Genetic Diversity and Inter-Trait Relationships among Maize Inbreds Containing Genes from Zea diploperennis and Hybrid Performance under Contrasting Environments


 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Development of Genetic Materials
2.2. Field Evaluation
2.2.1. Experimental Locations and Field Layout
2.2.2. Data Collection
2.3. Statistical Analyses
2.4. Molecular Analysis
2.4.1. DNA Extraction and Genotyping with SNP Markers
2.4.2. Data Cleaning and Genetic Diversity Analysis
2.4.3. Population Structure Analysis
3. Results
3.1. Inbred and Hybrid Performance and Reactions under Striga Infestation
3.2. Relative Contributions of Combining Ability Effects
3.3. General Combining Ability Effects
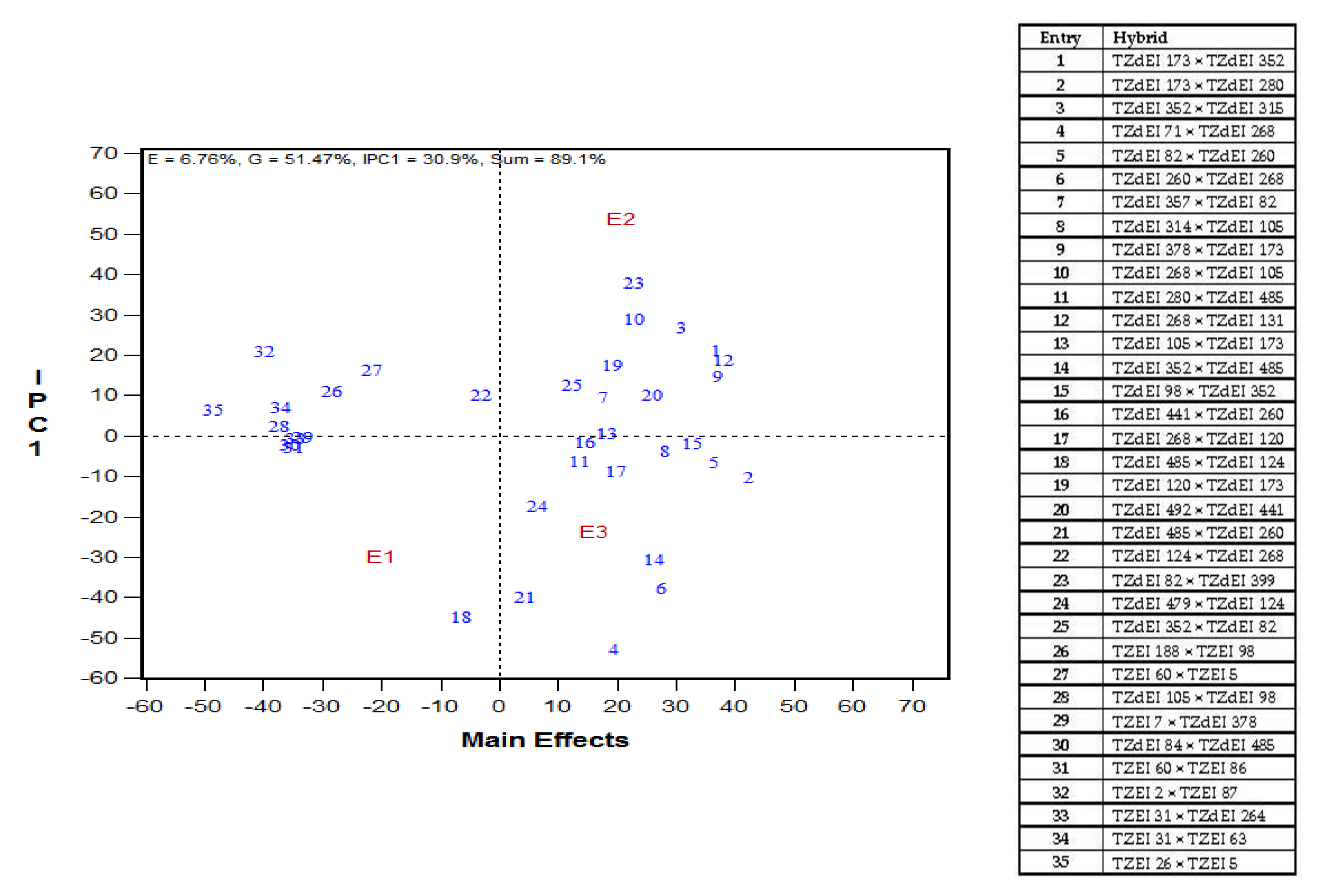
3.4. Relationship between Performance of Parental Inbred Lines and Their Hybrids
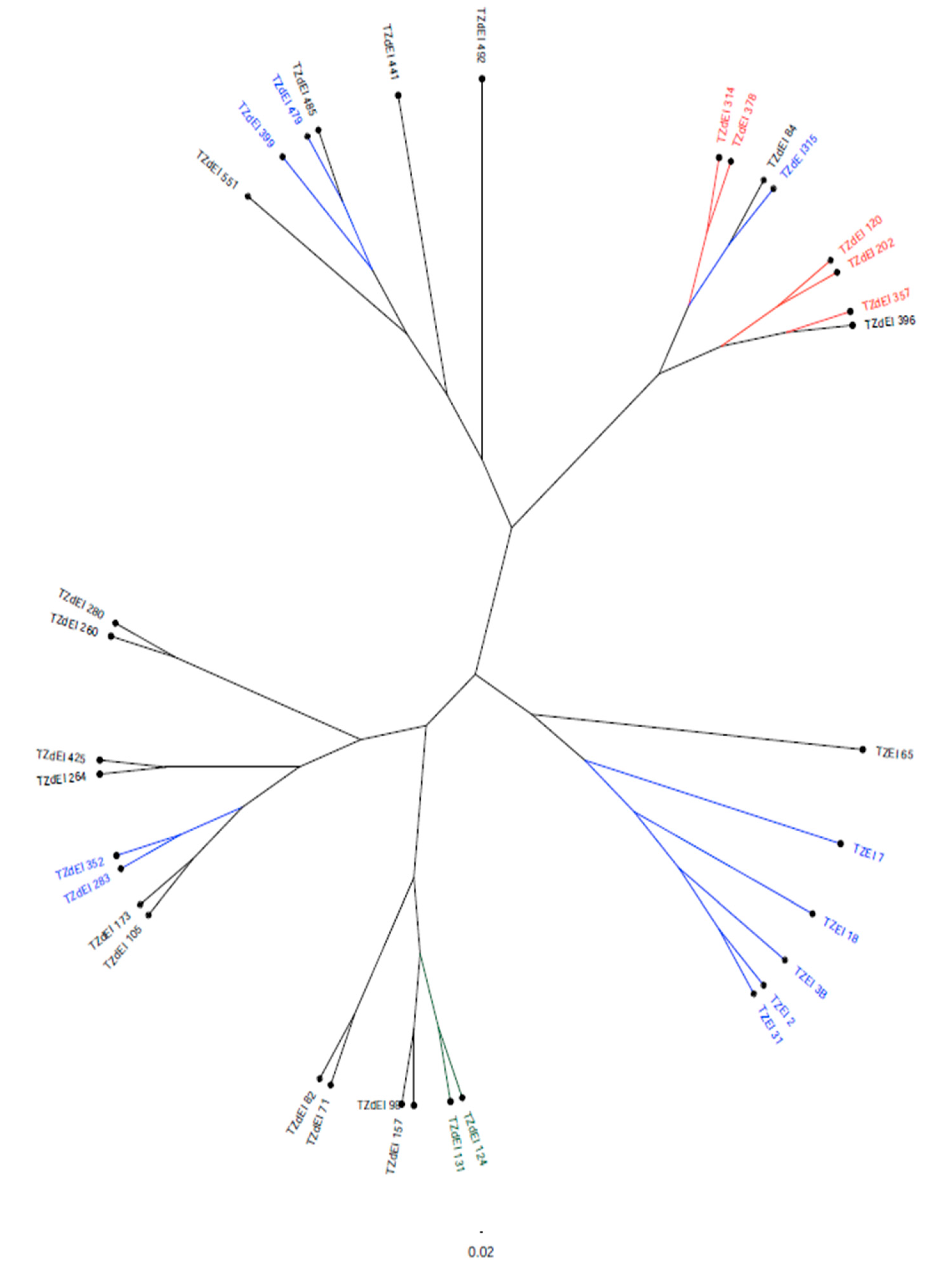
3.5. Genetic Diversity Analysis Using DArT-Seq Markers
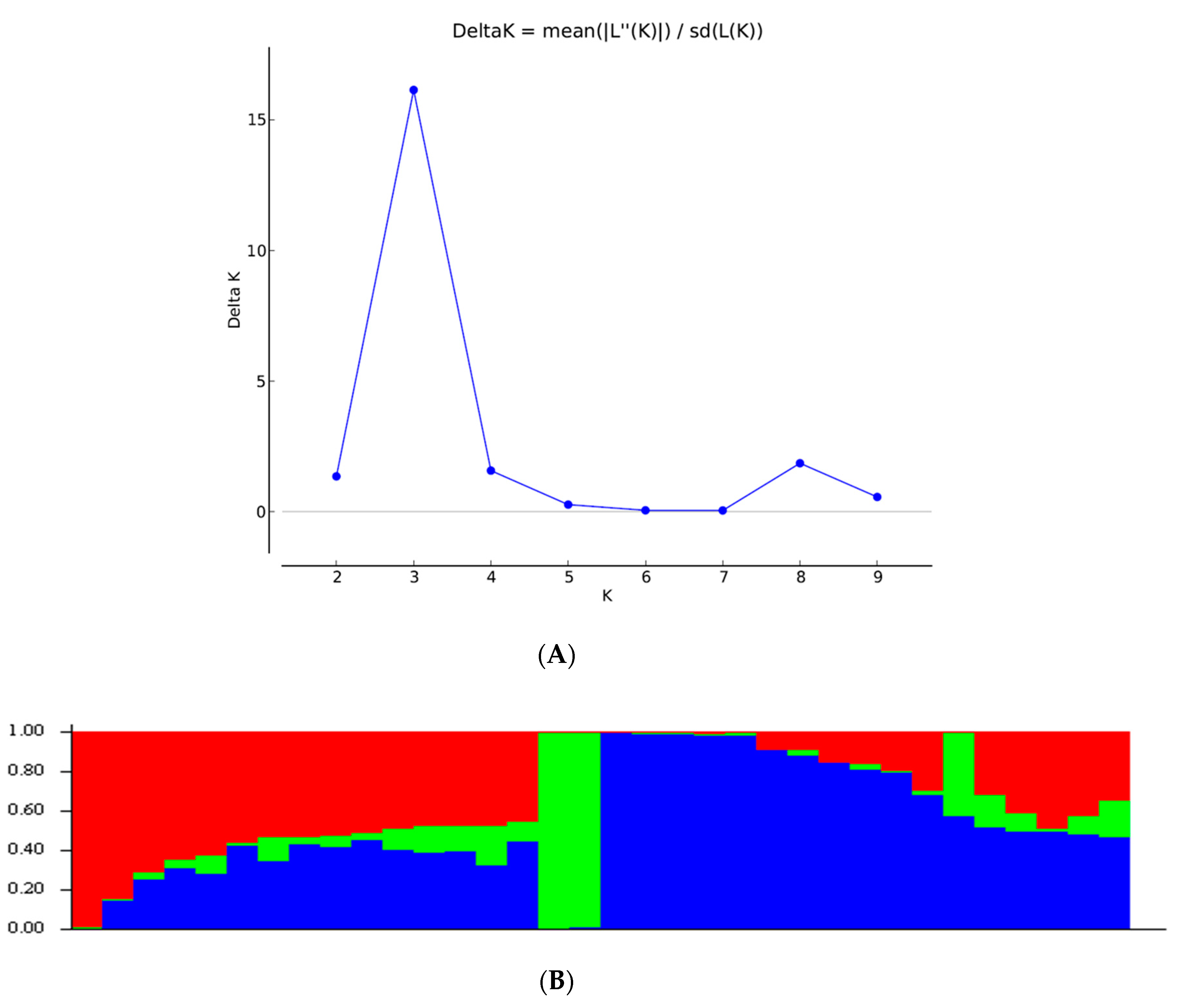
3.6. Population Structure
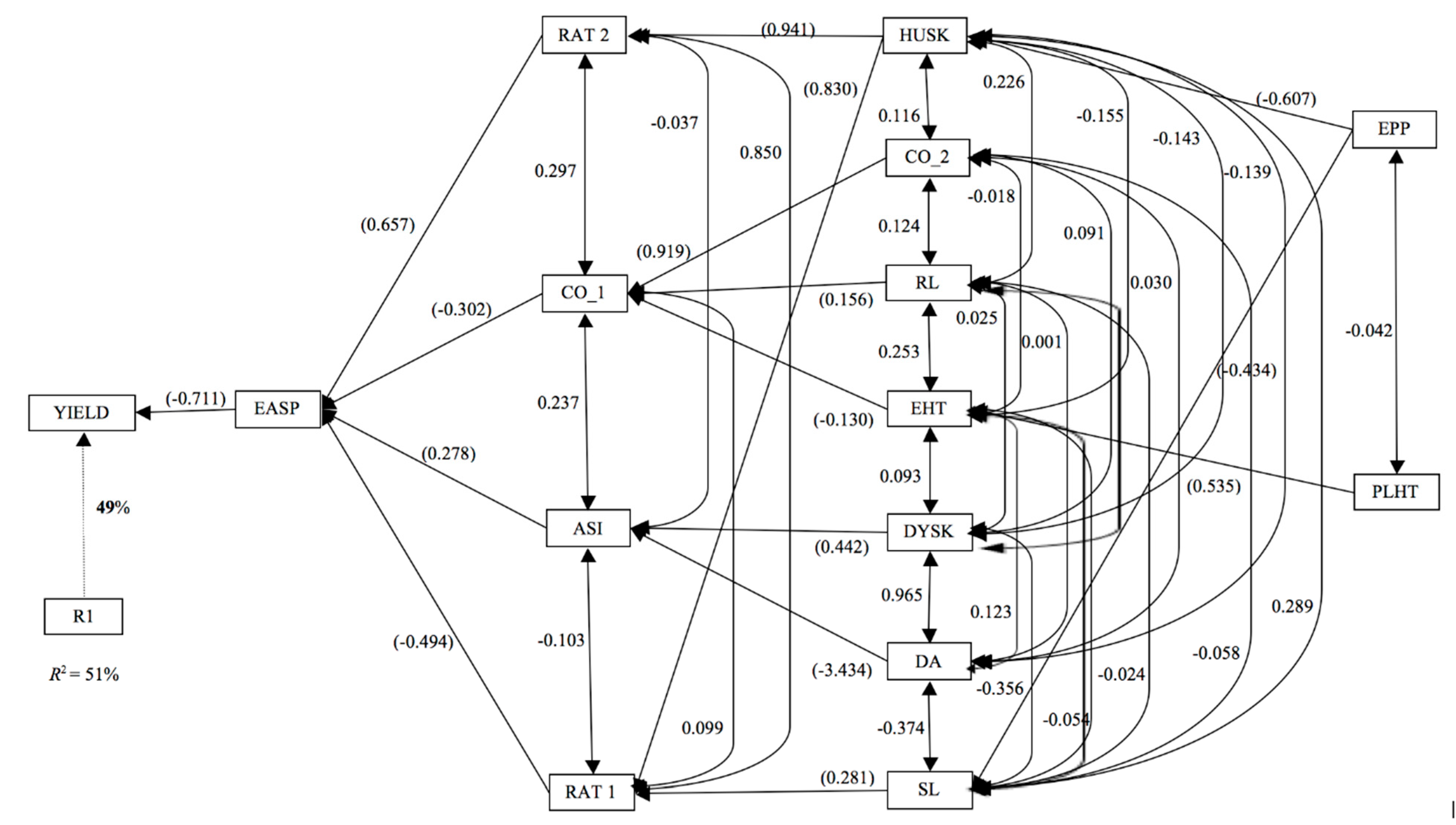
3.7. Relative Importance of Secondary Traits to Grain Yield under Striga Infestation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kling, J.G.; Fajemisin, J.M.; Badu-Apraku, B.; Diallo, A.; Menkir, A.; Melake-Berhan, A. Striga Resistance Breeding in Maize. In Breeding for Striga Resistance in Cereals, Proceedings of a Workshop held at IITA, Ibadan, Nigeria, 18–20 August 1999; Haussmann, B.I.G., Hess, D.E., Koyama, M.L., Grivet, L., Rattunde, H.F.W., Geiger, H.H., Eds.; Margraf Publishers: Weikersheim, Germany, 2000; pp. 103–118. [Google Scholar]
- Menkir, A.; Kling, J.G.; Badu-Apraku, B.; Ibikunle, O. Registration of 26 tropical maize germplasm lines with resistance to Striga hermonthica. Crop Sci. 2006, 46, 1007–1009. [Google Scholar] [CrossRef]
