Melon Genetic Resources Characterization for Rind Volatile Profile

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Cultivation and Sampling
2.3. Analysis Conditions
2.4. Data Processing and Statistical Analysis
3. Results and Discussion
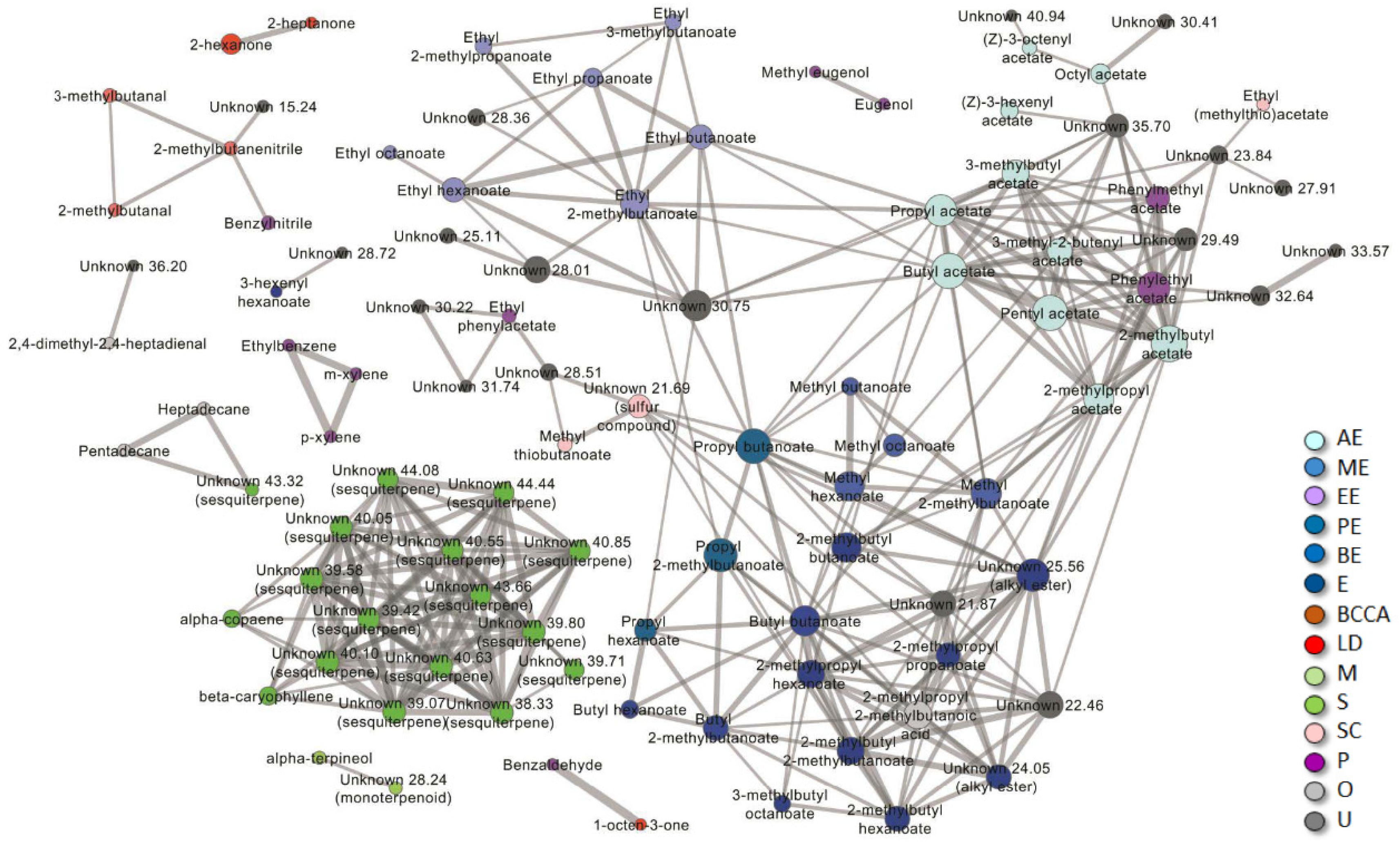
3.1. Melon Rind Volatile Profile
3.2. Rind vs. Flesh VOCs Profile
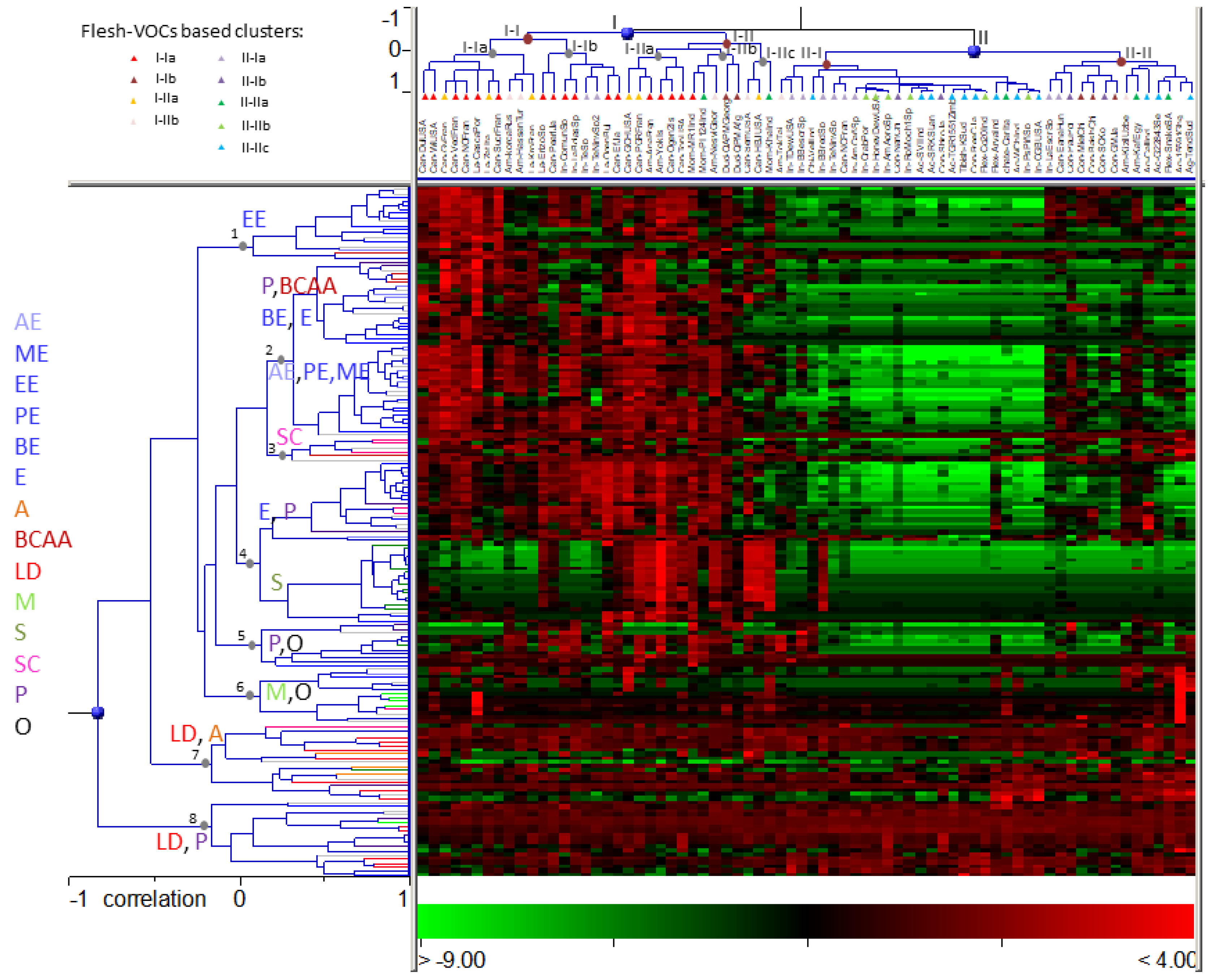
3.3. Melon Core Collection Characterization Based on Rind VOCs
3.3.1. Aromatic Accessions (cluster I)
3.3.2. Low or Non-Aromatic Accessions (cluster II)
3.4. Perspectives in Elucidation of Genomic Regions Involved
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pitrat, M. Melon Genetic Resources: Phenotypic Diversity and Horticultural Taxonomy. In Genetics and Genomics of Setaria; Springer Science and Business Media LLC: Cham, Switzerland, 2016; Volume 20, pp. 25–60. [Google Scholar]
- Stepansky, A.; Kovalski, I.; Perl-Treves, R. Intraspecific classification of melons (Cucumis melo L.) in view of their phenotypic and molecular variation. Plant Syst. Evol. 1999, 217, 313–332. [Google Scholar] [CrossRef]
- Burger, Y.; Sa’Ar, U.; Paris, H.; Lewinsohn, E.; Katzir, N.; Tadmor, Y.; Schaffer, A. Genetic variability for valuable fruit quality traits in Cucumis melo. Isr. J. Plant Sci. 2006, 54, 233–242. [Google Scholar] [CrossRef]
- Gur, A.; Gonda, I.; Portnoy, V.; Tzuri, G.; Chayut, N.; Cohen, S.; Yeselson, Y.; Meir, A.; Bar, E.; Davidovitz-Rikanati, R.; et al. Genomic Aspects of Melon Fruit Quality. In Genetics and Genomics of Setaria; Springer Science and Business Media LLC: Cham, Switzerland, 2016; Volume 20, pp. 377–408. [Google Scholar]
- Esteras, C.; Rambla, J.L.; Sánchez, G.; López-Gresa, M.P.; González-Mas, M.C.; Fernández-Trujillo, J.P.; Bellés, J.M.; Granell, A.; Picó, M.B. Fruit flesh volatile and carotenoid profile analysis within the Cucumis melo L. species reveals unexploited variability for future genetic breeding. J. Sci. Food Agric. 2018, 98, 3915–3925. [Google Scholar] [CrossRef] [PubMed]
