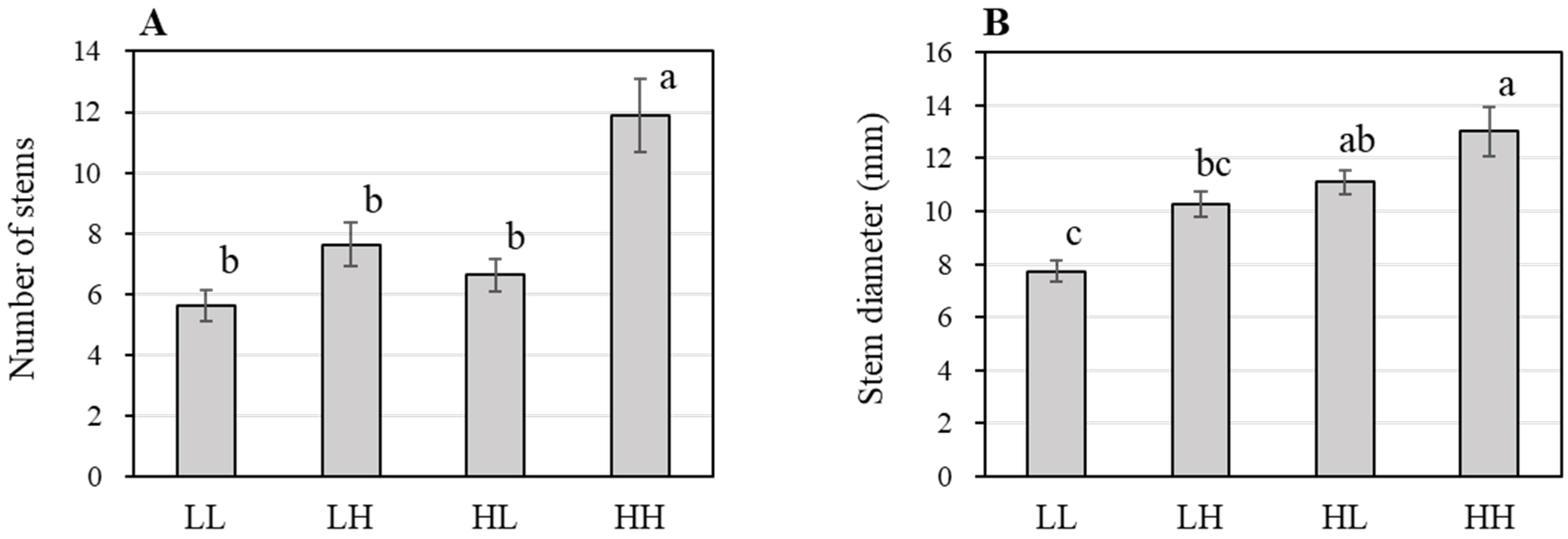
3.2. Effect of Variation in NSC on the Growth of Hydroponic Sweetpotato (Experiment Two)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Petsakos, A.; Prager, S.D.; Gonzalez, C.E.; Gama, A.C.; Sulser, T.B.; Gbegbelegbe, S.; Kikulwe, E.M.; Hareau, G. Understanding the consequences of changes in the production frontiers for roots, tubers and bananas. Glob. Food Secur. 2019, 20, 180–188. [Google Scholar] [CrossRef]
- Woolfe, J. Sweet Potato: An Untapped Food Resource, 1st ed.; Cambridge University Press: New York, NY, USA, 1992; p. 643. [Google Scholar]
- Pimentel, D.; Doughty, R.; Carothers, C.; Lamberson, S.; Bora, N.; Lee, K. Energy inputs in crop production in developing and developed countries. In Food Security and Environmental Quality in the Developing World; CRC Press: Boca Raton, FL, USA, 2002; pp. 129–151. [Google Scholar]
- Koçar, G.; Civaş, N. An overview of biofuels from energy crops: Current status and future prospects. Renew. Sustain. Energy Rev. 2013, 28, 900–916. [Google Scholar] [CrossRef]
- Lareo, C.; Ferrari, M.D. Sweet Potato as a Bioenergy Crop for Fuel Ethanol Production: Perspectives and Challenges. In Bioethanol Production from Food Crops; Academic Press: New York, NY, USA, 2019; pp. 115–147. [Google Scholar]
- Ziska, L.H.; Runion, G.B.; Tomecek, M.; Prior, S.A.; Torbet, H.A.; Sicher, R. An evaluation of cassava, sweet potato and field corn as potential carbohydrate sources for bioethanol production in Alabama and Maryland. Biomass Bioenergy 2009, 33, 1503–1508. [Google Scholar] [CrossRef]
- e Silva, J.O.V.; Almeida, M.F.; da Conceição Alvim-Ferraz, M.; Dias, J.M. Integrated production of biodiesel and bioethanol from sweet potato. Renew. Energy 2018, 124, 114–120. [Google Scholar] [CrossRef]
- Chu, C.Y.; Sen, B.; Lay, C.H.; Lin, Y.C.; Lin, C.Y. Direct fermentation of sweet potato to produce maximal hydrogen and ethanol. Appl. Energy 2012, 100, 10–18. [Google Scholar] [CrossRef]
- Lay, C.H.; Lin, H.C.; Sen, B.; Chu, C.Y.; Lin, C.Y. Simultaneous hydrogen and ethanol production from sweet potato via dark fermentation. J. Clean. Prod. 2012, 27, 155–164. [Google Scholar] [CrossRef]
- Kobayashi, T.; Tang, Y.; Urakami, T.; Morimura, S.; Kida, K. Digestion performance and microbial community in full-scale methane fermentation of stillage from sweet potato-shochu production. J. Environ. Sci. 2014, 26, 423–431. [Google Scholar] [CrossRef]
- Wang, F.; Jiang, Y.; Guo, W.; Niu, K.; Zhang, R.; Hou, S.; Wang, M.; Yi, Y.; Zhu, C.; Jia, C.; et al. An environmentally friendly and productive process for bioethanol production from potato waste. Biotechnol. Biofuels 2016, 9, 50. [Google Scholar] [CrossRef]
- Monteiro, R.L.; de Moraes, J.O.; Domingos, J.D.; Carciofi, B.A.M.; Laurindo, J.B. Evolution of the physicochemical properties of oil-free sweet potato chips during microwave vacuum drying. Innov. Food Sci. Emerg. Technol. 2020, 63, 102317. [Google Scholar] [CrossRef]