- Amegbor, I.K.; Badu-Apraku, B.; Annor, B. Combining ability and heterotic patterns of extra-early maturing white maize inbreds with genes from Zea diploperennis under multiple environments. Euphytica 2017, 213, 24. [Google Scholar] [CrossRef]
- Dao, A.; Sanou, J.; Mitchell, S.E.; Gracen, V.; Danquah, E.Y. Genetic diversity among INERA maize inbred lines with single nucleotide polymorphism (SNP) markers and their relationship with CIMMYT, IITA, and temperate lines. BMC Genet. 2014, 15, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semagn, K.; Magorokosho, C.; Vivek, B.S.; Makumbi, D.; Beyene, Y.; Mugo, S.; Prasanna, B.M.; Warburton, M.L. Molecular characterization of diverse CIMMYT maize inbred lines from eastern and southern Africa using single nucleotide polymorphic markers. Genomics 2012, 13, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Yan, J.; Guimarães, C.T.; Taba, S.; Hao, Z.; Gao, S.; Chen, S.; Li, J.; Zhang, S.; Vivek, B.S.; et al. Molecular characterization of global maize breeding germplasm based on genome-wide single nucleotide polymorphisms. Theor. Appl. Genet. 2009, 120, 93–115. [Google Scholar] [CrossRef]
- Senior, M.L.; Murphy, J.P.; Goodman, M.M.; Stuber, C.W. Utility of SSRs for determining genetic similarities and relationships in maize using an agarose gel system. Crop Sci. 1998, 38, 1088–1098. [Google Scholar] [CrossRef]
- Yan, J.; Shah, T.; Warburton, M.L.; Buckler, E.S.; McMullen, M.D.; Crouch, J. Genetic characterization and linkage disequilibrium estimation of a global maize collection using SNP markers. PLoS ONE 2009, 4, e8451. [Google Scholar] [CrossRef] [Green Version]
- Van-Inghelandt, D.V.; Melchinger, A.E.; Lebreton, C.; Stich, B. Population structure and genetic diversity in a commercial maize breeding program assessed with SSR and SNP markers. Theor. Appl. Genet. 2010, 120, 1289–1299. [Google Scholar] [CrossRef] [Green Version]