- Moing, A.; Allwood, J.W.; Aharoni, A.; Baker, J.; Beale, M.H.; Ben-Dor, S.; Biais, B.; Brigante, F.; Burger, Y.; Deborde, C.; et al. Comparative Metabolomics and Molecular Phylogenetics of Melon (Cucumis melo, Cucurbitaceae) Biodiversity. Metabolimics 2020, 10, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nee, M.; Kirkbride, J.H. Biosystematic Monograph of the Genus Cucumis (Cucurbitaceae)-Botanical Identification of Cucumbers and Melons. Bull. Torrey Bot. Club 1994, 121, 300. [Google Scholar] [CrossRef]
- Pitrat, M. Melon Cucumis melo L. In Handbook of Crop Breeding, Vol I: Vegetables, 1st ed.; Prohens, J., Nuez, F., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2008; Volume 1, pp. 283–315. [Google Scholar]
- Bernillon, S.; Biais, B.; Deborde, C.; Maucourt, M.; Cabasson, C.; Gibon, Y.; Hansen, T.H.; Husted, S.; De Vos, R.C.H.; Mumm, R.; et al. Metabolomic and elemental profiling of melon fruit quality as affected by genotype and environment. Metabolomics 2012, 9, 57–77. [Google Scholar] [CrossRef]
- Granell, A.; Rambla, J.L. Biosynthesis of volatile compounds (Chapter 6). In The Molecular Biology and Biochemistry of Fruit Ripening, 1st ed.; Seymour, G.B., Tucker, G.A., Poole, M., Giovannoni, J.J., Eds.; John Wiley & Sons, Inc.: Oxford, UK, 2013; pp. 135–161. [Google Scholar]
- Aubert, C.; Bourger, N. Investigation of Volatiles in Charentais Cantaloupe Melons (Cucumis melo Var. cantalupensis). Characterization of Aroma Constituents in Some Cultivars. J. Agric. Food Chem. 2004, 52, 4522–4528. [Google Scholar] [CrossRef]
- Aubert, C.; Pitrat, M. Volatile Compounds in the Skin and Pulp of Queen Anne’s Pocket Melon. J. Agric. Food Chem. 2006, 54, 8177–8182. [Google Scholar] [CrossRef]
- Obando-Ulloa, J.M.; Moreno, E.; Garcia-Mas, J.; Nicolai, B.; Lammertyn, J.; Monforte, A.J.; Fernández-Trujillo, J.P. Climacteric or non-climacteric behavior in melon fruit. Postharvest Biol. Technol. 2008, 49, 27–37. [Google Scholar] [CrossRef]