- Nedunchezhiyan, M.; Byju, G.; Jata, S.K. Sweet potato agronomy. Fruit Veg. Cereal Sci. Biotechnol. 2012, 6, 1–10. [Google Scholar]
- Halavatau, S.; O’Sullivan, J.N.; Asher, C.J.; Blamey, F.P.C. Better nutrition improves sweet potato and taro yields in the South Pacific. Trop. Agric. 1998, 75, 6–12. [Google Scholar]
- Laxminarayana, K.; John, K.S.; Ravindran, C.S.; Naskar, S.K. Effect of lime, inorganic, and organic sources on soil fertility, yield, quality, and nutrient uptake of sweet potato in Alfisols. Commun. Soil Sci. Plant Anal. 2011, 42, 2515–2525. [Google Scholar] [CrossRef]
- Sawahata, H. Studies on the Characteristics of the Thickening of Storage Root of Sweet Potato: II. Influence of the supply of mineral nutrients on the thickening of storage root. Jpn. J. Crop Sci. 1989, 58, 290–296. [Google Scholar] [CrossRef]
- Osaki, M.; Ueda, H.; Shinano, T.; Matsui, H.; Tadano, T. Accumulation of carbon and nitrogen compounds in sweet potato plants grown under deficiency of N, P, or K nutrients. Soil Sci. Plant Nutr. 1995, 41, 557–566. [Google Scholar] [CrossRef]
- Taranet, P.; Harper, S.; Kirchhof, G.; Fujinuma, R.; Menzies, N. Growth and yield response of glasshouse-and field-grown sweetpotato to nitrogen supply. Nutr. Cycl. Agroecosyst. 2017, 108, 309–321. [Google Scholar] [CrossRef]
- Jia, Z.D.; Ma, P.Y.; Bian, X.F.; Guo, X.D.; Xie, Y.Z. The effects of different N and K fertilizer ratio and planting density on yield and dry matter accumulation of sweetpotato. Acta Agric. Boreali-Sin. 2012, 27, 321–327. [Google Scholar]
- Wang, S.Y.; Li, H.; Liu, Q.; Shi, Y.X. Interactive effects of nitrogen and potassium on root growth and leaf enzyme activities of sweet potato. Acta Agric. Boreali-Sin. 2015, 30, 167–173. [Google Scholar]
- Wang, S.; Liu, Q.; Shi, Y.; Li, H.; Wang, S.; Liu, Q.; Shi, Y.; Li, H. Interactive effects of nitrogen and potassium on photosynthesis product distribution and accumulation of sweetpotato. Sci. Agric. Sin. 2017, 50, 2706–2716. [Google Scholar]
- George, M.S.; Lu, G.; Zhou, W. Genotypic variation for potassium uptake and utilization efficiency in sweet potato (Ipomoea batatas L.). Field Crop. Res. 2002, 77, 7–15. [Google Scholar] [CrossRef]
- Villordon, A.; LaBonte, D.; Firon, N.; Carey, E. Variation in nitrogen rate and local availability alter root architecture attributes at the onset of storage root initiation in ‘Beauregard’ sweetpotato. HortScience 2013, 48, 808–815. [Google Scholar] [CrossRef]
- Villordon, A.Q.; Ginzberg, I.; Firon, N. Root architecture and root and tuber crop productivity. Trends Plant Sci. 2014, 19, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Savvas, D. Hydroponics: A modern technology supporting the application of integrated crop management in greenhouse. J. Food Agric. Environ. 2003, 1, 80–86. [Google Scholar]