- Dos-Santos, J.P.R.; Pires, L.P.M.; Pires, M.; Vasconcellos, R.C.C.; Pereira, G.S.; Pinho, R.G.V.; Balestre, M. Genomic selection to resistance to Stenocarpella maydis in maize lines using DArTseq markers. BMC Genet. 2016, 17, 86. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, H.; Li, L.; Lan, H.; Ren, Z.; Liu, D.; Wu, L.; Liu, H.; Jaqueth, J.; Li, B.; et al. Characterizing the population structure and genetic diversity of maize breeding germplasm in Southwest China using genome-wide SNP markers. BMC Genom. 2016, 17, 697. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, B.M. Diversity in global maize germplasm: Characterization and utilization. J. Biosci. 2012, 37, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Menkir, A.; Kling, J.G.; Badu-Apraku, B.; Ingelbrecht, I. Molecular marker-based genetic diversity assessment of Striga-resistant maize inbred lines. Theor. Appl. Genet. 2005, 110, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Gebremeskel, S.; Garcia-Oliveira, A.L.; Menkir, A.; Adetimirin, V.O.; Gedil, M. Effectiveness of predictive markers for marker assisted selection of pro-vitamin A carotenoids in medium-late maturing maize (Zea mays L.) inbred lines. J. Cereal. Sci. 2018, 79, 27–34. [Google Scholar] [CrossRef]
- Akaogu, I.C.; Badu-Apraku, B.; Adetimirin, V.O.; VROH-BI, I.; Oyekunle, M.; Akinwale, R.O. Genetic diversity assessment of extra-early maturing yellow maize inbreds and hybrid performance in Striga-infested and Striga-free environments. J. Agric. Sci. 2012, 151, 519–537. [Google Scholar] [CrossRef]