- Obando-Ulloa, J.M.; Ruiz, J.; Monforte, A.J.; Fernández-Trujillo, J.P.; Ruiz-Carrascal, J. Aroma profile of a collection of near-isogenic lines of melon (Cucumis melo L.). Food Chem. 2010, 118, 815–822. [Google Scholar] [CrossRef] [Green Version]
- Verzera, A.; Dima, G.; Tripodi, G.; Ziino, M.; Lanza, C.M.; Mazzaglia, A. Fast Quantitative Determination of Aroma Volatile Constituents in Melon Fruits by Headspace–Solid-Phase Microextraction and Gas Chromatography–Mass Spectrometry. Food Anal. Methods 2010, 4, 141–149. [Google Scholar] [CrossRef]
- Condurso, C.; Verzera, A.; Dima, G.; Tripodi, G.; Crinò, P.; Paratore, A.; Romano, D. Effects of different rootstocks on aroma volatile compounds and carotenoid content of melon fruits. Sci. Hortic. 2012, 148, 9–16. [Google Scholar] [CrossRef]
- Escribano, S.; Lázaro, A. Sensorial characteristics of Spanish traditional melon genotypes: Has the flavor of melon changed in the last century? Eur. Food Res. Technol. 2012, 234, 581–592. [Google Scholar] [CrossRef]
- Pang, X.; Chen, D.; Hu, X.; Zhang, Y.; Wu, J. Verification of Aroma Profiles of Jiashi Muskmelon Juice Characterized by Odor Activity Value and Gas Chromatography–Olfactometry/Detection Frequency Analysis: Aroma Reconstitution Experiments and Omission Tests. J. Agric. Food Chem. 2012, 60, 10426–10432. [Google Scholar] [CrossRef]
- Vallone, S.; Sivertsen, H.; Anthon, G.E.; Barrett, D.M.; Mitcham, E.J.; Ebeler, S.E.; Negre-Zakharov, F. An integrated approach for flavour quality evaluation in muskmelon (Cucumis melo L. reticulatus group) during ripening. Food Chem. 2013, 139, 171–183. [Google Scholar] [CrossRef]
- Bai, X.-H.; Teng, L.-H.; Lü, D.-Q.; Qi, H.-Y. Co-Treatment of EFF and 1-MCP for Enhancing the Shelf-Life and Aroma Volatile Compounds of Oriental Sweet Melons (Cucumis melo var. makuwa Makino). J. Integr. Agric. 2014, 13, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Cao, S.; Jin, Y.; Tang, Y.; Qi, H. The Relationship between CmADHs and the Diversity of Volatile Organic Compounds of Three Aroma Types of Melon (Cucumis melo). Front. Physiol. 2016, 7, 254. [Google Scholar] [CrossRef] [Green Version]