- Jones, J.B., Jr. Hydroponics: Its history and use in plant nutrition studies. J. Plant Nutr. 1982, 5, 1003–1030. [Google Scholar] [CrossRef]
- Savvas, D.; Adamidis, K. Automated management of nutrient solutions based on target electrical conductivity, pH, and nutrient concentration ratios. J. Plant Nutr. 1999, 22, 1415–1432. [Google Scholar] [CrossRef]
- Öztekin, G.B.; Uludağ, T.; Tüzel, Y. Growing spinach (Spinacia oleracea L.) in a floating system with different concentrations of nutrient solution. Appl. Ecol. Environ. Res. 2018, 16, 3333–3350. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Watanabe, S.; Nakashima, T.; Okano, K. Effects of salinity at two ripening stages on the fruit quality of single-truss tomato grown in hydroponics. J. Hortic. Sci. Biotechnol. 1999, 74, 690–693. [Google Scholar] [CrossRef]
- Wu, M.; Kubota, C. Effects of electrical conductivity of hydroponic nutrient solution on leaf gas exchange of five greenhouse tomato cultivars. HortTechnology 2008, 18, 271–277. [Google Scholar] [CrossRef]
- Wanzheng, M.; Hanping, M.; Jing, H.; Jiheng, N.; Zhongfang, L. Effects of different nutrient solution concentration on the growth and development of cucumber in greenhouse. In Proceedings of the 2011 International Conference on New Technology of Agricultural, Zibo, China, 27–29 May 2011; IEEE: New York, NY, USA, 2011; pp. 277–281. [Google Scholar]
- Kang, J.G.; van Iersel, M.W. Nutrient solution concentration affects shoot: Root ratio, leaf area ratio, and growth of subirrigated salvia (Salvia splendens). HortScience 2004, 39, 49–54. [Google Scholar] [CrossRef]
- Valdez, M.T.; Ito, T.; Shinohara, Y.; Maruo, T. Effects of nutrient solution levels on the growth, yield and mineral contents in hydroponically-grown bush snap bean. Environ. Control Biol. 2002, 40, 167–175. [Google Scholar] [CrossRef][Green Version]
- Rouphael, Y.; Cardarelli, M.; Lucini, L.; Rea, E.; Colla, G. Nutrient solution concentration affects growth, mineral composition, phenolic acids, and flavonoids in leaves of artichoke and cardoon. HortScience 2012, 47, 1424–1429. [Google Scholar] [CrossRef]
- Hoang, N.N.; Kitaya, Y.; Shibuya, T.; Endo, R. Development of an in vitro hydroponic culture system for wasabi nursery plant production—Effects of nutrient concentration and supporting material on plantlet growth. Sci. Hortic. 2019, 245, 237–243. [Google Scholar] [CrossRef]
- Shinohara, Y.; Suzuki, Y. Effects of light and nutritional conditions on the ascorbic acid content of lettuce. J. Jpn. Soc. Hortic. Sci. 1981, 50, 239–246. [Google Scholar] [CrossRef]
- Fallovo, C.; Rouphael, Y.; Rea, E.; Battistelli, A.; Colla, G. Nutrient solution concentration and growing season affect yield and quality of Lactuca sativa L. var. acephala in floating raft culture. J. Sci. Food Agric. 2009, 89, 1682–1689. [Google Scholar] [CrossRef]