- Mohammadi, S.A.; Prasanna, B.M.; Singh, N.N. Sequential path model for determining interrelationships among grain yield and related characters in maize. Crop Sci. 2003, 43, 1690–1697. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Talabi, A.O.; Oyekunle, M.; Akaogu, I.C.; Akinwale, R.O.; Annor, B.; Melaku, G.; Fasanmade, Y.; Aderounmu, M. Gene action and heterotic groups of early white quality protein maize inbreds under multiple stress environments. Crop Sci. 2015, 56, 183–199. [Google Scholar] [CrossRef]
- Lane, J.A.; Child, D.V.; Moore, T.H.M.; Arnold, G.M.; Bailey, J.A. Phenotypic characterization of resistance in Zea diploperennis to Striga hermonthica. Maydica 1997, 42, 45–51. [Google Scholar]
- Comstock, R.E.; Robinson, H.F. The components of genetic variance in populations of bi-parental progenies and their use in estimating the average degree of dominance. Biometrics 1948, 4, 254–266. [Google Scholar] [CrossRef]
- Kim, S.K. Breeding maize for Striga tolerance and the development of a field technique. In Combating Striga in Africa, Proceedings of the International Workshop organized by IITA, ICRISAT and IDRC at, IITA, Ibadan, Nigeria, 22-24 August, 1988; Kim, S.K., Ed.; International Institute of Tropical Agriculture: Ibadan, Nigeria, 1991; pp. 96–108. [Google Scholar]
- Badu-Apraku, B.; Akinwale, R.O. Cultivar evaluation and trait analysis of tropical early maturing maize under Striga-infested and Striga-free environments. Field Crops Res. 2011, 121, 186–194. [Google Scholar] [CrossRef]
- DeVries, J. The Inheritance of Striga Reactions in Maize. In Breeding for Striga Resistance in Cereals; Margraf: Weikersheim, Germany, 2000; pp. 73–84. [Google Scholar]
- SAS Institute Inc. Base SAS 9.3 Procedures Guide; SAS Institute Inc.: Cary, NC, USA, 2011. [Google Scholar]
- Hallauer, A.R.; Miranda, J.B. Quantitative Genetics in Maize Breeding, 2nd ed.; Iowa State University Press: Ames, IA, USA, 1988. [Google Scholar]