- Gonda, I.; Lev, S.; Bar, E.; Sikron, N.; Portnoy, V.; Davidovich-Rikanati, R.; Burger, J.; Schaffer, A.A.; Tadmor, Y.; Giovannonni, J.J.; et al. Catabolism ofl-methionine in the formation of sulfur and other volatiles in melon (Cucumis melo L.) fruit. Plant J. 2013, 74, 458–472. [Google Scholar] [CrossRef]
- Freilich, S.; Lev, S.; Gonda, I.; Reuveni, E.; Portnoy, V.; Oren, E.; Lohse, M.; Galpaz, N.; Bar, E.; Tzuri, G.; et al. Systems approach for exploring the intricate associations between sweetness, color and aroma in melon fruits. BMC Plant Biol. 2015, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Gonda, I.; Burger, Y.; Schaffer, A.A.; Ibdah, M.; Tadmor, Y.; Katzir, N.; Fait, A.; Lewinsohn, E. Biosynthesis and perception of melon aroma. In Biotechnology in Flavor Production; Wiley: Hoboken, NJ, USA, 2016; pp. 281–305. [Google Scholar]
- Gonda, I.; Davidovich-Rikanati, R.; Bar, E.; Lev, S.; Jhirad, P.; Meshulam, Y.; Wissotsky, G.; Portnoy, V.; Burger, J.; Schaffer, A.A.; et al. Differential metabolism of L–phenylalanine in the formation of aromatic volatiles in melon (Cucumis melo L.) fruit. Phytochemistry 2018, 148, 122–131. [Google Scholar] [CrossRef]
- Galpaz, N.; Gonda, I.; Shem-Tov, D.; Barad, O.; Tzuri, G.; Lev, S.; Fei, Z.; Xu, Y.; Mao, L.; Jiao, C.; et al. Deciphering genetic factors that determine melon fruit-quality traits using RNA -Seq-based high-resolution QTL and eQTL mapping. Plant J. 2018, 94, 169–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feder, A.; Jiao, C.; Galpaz, N.; Vrebalov, J.T.; Xu, Y.; Portnoy, V.; Tzuri, G.; Gonda, I.; Burger, Y.; Gur, A.; et al. Melon ethylene-mediated transcriptome and methylome dynamics provide insights to volatile production. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Perpiñá, G.; Roselló, S.; Esteras, C.; Beltrán, J.; Monforte, A.J.; Cebolla-Cornejo, J.; Picó, B. Analysis of aroma-related volatile compounds affected by ‘Ginsen Makuwa’ genomic regions introgressed in ‘Vedrantais’ melon background. Sci. Hortic. in press.