- Sarooshi, R.A.; Cresswell, G.C. Effects of hydroponic solution composition, electrical conductivity and plant spacing on yield and quality of strawberries. Aust. J. Exp. Agric. 1994, 34, 529–535. [Google Scholar] [CrossRef]
- Lieten, P. The effect of nutrition prior to and during flower differentiation on phyllody and plant performance of short day strawberry Elsanta. Acta Hortic. 2002, 567, 345–348. [Google Scholar] [CrossRef]
- Gallace, N.; Boonen, M.; Lieten, P.; Bylemans, D. Electrical conductivity of the nutrient solution: Implications for flowering and yield in day-neutral cultivars. Acta Hortic. 2017, 1156, 223–228. [Google Scholar] [CrossRef]
- Eguchi, T.; Yoshida, S. A cultivation method to ensure tuberous root formation in sweetpotatoes (Ipomoea batatas (L.) Lam.). Environ. Control Biol. 2004, 42, 259–266. [Google Scholar] [CrossRef]
- Eguchi, T.; Moriyama, S.; Miyajima, I.; Yoshida, S.; Chikushi, J. A hydroponic method suitable for tops production of a sweetpotato [Ipomoea batatas] cultivar ‘Suioh’. J. Sci. High Technol. Agric. 2007, 19, 167–174. [Google Scholar] [CrossRef]
- Sakamoto, M.; Suzuki, T. Effect of pot volume on the growth of sweetpotato cultivated in the new hydroponic system. Sustain. Agric. Res. 2018, 7, 137–145. [Google Scholar] [CrossRef]
- Kitaya, Y.; Hirai, H.; Endo, R.; Shibuya, T. Effects of water contents and CO2 concentrations in soil on growth of sweet potato. Field Crops Res. 2013, 152, 36–43. [Google Scholar]
- Eguchi, T.; Ito, Y.; Yoshida, S. Periodical wetting increases α-tocopherol content in the tuberous roots of sweetpotato (Ipomoea batatas (L.) Lam.). Environ. Control Biol. 2012, 50, 297–303. [Google Scholar] [CrossRef][Green Version]
- Kitaya, Y.; Hirai, H.; Wei, X.; Islam, A.F.M.S.; Yamamoto, M. Growth of sweetpotato cultured in the newly designed hydroponic system for space farming. Adv. Space Res. 2008, 41, 730–735. [Google Scholar] [CrossRef]
- Mortley, D.G.; Loretan, P.A.; Bonsi, C.K.; Hill, W.A.; Morris, C.E. Plant spacing influences yield and linear growth rate of sweetpotatoes grown hydroponically. HortScience 1991, 26, 1274–1275. [Google Scholar] [CrossRef]
- Almazan, A.M.; Zhou, X. Biomass yield and composition of sweetpotato grown in a nutrient film technique system. Plant Foods Hum. Nutr. 1997, 50, 259–268. [Google Scholar] [CrossRef]
- Uewada, T. The solution culture of sweet potatoes. Environ. Control Biol. 1990, 28, 135–140. [Google Scholar] [CrossRef]
- Japan Meteorological Agency Official Website. Past Weather Data of Wakayama City in 2018. Available online: http://www.data.jma.go.jp/obd/stats/etrn/view/monthly_s1.php?prec_no=65&block_no=47777&year=2018&month=&day=&view=p1 (accessed on 23 September 2020).