- Badu-Apraku, B.; Lum, A.F.; Akinwale, R.O.; Oyekunle, M. Biplot analysis of diallel crosses of early maturing tropical yellow maize inbreds in stress and nonstress environments. Crop Sci. 2011, 51, 173–188. [Google Scholar] [CrossRef]
- Zobel, R.W.; Wright, M.J.; Gauch, H.G. Statistical analysis of a yield trial. Agron. J. 1988, 80, 388–393. [Google Scholar] [CrossRef]
- Gauch, H.G.; Zobel, R.W. Predictive and postdictive success of statistical analyses of 14 yield trials. Theor. App. Genet. 1988, 76, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Crossa, J. Statistical analyses of multilocation trials. Adv. Agron. 1990, 44, 55–85. [Google Scholar]
- Baker, R.J. Issues in diallel analysis. Crop Sci. 1978, 18, 533–536. [Google Scholar] [CrossRef]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Longman Technical: Harlow, UK, 1996. [Google Scholar]
- Badu-Apraku, B.; Akinwale, R.O.; Fakorede, M.A.B.; Oyekunle, M.; Franco, J. Relative changes in genetic variability and correlations in an early-maturing maize population during recurrent selection. Theor. App. Genet. 2012, 125, 1289–1301. [Google Scholar] [CrossRef] [PubMed]
- Badu-Apraku, B.; Akinwale, R.O.; Oyekunle, M. Efficiency of secondary traits in selecting for improved grain yield in extra-early maize under Striga-infested and Striga-free environments. Plant Breed. 2014, 133, 373–380. [Google Scholar] [CrossRef]
- Talabi, A.O.; Badu-Apraku, B.; Fakorede, M.A.B. Genetic variances and relationship among traits of an early-maturing maize population under drought-stress and low-N environments. Crop Sci. 2016, 57, 681–692. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Liu, K.J.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin Software. Available online: http://darwin.cirad.fr/2006 (accessed on 27 May 2020).