- El-Sharkawy, I.; Manriquez, D.; Flores, F.B.; Regad, F.; Bouzayen, M.; Latché, A.; Pech, J.-C. Functional Characterization of a Melon Alcohol Acyl-transferase Gene Family Involved in the Biosynthesis of Ester Volatiles. Identification of the Crucial Role of a Threonine Residue for Enzyme Activity. Plant Mol. Biol. 2005, 59, 345–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, P.L.; Wang, Y.; Lin, J. Analysis of honeydew melon (Cucumis melo var. inodorus) flavour and GC-MS/MS identification of (E,Z)-2,6-nonadienyl acetate. Flavour Fragr. J. 2009, 24, 341–347. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; Quirantes-Piné, R.; Fernandez-Gutierrez, A.; Segura-Carretero, A. Comparative characterization of phenolic and other polar compounds in Spanish melon cultivars by using high-performance liquid chromatography coupled to electrospray ionization quadrupole-time of flight mass spectrometry. Food Res. Int. 2013, 54, 1519–1527. [Google Scholar] [CrossRef] [Green Version]
- Allwood, J.W.; Cheung, W.; Xu, Y.; Mumm, R.; De Vos, R.C.; Deborde, C.; Biais, B.; Maucourt, M.; Berger, Y.; Schaffer, A.A.; et al. Metabolomics in melon: A new opportunity for aroma analysis. Phytochemistry 2014, 99, 61–72. [Google Scholar] [CrossRef]
- Portnoy, V.; Benyamini, Y.; Bar, E.; Harel-Beja, R.; Gepstein, S.; Giovannoni, J.J.; Schaffer, A.A.; Burger, J.; Tadmor, Y.; Lewinsohn, E.; et al. The molecular and biochemical basis for varietal variation in sesquiterpene content in melon (Cucumis melo L.) rinds. Plant Mol. Biol. 2008, 66, 647–661. [Google Scholar] [CrossRef]
- Esteras, C.; Formisano, G.; Roig, C.; Díaz, A.; Blanca, J.; Garcia-Mas, J.; Gómez-Guillamón, M.; López-Sesé, A.I.; Lázaro, A.; Monforte, A.J.; et al. SNP genotyping in melons: Genetic variation, population structure, and linkage disequilibrium. Theor. Appl. Genet. 2013, 126, 1285–1303. [Google Scholar] [CrossRef]
- Leida, C.; Moser, C.; Esteras, C.; Sulpice, R.; Lunn, J.E.; De Langen, F.; Monforte, A.J.; Picó, M.B. Variability of candidate genes, genetic structure and association with sugar accumulation and climacteric behavior in a broad germplasm collection of melon (Cucumis melo L.). BMC Genet. 2015, 16, 28. [Google Scholar] [CrossRef] [Green Version]
- Sabato, D.; Esteras, C.; Grillo, O.; Picó, M.B.; Bacchetta, G. Seeds morpho-colourimetric analysis as complementary method to molecular characterization of melon diversity. Sci. Hortic. 2015, 192, 441–452. [Google Scholar] [CrossRef]
- Sánchez, G.; Martínez, J.; Romeu, J.; García, J.; Monforte, A.J.; Badenes, M.L.; Granell, A. The peach volatilome modularity is reflected at the genetic and environmental response levels in a QTL mapping population. BMC Plant Biol. 2014, 14, 1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, G.; Besada, C.; Badenes, M.L.; Monforte, A.J.; Granell, A. A Non-Targeted Approach Unravels the Volatile Network in Peach Fruit. PLoS ONE 2012, 7, e38992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorrilla-Fontanesi, Y.; Rambla, J.L.; Cabeza, A.; Medina, J.J.; Sánchez-Sevilla, J.F.; Valpuesta, V.; Botella, J.R.; Granell, A.; Amaya, I. Genetic Analysis of Strawberry Fruit Aroma and Identification of O-Methyltransferase FaOMT as the Locus Controlling Natural Variation in Mesifurane Content. Plant Physiol. 2012, 159, 851–870. [Google Scholar] [CrossRef] [Green Version]