- Japan Meteorological Agency Official Website. Past Weather Data of Wakayama City in 2019. Available online: http://www.data.jma.go.jp/obd/stats/etrn/view/monthly_s1.php?prec_no=65&block_no=47777&year=2019&month=&day=&view=p1 (accessed on 23 September 2020).
- Sonneveld, C.; Voogt, W. Plant Nutrition of Greenhouse Crops; Springer: Dordrecht, The Netherlands, 2009; pp. 393–403. [Google Scholar]
- Trejo-Téllez, L.I.; Gómez-Merino, F.C. Nutrient solutions for hydroponic systems. In Hydroponics: A Standard Methodology for Plant Biological Researches; Asao, T., Ed.; InTechOpen: Rjeka, Croatia, 2012; pp. 1–22. [Google Scholar]
- Dewir, Y.H.; Chakrabarty, D.; Ali, M.B.; Hahn, E.J.; Paek, K.Y. Effects of hydroponic solution EC, substrates, PPF and nutrient scheduling on growth and photosynthetic competence during acclimatization of micropropagated Spathiphyllum plantlets. Plant Growth Regul. 2005, 46, 241–251. [Google Scholar] [CrossRef]
- Tabatabaie, S.J.; Nazari, J. Influence of nutrient concentrations and NaCl salinity on the growth, photosynthesis, and essential oil content of peppermint and lemon verbena. Turk. J. Agric. For. 2007, 31, 245–253. [Google Scholar]
- Chang, D.C.; Cho, I.C.; Suh, J.T.; Kim, S.J.; Lee, Y.B. Growth and yield response of three aeroponically grown potato cultivars (Solanum tuberosum L.) to different electrical conductivities of nutrient solution. Am. J. Potato Res. 2011, 88, 450–458. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Kelm, M.; Brück, H.; Hermann, M.; Sattelmacher, B. Plant Productivity and Water Use Efficiency of Sweetpotato (Ipomoea batatas) as Affected by Nitrogen Supply; CIP Program Report 1999–2000; International Potato Center: Lima, Peru, 2000; pp. 273–279. [Google Scholar]
- Osaki, M.; Ueda, H.; Shinano, T.; Matsui, H.; Tadano, T. Accumulation of carbon and nitrogen compounds in sweet potato plants grown under different nitrogen application rates. Soil Sci. Plant Nutr. 1995, 41, 547–555. [Google Scholar] [CrossRef]
- Manzocco, L.; Foschia, M.; Tomasi, N.; Maifreni, M.; Dalla Costa, L.; Marino, M.; Cortella, G.; Cesco, S. Influence of hydroponic and soil cultivation on quality and shelf life of ready-to-eat lamb’s lettuce (Valerianella locusta L. Laterr). J. Sci. Food Agric. 2011, 91, 1373–1380. [Google Scholar] [CrossRef]
- Miller, M.H.; Walker, G.K.; Tollenaar, M.; Alexander, K.G. Growth and yield of maize (Zea mays L.) grown outdoors hydroponically and in soil. Can. J. Soil Sci. 1989, 69, 295–302. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C. Enhancing nitrogen use efficiency in crop plants. Adv. Agron. 2005, 88, 97–185. [Google Scholar]
- Yoshida, Y.; Fujime, Y.; Chujo, T. Effects of nitrogen nutrition on flower bud development and fruit malformation in ‘Ai-Berry’ strawberry. J. Jpn. Soc. Hortic. Sci. 1992, 60, 869–879. [Google Scholar] [CrossRef][Green Version]