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, D.A. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Yang, X.; Xu, Y.; Shah, T.; Li, H.; Han, Z.; Li, J.; Yan, J. Comparison of SSRs and SNPs in assessment of genetic relatedness in maize. Genetica 2011, 139, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Menkir, A.; Kamara, A.Y.; Akanvou, L.; Chabi, Y. Response of early maturing maize to multiple stresses in the Guinea savanna of West and Central Africa. J. Genet. Breed. 2004, 58, 119–130. [Google Scholar]
- Ifie, B.E.; Badu-Apraku, B.; Gracen, V.; Danquah, E.Y. Genetic analysis of grain yield of IITA and CIMMYT early maturing maize inbreds under Striga-infested and low-soil nitrogen environments. Crop Sci. 2015, 55, 610–623. [Google Scholar] [CrossRef]
- Adetimirin, V.O.; Aken’Ova, M.E.; Kim, S.K. Effects of Striga hermonthica on yield components in maize. J. Agric. Sci. 2000, 135, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Adetimirin, V.O.; The, C.; Dossou, R. Yield losses in maize due to Striga hermonthica in West and Central Africa. Int. J. Pest Manag. 2002, 48, 211–217. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Menkir, A.; Ajala, S.O.; Akinwale, R.O.; Oyekunle, M.; Obeng-Antwi, K. Performance of tropical early maturing maize cultivars in multiple stress environments. Can. J. Plant Sci. 2010, 90, 831–852. [Google Scholar] [CrossRef] [Green Version]
- Gethi, J.G.; Smith, M.E. Genetic responses of single crosses of maize to Striga hermonthica (Del.) Benth. and Striga asiatica (L.) kuntze. Crop Sci. 2004, 44, 2068–2077. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Lum, A.F. Evaluation of experimental varieties from recurrent selection for Striga resistance in two extra-early maize populations in the savannas of West and Central Africa. Exp. Agric. 2007, 43, 183–200. [Google Scholar] [CrossRef]
- Kim, S.K. Genetics of maize tolerance to Striga hermonthica. Crop Sci. 1994, 34, 900–907. [Google Scholar] [CrossRef]
- Akanvou, L.; Doku, E.V.; Kling, J.G. Estimates of genetic variances and interrelationships of traits associated with Striga resistance in maize. Afr. Crop Sci. J. 1997, 5, 1–8. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Menkir, A.; & Lum, A.F. Genetic variability for grain yield and components in an early tropical yellow maize population under Striga hermonthica infestation. Crop Improv. 2007, 20, 107–122. [Google Scholar] [CrossRef]
- Laborda, P.R.; Oliveira, K.M.; Garcia, A.A.F.; Paterniani, M.E.A.; deSouza, A.P. Tropical maize germplasm: What can we say about its genetic diversity in the light of molecular markers. Theor. Appl. Genet. 2005, 111, 1288–1299. [Google Scholar] [CrossRef]
- Xia, X.C.; Reif, J.C.; Melchinger, A.E.; Frisch, M.; Hoisington, D.A.; Beck, D.; Pixley, K.; Warburton, M.L. Genetic diversity among CIMMYT maize inbred lines investigated with SSR markers: II. Subtropical, tropical midaltitude, and highland maize inbred lines and their relationships with elite U.S. and European maize. Crop Sci. 2005, 45, 2573–2582. [Google Scholar] [CrossRef]