- Rambla, J.L.; Medina, A.; Fernández-Del-Carmen, A.; Barrantes, W.; Grandillo, S.; Cammareri, M.; López-Casado, G.; Rodrigo, G.; Alonso, A.; García-Martínez, S.; et al. Identification, introgression, and validation of fruit volatile QTLs from a red-fruited wild tomato species. J. Exp. Bot. 2016, 68, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Verzera, A.; Dima, G.; Tripodi, G.; Condurso, C.; Crinò, P.; Romano, D.; Mazzaglia, A.; Lanza, C.; Restuccia, C.; Paratore, A. Aroma and sensory quality of honeydew melon fruits (Cucumis melo L. subsp. melo var. inodorus H. Jacq.) in relation to different rootstocks. Sci. Hortic. 2014, 169, 118–124. [Google Scholar] [CrossRef]
- Dogimont, C. Gene List for Melon. Cucurbit Genet. Coop. Rep. 2011, 33–34, 104–133. [Google Scholar]
- López, C.; Ferriol, M.; Picó, M.B. Mechanical transmission of Tomato leaf curl New Delhi virus to cucurbit germplasm: Selection of tolerance sources in Cucumis melo. Euphytica 2015, 204, 679–691. [Google Scholar] [CrossRef]
- Sharon-Asa, L.; Shalit, M.; Frydman, A.; Bar, E.; Holland, D.; Or, E.; Lavi, U.; Lewinsohn, E.; Eyal, Y. Citrus fruit flavor and aroma biosynthesis: Isolation, functional characterization, and developmental regulation of Cstps1, a key gene in the production of the sesquiterpene aroma compound valencene. Plant J. 2003, 36, 664–674. [Google Scholar] [CrossRef]
- Pechous, S.W.; Whitaker, B.D. Cloning and functional expression of an (E, E)-a-farnesene synthase cDNA from peel tissue of apple fruit. Planta 2004, 219, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, T.; Ito, M.; Honda, G. Molecular Cloning, Functional Expression and Characterization of (E)-.BETA.-Farnesene Synthase from Citrus junos. Biol. Pharm. Bull. 2001, 24, 1171–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenço, A.M.; Haddi, K.; Ribeiro, B.M.; Corrêia, R.F.T.; Tomé, H.V.V.; Santos-Amaya, O.; Pereira, E.J.G.; Guedes, R.N.C.; Santos, G.R.; Oliveira, E.E.; et al. Essential oil of Siparuna guianensis as an alternative tool for improved lepidopteran control and resistance management practices. Sci. Rep. 2018, 8, 7215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapp, S.C.; Croteau, R.B. Genomic organization of plant terpene synthases and molecular evolutionary implications. Genetics 2001, 158, 811–832. [Google Scholar]
- Monforte, A.J.; Garcia-Mas, J.; Arús, P. Genetic variability in melon based on microsatellite variation. Plant Breed. 2003, 122, 153–157. [Google Scholar] [CrossRef]
- Blanca, J.; Esteras, C.; Ziarsolo, P.; Perez, D.; Fernã¡ndez-Pedrosa, V.; Collado, C.; De Pablos, R.R.; Ballester, A.; Roig, C.; Cañizares, J.; et al. Transcriptome sequencing for SNP discovery across Cucumis melo. BMC Genom. 2012, 13, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Lian, Q.; Zhang, Z.; Fu, Q.; He, Y.; Ma, S.; Ruggieri, V.; Monforte, A.J.; Wang, P.; Julca, I.; et al. A comprehensive genome variation map of melon identifies multiple domestication events and loci influencing agronomic traits. Nat. Genet. 2019, 51, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, M.J.; Díaz, A.; Dhillon, N.P.; Reddy, U.K.; Picó, M.B.; Monforte, A.J. Re-evaluation of the role of Indian germplasm as center of melon diversification based on genotyping-by-sequencing analysis. BMC Genom. 2019, 20, 448. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, R.G. Phenylpropenes: Occurrence, Distribution, and Biosynthesis in Fruit. J. Agric. Food Chem. 2016, 66, 2259–2272. [Google Scholar] [CrossRef]
- Liu, Z.; Smagghe, G.; Lei, Z.; Wang, J.-J. Identification of Male- and Female-Specific Olfaction Genes in Antennae of the Oriental Fruit Fly (Bactrocera dorsalis). PLoS ONE 2016, 11, e0147783. [Google Scholar] [CrossRef] [Green Version]