- Dittakit, P.; Thongket, T. Increased nutrient solution concentration during early fruit development stages enhances pungency and phenylalanine ammonia-lyase activity in hot chili (Capsicum annuum L.). Am. J. Agric. Biol. Sci. 2014, 9, 72. [Google Scholar] [CrossRef]
- Maruyama, S.; Ishigami, Y.; Goto, E. Effect of nutrient solution concentration at the heading time on the growth, development, and seed storage protein content of rice plants in a controlled environment. Environ. Control Biol. 2010, 48, 17–24. [Google Scholar] [CrossRef]
- Wu, M.; Kubota, C. Effects of high electrical conductivity of nutrient solution and its application timing on lycopene, chlorophyll and sugar concentrations of hydroponic tomatoes during ripening. Sci. Hortic. 2008, 116, 122–129. [Google Scholar] [CrossRef]
- Hochmuth, G.J.; Hochmuth, R.C. Nutrient Solution Formulation for Hydroponic (Perlite, Rockwool, NFT) Tomatoes in Florida; HS796; University of Florida Cooperative Extension Service: Gainesville, FL, USA, 2001. [Google Scholar]
- Ankumah, R.O.; Khan, V.; Mwamba, K.; Kpomblekou-A, K. The influence of source and timing of nitrogen fertilizers on yield and nitrogen use efficiency of four sweet potato cultivars. Agric. Ecosyst. Environ. 2003, 100, 201–207. [Google Scholar] [CrossRef]
- Du, X.; Xi, M.; Kong, L. Split application of reduced nitrogen rate improves nitrogen uptake and use efficiency in sweetpotato. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Du, X.; Zhang, X.; Xi, M.; Kong, L. Split application enhances sweetpotato starch production by regulating the conversion of sucrose to starch under reduced nitrogen supply. Plant Physiol. Biochem. 2020, 151, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.B.; Warren, J.G.; Mullins, G.L. Nitrogen rate and application timing affect ‘Beauregard’ sweetpotato yield and quality. HortScience 2005, 40, 214–217. [Google Scholar] [CrossRef]
- Patharkar, O.R.; Walker, J.C. Connections between abscission, dehiscence, pathogen defense, drought tolerance, and senescence. Plant Sci. 2019, 284, 25–29. [Google Scholar] [CrossRef]
- Liljegren, S.J. Organ abscission: Exit strategies require signals and moving traffic. Curr. Opin. Plant Biol. 2012, 15, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Patterson, S.E.; Bolivar-Medina, J.L.; Falbel, T.G.; Hedtcke, J.L.; Nevarez-McBride, D.; Maule, A.F.; Zalapa, J.E. Are we on the right track: Can our understanding of abscission in model systems promote or derail making improvements in less studied crops? Front. Plant Sci. 2016, 6, 1268. [Google Scholar] [CrossRef]
- Patharkar, O.R.; Walker, J.C. Advances in abscission signaling. J. Exp. Bot. 2018, 69, 733–740. [Google Scholar] [CrossRef]
- Somda, Z.C.; Kays, S.J. Sweet potato canopy morphology: Leaf distribution. J. Am. Soc. Hortic. Sci. 1990, 115, 39–45. [Google Scholar] [CrossRef]