- Hamblin, M.T.; Warburton, M.L.; Buckler, E.S. Empirical comparison of simple sequence repeats and single nucleotide polymorphisms in assessment of maize diversity and relatedness. PLoS ONE 2007, 2, 1367. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Yang, X.; Shah, T.; Sánchez-Villeda, H.; Li, J.; Warburton, M.; Zhou, Y.; Crouch, J.H.; Xu, Y. High-throughput SNP genotyping with the Golden Gate assay in maize. Mol. Breed. 2010, 25, 441–451. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Degree of Freedom | Grain Yield (kg ha−1) | Days to Silking | Anthesis-Silking Interval | Ear Aspect (Scale 1–9) | Ears per Plant | Striga Damage Rating (Scale 1–9) | No. of Emerged Striga Plants | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Striga Infestation | 8 WAP | 10 WAP | 8 WAP | 10 WAP | ||||||
| Environment (E) | 2 | 6,641,695 ** | 750.56 ** | 5.08 ** | 0.31ns | 0.14 ** | 3.49 ** | 5.91 ** | 9.03 ** | 12.10 ** |
| Genotype (G) | 35 | 998,643 ** | 20.85 ** | 1.76 * | 1.24 ** | 0.07 ** | 1.22 ** | 2.18 ** | 1.43 ** | 1.80 ** |
| BLOCK (E–REP) | 30 | 290,682ns | 5.25ns | 1.30ns | 0.37ns | 0.03ns | 0.40ns | 0.35ns | 0.45ns | 0.42ns |
| Replication/(E) | 3 | 360,056ns | 15.48 ** | 7.17 ** | 1.99 ** | 0.11 ** | 0.22ns | 0.70ns | 1.17ns | 0.46ns |
| G–E | 67 | 310,475ns | 5.62 * | 1.81 * | 0.77 ** | 0.04ns | 0.66 ** | 0.84 ** | 0.63ns | 0.62 * |
| Error | 72 | 221,604 | 3.43 | 1.13 | 0.32 | 0.03 | 0.33 | 0.32 | 0.5 | 0.39 |
| Heritability | 0.7 | 0.74 | 0.33 | 0.39 | 0.48 | 0.47 | 0.63 | 0.33 | 0.51 | |
| HYBRID | ||||||||||
| Environment (E) | 2 | 87,669,271.9 ** | 2769.24 ** | 45.18 ** | 4.33 ** | 1.46 ** | 10.56 ** | 2.82 * | 62.30 ** | 22.92 ** |
| SET | 5 | 3,590,873.0 ** | 83.89 ** | 9.80 ** | 2.23 ** | 0.08 ** | 1.22ns | 1.89 * | 17.77 ** | 8.95 ** |
| E–SET | 10 | 5,133,848.0 ** | 17.07 ** | 1.03ns | 1.39 ** | 0.03ns | 2.26 ** | 1.26 * | 1.61 ** | 1.74 ** |
| HYBRID (G) | 155 | 2,895,635.2 ** | 18.79 ** | 3.25 ** | 1.68 ** | 0.04 ** | 3.08 ** | 3.19 ** | 2.56 ** | 1.65 ** |
| GCAm/SET | 24 | 4,989,075.8 ** | 23.03 ** | 5.52 ** | 2.13 ** | 0.04 ** | 5.12 ** | 5.02 ** | 3.88 ** | 2..42 ** |
| GCAf/SET | 24 | 4,731,992.5 ** | 33.47 ** | 4.40 ** | 3.80 ** | 0.06 ** | 5.69 ** | 6.66 ** | 3.36 ** | 2.52 ** |
| SCA/SET | 96 | 1,320,972.3 ** | 7.78 ** | 1.60ns | 0.82 ** | 0.02ns | 1.35 ** | 1.06 ** | 1.10 ** | 0.75 ** |
| G–E | 310 | 1,951,558.3 ** | 7.01 ** | 2.18 ** | 0.81 ** | 0.03 ** | 1.10 ** | 1.07 ** | 0.76 ** | 0.64 ** |
| E–GCAm/SET | 48 | 2,729,848.9 ** | 7.10 ** | 2.56 ** | 0.86 ** | 0.04 ** | 0.97ns | 1.02 ** | 0.90 ** | 0.85 ** |
| E–GCAf/SET | 48 | 2,374,399.6 ** | 7.85 ** | 2.66 ** | 1.01 ** | 0.03 ** | 1.26 ** | 1.66 ** | 1.10 ** | 0.99 ** |
| E–CA/SET | 192 | 1,404,975.6 * | 5.63 ** | 1.87ns | 0.67 ** | 0.02ns | 0.93 * | 0.87 * | 0.58 ** | 0.45 ** |
| Error | 359 | 622,139 | 3.67 | 1.6 | 0.43 | 0.02 | 0.72 | 0.67 | 0.43 | 0.33 |
| Heritability | 0.3 | 0.64 | 0.6 | 0.54 | 0.38 | 0.65 | 0.67 | 0.67 | 0.64 | |
| Non-Infested | HC | RL | PASP (scale 1–9) | |||||||
| Environment (E) | 3 | 263,742,713.0 ** | 157.20 ** | 9.51 ** | 116.51 ** | 0.34 ** | 227.90 ** | 331.83 * | 177.54 ** | |
| SET | 5 | 18,872,443.0 ** | 98.40 ** | 1.33 ** | 2.40 ** | 0.02ns | 1.91 ** | 9.94 * | 12.91 ** | |
| E–SET | 15 | 4,791,105.0 ** | 13.82 ** | 0.74 ** | 2.10 ** | 0.01ns | 1.08 ** | 3.69 ** | 1.38 ** | |