- Castro, G.; Perpiñá, G.; Esteras, C.; Armengol, J.; Picó, B.; Pérez-De-Castro, A. Resistance in melon to Monosporascus cannonballus and M. eutypoides: Fungal pathogens associated with Monosporascus root rot and vine decline. Ann. Appl. Biol. 2020, 177, 101–111. [Google Scholar] [CrossRef]
- Linhares, C.M.D.S.; Ambrósio, M.M.Q.; Castro, G.; Torres, S.B.; Esteras, C.; Nunes, G.H.D.S.; Picó, M.B. Effect of temperature on disease severity of charcoal rot of melons caused by Macrophomina phaseolina: Implications for selection of resistance sources. Eur. J. Plant Pathol. 2020, 158, 431–441. [Google Scholar] [CrossRef]
- Castro, G.; Perpiñá, G.; Monforte, A.J.; Picó, M.B.; Esteras, C. New melon introgression lines in a Piel de Sapo genetic background with desirable agronomical traits from dudaim melons. Euphytica 2019, 215, 169. [Google Scholar] [CrossRef]
- Perpiñá, G.; Esteras, C.; Gibon, Y.; Monforte, A.J.; Picó, B. A new genomic library of melon introgression lines in a cantaloupe genetic background for dissecting desirable agronomical traits. BMC Plant Biol. 2016, 16, 154. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L. Genetic Dissection of Fruit Quality and Ripening Traits in Melon. Doctoral Thesis, Universitat Autònoma de Barcelona, Barcelona, Spain, 2018. [Google Scholar]



{kind=link}
{kind=link}
| Code | Local name/germplasm bank name | subsp. | Group | Origin |
|---|---|---|---|---|
| Ac-G22843Se | G22843 (PI 436534) 1 | agrestis | Acidulus | Senegal |
| Ac-TGR1551Zimb | TGR1551 (PI482420) 1 | agrestis | Acidulus | Zimbabwe |
| Ac-SRKSLan | SRK 2 | agrestis | Acidulus | Sri Lanka |
| Ac-SVIInd | SVI 2 | agrestis | Acidulus | India |
| Chi-VellInd | Velleri (PI 164320) 1 | agrestis | Chito | India |
| Con-GMJa | Ginsen Makuwa (PI 420176 ) 1 | agrestis | Conomon-Makuwa-Chinensis | Japan |
| Con-BaishChi | Baishami 2 | agrestis | Conomon-Makuwa-Chinensis | China |
| Con-FreeCJa | Freeman’s Cucumber 3 | agrestis | Conomon-Makuwa-Chinensis | Japan |
| Con-MielChi | Miel Blanc 3 | agrestis | Conomon-Makuwa-Chinensis | China |
| Con-NanChi | Nanbukin 2 | agrestis | Conomon-Makuwa-Chinensis | China |
| Con-PauPol | Paul 2 | agrestis | Conomon-Makuwa-Chinensis | Poland |
| Con-SCKo | Songwhan Charmi (PI 161375) 1 | agrestis | Conomon-Makuwa-Chinensis | Korea |
| Con-ShiroJa | Shiro Uri Okayama 3 | agrestis | Conomon-Makuwa-Chinensis | Japan |
| Mom-KhaInd | Kharbuja (CUM 438) 4 | agrestis | Momordica | India |
| Mom-PI124Ind | PI 124112 1 | agrestis | Momordica | India |
| Mom-MR1Ind | MR1 3 | agrestis | Momordica | India |
| Tibish-KSud | Tibish Khurtagat 2 | agrestis | Tibish | Sudan |
| Ag-15591Gha | PI 185111 1 | agrestis | wild melon | Ghana |
| Ag-CallInd | Callosus 3 | agrestis | wild melon | India |
| Ag-TendSud | Tendelti 2 | agrestis | wild melon | Sudan |
| Ag-WChInd | Wild Chibbar 3 | agrestis | wild melon | India |
| Am-KafEgy | Kafr Hakim (PI 288233) 1 | melo | Ameri | Egypt |
| Am-korcaRus | Korça (CUM 168) 4 | melo | Ameri | Russia |
| Am-TokTaj | Tokash 3 | melo | Ameri | Tajikistan |
| Am-HassanTur | Hassanbey (PI 169368) 1 | melo | Ameri | Turkey |
| Am-KizilUzbe | Kizil-uruk 3 | melo | Ameri | Uzbekistan |
| Am-AnaFran | Ananas ’D’Amerique 2 | melo | Ameri | France |
| Am-NesviGeor | Mucha Nesvi 2 | melo | Ameri | Georgia |
| Am-YokIs | Yokneam 2 | melo | Ameri | Israel |
| Can-EanaHun | Ezüst Ananasz (CUM 305) 4 | melo | Cantalupensis-Reticulatus | Hungary |
| Can-PearlJa | Pearl (PI 266947) 1 | melo | Cantalupensis-Reticulatus | Japan |
| Can-GCHUSA | Golden Champlain (CUM 474) 4 | melo | Cantalupensis-Reticulatus | USA |
| Can-TopUSA | Topmark 3 | melo | Cantalupensis-Reticulatus | USA |