- Somda, Z.C.; Mahomed, M.T.M.; Kays, S.J. Analysis of leaf shedding and dry matter recycling in sweetpotato. J. Plant Nutr. 1991, 14, 1201–1212. [Google Scholar] [CrossRef]
- McLaurin, W.J.; Kays, S.J. Substantial leaf shedding—A consistent phenomenon among high-yielding sweetpotato cultivars. HortScience 1993, 28, 826–827. [Google Scholar] [CrossRef]
- Lynch, J.; Brown, K.M. Ethylene and plant responses to nutritional stress. Physiol. Plant. 1997, 100, 613–619. [Google Scholar] [CrossRef]
- Bouwkamp, J.C. Sweet potato products: A Natural Resource for the Tropics; CRC Press: Boca Taton, FL, USA, 1985. [Google Scholar]
- Matile, P.; Hortensteiner, S.; Thomas, H.; Krautler, B. Chlorophyll breakdown in senescent leaves. Plant Physiol. 1996, 112, 1403–1409. [Google Scholar] [CrossRef]
- Huang, Z.A.; Jiang, D.A.; Yang, Y.; Sun, J.W.; Jin, S.H. Effects of nitrogen deficiency on gas exchange, chlorophyll fluorescence, and antioxidant enzymes in leaves of rice plants. Photosynthetica 2004, 42, 357–364. [Google Scholar] [CrossRef]
- Sakamoto, M.; Munemura, I.; Tomita, R.; Kobayashi, K. Involvement of hydrogen peroxide in leaf abscission signaling, revealed by analysis with an in vitro abscission system in Capsicum plants. Plant J. 2008, 56, 13–27. [Google Scholar] [CrossRef]
- Sakamoto, M.; Munemura, I.; Tomita, R.; Kobayashi, K. Reactive oxygen species in leaf abscission signaling. Plant Signal. Behav. 2008, 3, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Bar-Dror, T.; Dermastia, M.; Kladnik, A.; Žnidarič, M.T.; Novak, M.P.; Meir, S.; Burd, S.; Philosoph-Hadas, S.; Ori, N.; Sonego, L.; et al. Programmed cell death occurs asymmetrically during abscission in tomato. Plant Cell 2011, 23, 4146–4163. [Google Scholar] [CrossRef]
- Liao, W.; Wang, G.; Li, Y.; Wang, B.; Zhang, P.; Peng, M. Reactive oxygen species regulate leaf pulvinus abscission zone cell separation in response to water-deficit stress in cassava. Sci. Rep. 2016, 6, 1–17. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Number of Total Leaves | Abscised Leaf Ratio (%) |
|---|---|---|
| LL | 277 c | 63.6 ab |
| LH | 545 ab | 58.9 c |
| HL | 417 bc | 72.9 a |
| HH | 761 a | 66.9 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakamoto, M.; Suzuki, T. Effect of Nutrient Solution Concentration on the Growth of Hydroponic Sweetpotato. Agronomy 2020, 10, 1708. https://doi.org/10.3390/agronomy10111708
Sakamoto M, Suzuki T. Effect of Nutrient Solution Concentration on the Growth of Hydroponic Sweetpotato. Agronomy. 2020; 10(11):1708. https://doi.org/10.3390/agronomy10111708
Chicago/Turabian StyleSakamoto, Masaru, and Takahiro Suzuki. 2020. "Effect of Nutrient Solution Concentration on the Growth of Hydroponic Sweetpotato" Agronomy 10, no. 11: 1708. https://doi.org/10.3390/agronomy10111708
APA StyleSakamoto, M., & Suzuki, T. (2020). Effect of Nutrient Solution Concentration on the Growth of Hydroponic Sweetpotato. Agronomy, 10(11), 1708. https://doi.org/10.3390/agronomy10111708