| Can-HBJUSA | Ar Hale’s Best Jumbo 2 | melo | Cantalupensis-Reticulatus | USA |
| Can-DulUSA | Dulce 3 | melo | Cantalupensis-Reticulatus | USA |
| Can-Ogen2Is | Dvash ha ogen 3 | melo | Cantalupensis-Reticulatus | Israel |
| Can-EfJa | Earl’s Favourite 2 | melo | Cantalupensis-Reticulatus | Japan |
| Can-GyFran | Gynadou 2 | melo | Cantalupensis-Reticulatus | France |
| Can-NOFran | Nantais Oblong 2 | melo | Cantalupensis-Reticulatus | France |
| Can-NCFran | Noir des Carmes 3 | melo | Cantalupensis-Reticulatus | France |
| Can-PGRFran | Petit Gris de Rennes 3 | melo | Cantalupensis-Reticulatus | France |
| Can-SemUSA | Seminole 3 | melo | Cantalupensis-Reticulatus | USA |
| Can-SucrFran | Sucrin de Tours 2 | melo | Cantalupensis-Reticulatus | France |
| Can-VedFran | Vedrantais 3 | melo | Cantalupensis-Reticulatus | France |
| Can-WiUSA | Wi-998 3 | melo | Cantalupensis-Reticulatus | USA |
| Chate-CarIta | Carosello (CUM 363) 4 | melo | Chate | Italy |
| Dud-QAPMGeorg | Queen Ann’s Pocket Melon (PI 273438) 1 | melo | Dudaim | Georgia |
| Dud-QPMAfg | Queen’s pocket melon 2 | melo | Dudaim | Afganistan |
| Flex-Co20Ind | Snakemelon (CUM 225) 4 | melo | Flexuosus | India |
| Flex-AryaInd | Arya 3 | melo | Flexuosus | India |
| Flex-SnakeSA | Snake melón (CUM 353) 4 | melo | Flexuosus | Saudi Arab. |
| La-KroFran | Kroumir 2 | melo | indeterminate landrace | France |
| La-OgenBul | Ogen 3 | melo | indeterminate landrace | Bulgaria |
| La-ZatIta | Zatta 2 | melo | indeterminate landrace | Italy |
| La-CascaPor | Casca de Carvalho 3 | melo | indeterminate landrace, probably Inodorus | Portugal |
| La-ErizoSp | Eriçó mallorquin 3 | melo | indeterminate landrace, probably Inodorus | Spain |
| In-AmCañSp | Caña Dulce 3 | melo | Inodorus | Spain |
| In-LaEscrSp | Escrito Oloroso 3 | melo | Inodorus | Spain |
| In-BBescrSp | Blanco Escrito 3 | melo | Inodorus | Spain |
| In-TeNinvSp2 | Tendral Negro 3 | melo | Inodorus | Spain |
| In-TeSp | Tendral 3 | melo | Inodorus | Spain |
| In-PsPiñSp | Piel de sapo Piñonet 3 | melo | Inodorus | Spain |
| In-TeNinvSp | Negro de Invierno 3 | melo | Inodorus | Spain |
| In-BBredSp | Blanco Redondo 3 | melo | Inodorus | Spain |
| In-RoMoch1Sp | Mochuelo 3 | melo | Inodorus | Spain |
| In-ComunSp | Comun 3 | melo | Inodorus | Spain |
| In-LaBolasSp | Bolas 3 | melo | Inodorus | Spain |
| In-AmAoroSp | Amarillo oro 3 | melo | Inodorus | Spain |
| In-CGBUSA | Casaba Golden Beauty 3 | melo | Inodorus | USA |
| In-CrabPor | Crabranco 2 | melo | Inodorus | Portugal |
| In-HoneyDewUSA | Honeydew green flesh 3 | melo | Inodorus | USA |
| In-TDewUSA | Tam Dew 3 | melo | Inodorus | USA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esteras, C.; Rambla, J.L.; Sánchez, G.; Granell, A.; Picó, M.B. Melon Genetic Resources Characterization for Rind Volatile Profile. Agronomy 2020, 10, 1512. https://doi.org/10.3390/agronomy10101512
Esteras C, Rambla JL, Sánchez G, Granell A, Picó MB. Melon Genetic Resources Characterization for Rind Volatile Profile. Agronomy. 2020; 10(10):1512. https://doi.org/10.3390/agronomy10101512
Chicago/Turabian StyleEsteras, Cristina, Jose Luis Rambla, Gerardo Sánchez, Antonio Granell, and María Belén Picó. 2020. "Melon Genetic Resources Characterization for Rind Volatile Profile" Agronomy 10, no. 10: 1512. https://doi.org/10.3390/agronomy10101512
APA StyleEsteras, C., Rambla, J. L., Sánchez, G., Granell, A., & Picó, M. B. (2020). Melon Genetic Resources Characterization for Rind Volatile Profile. Agronomy, 10(10), 1512. https://doi.org/10.3390/agronomy10101512