| HYBRID (G) | 155 | 6,541,812.0 ** | 14.56 ** | 0.79 ** | 1.45 ** | 0.02 ** | 0.61 ** | 2.23 ** | 2.36 ** | |
| GCAm/SET | 24 | 10,693,735.0 ** | 21.53 ** | 1.42 ** | 1.83 ** | 0.03 ** | 0.59 ** | 1.89 * | 3.44 ** | |
| GCAf/SET | 24 | 11,581,716.0 ** | 22.55 ** | 1.14 ** | 1.89 ** | 0.03 ** | 1.31 ** | 2.98 ** | 3.16 ** | |
| SCA/SET | 96 | 3,032,926.0 ** | 6.10 ** | 0.47ns | 0.98 ** | 0.02 ** | 0.36 ** | 1.55 * | 1.06 ** | |
| G*E | 465 | 1,664,380.0 ** | 3.54 ** | 0.45 ** | 0.68 ** | 0.01 * | 0.40 ** | 1.58 ** | 0.77 ** | |
| E*GCAm/SET | 72 | 2,141,306.0 ** | 3.86 ** | 0.68 ** | 0.92 ** | 0.02 ** | 0.51 ** | 1.64 ** | 0.78 ** | |
| E* GCAf/SET | 72 | 2,410,645.0 ** | 4.31 ** | 0.42ns | 0.89 ** | 0.02 ** | 0.58 ** | 1.68 ** | 0.77 ** | |
| E*SCA/SET | 288 | 1,108,390.0 * | 2.67 ** | 0.39ns | 0.45 ** | 0.01ns | 0.30 * | 1.36ns | 0.73 ** | |
| Error | 480 | 859,535 | 1.85 | 0.37 | 0.3 | 0.01 | 0.24 | 1.18 | 0.42 | |
| Heritability | 0.76 | 0.76 | 0.54 | 0.44 | 0.47 | 0.34 | 0.64 | |||
| Genotypes | Grain Yield | ASI | EPP | EASP | SDR | NESP | Selection Index | ||
|---|---|---|---|---|---|---|---|---|---|
| 8 WAP | 10 WAP | 8 WAP | 10 WAP | ||||||
| TZEI 18 | 1778 | 2.33 | 0.74 | 4.17 | 4.33 | 4.83 | 2.42 | 3.1 | −2.48 |
| TZdEI 71 | 2009 | 1.67 | 0.64 | 5.67 | 4.67 | 5.83 | 1.24 | 1.81 | −2.43 |
| TZdEI 82 | 1362 | 0.6 | 0.83 | 5 | 4.4 | 5 | 0.14 | 0.55 | 0.31 |
| TZdEI 84 | 909 | 1.83 | 0.75 | 5.67 | 5 | 5.5 | 0.41 | 1.2 | −5.33 |
| TZdEI 98 | 2414 | 0.5 | 0.86 | 4 | 3.83 | 4.17 | 1.3 | 1.96 | 5.51 |
| TZdEI 105 | 1425 | 1.83 | 0.85 | 4.83 | 4.17 | 4.5 | 1.42 | 1.9 | −0.26 |
| TZdEI 120 | 1332 | 0.83 | 0.75 | 4.5 | 4.67 | 5.17 | 1.04 | 1.6 | −3.07 |
| TZdEI 124 | 1971 | 0.67 | 0.94 | 4.67 | 3.33 | 4 | 1.27 | 1.6 | 5.93 |
| TZdEI 131 | 1921 | 1.5 | 0.93 | 4.5 | 3.5 | 4.67 | 1.53 | 2.38 | 3.38 |
| TZdEI 157 | 1446 | 0.67 | 0.92 | 4.33 | 4 | 4.67 | 1.54 | 2.23 | 0.18 |
| TZdEI 173 | 1273 | 1.67 | 1.03 | 4.67 | 4 | 4.33 | 1.17 | 1.92 | 1.5 |
| TZdEI 202 | 1085 | 1.67 | 0.75 | 4.67 | 4.5 | 5 | 1.01 | 1.65 | −3.62 |
| TZdEI 260 | 1406 | 0.83 | 0.95 | 4.83 | 4.17 | 4.83 | 0.46 | 1.12 | 1.61 |
| TZdEI 264 | 1255 | 1.67 | 0.91 | 5.17 | 4 | 5.67 | 2.04 | 2.75 | −3.21 |
| TZdEI 268 | 1329 | 0.33 | 0.94 | 5.17 | 4 | 4.83 | 1.49 | 2.15 | −0.27 |
| TZdEI 280 | 1815 | 1.83 | 0.85 | 4.83 | 4.17 | 5 | 1.79 | 2.12 | 0.16 |
| TZdEI 283 | 2879 | 1.5 | 0.98 | 3.5 | 3.33 | 3.67 | 1.6 | 2.56 | 9.78 |
| TZdEI 314 | 1239 | 1 | 0.84 | 4.83 | 4.33 | 5.33 | 2.24 | 2.92 | −4.41 |
| TZdEI 315 | 1181 | 1 | 0.9 | 4.83 | 4.5 | 5.33 | 1.28 | 2.01 | −2.84 |
| TZdEI 352 | 1628 | 2.5 | 1.15 | 4.5 | 3 | 3.5 | 1.33 | 1.94 | 7.49 |
| TZdEI 357 | 1962 | 1.67 | 0.95 | 4.5 | 3.67 | 3.83 | 1.43 | 2.31 | 4.81 |
| TZdEI 378 | 1593 | 1.17 | 0.99 | 4.5 | 4.33 | 4.83 | 1.75 | 2.41 | 0.19 |
| TZdEI 396 | 1791 | 0.83 | 0.99 | 4.67 | 4.17 | 5 | 1.81 | 2.61 | 0.92 |
| TZdEI 399 | 1541 | 2 | 0.86 | 4.67 | 3.67 | 4.67 | 2.17 | 2.74 | −0.26 |
| TZdEI 425 | 1011 | 1.5 | 0.92 | 5.5 | 4.25 | 5.5 | 1.6 | 2.4 | −3.81 |
| TZdEI 441 | 1319 | 2 | 0.98 | 4.83 | 3.5 | 4.17 | 1.19 | 1.8 | 2.73 |
| TZdEI 479 | 1607 | 1.33 | 0.95 | 4.67 | 4 | 4.67 | 1.24 | 2.04 | 1.6 |
| TZdEI 485 | 1028 | 1.17 | 0.84 | 5 | 3.83 | 5 | 0.97 | 1.01 | −1.07 |
| TZdEI 492 | 1706 | 1.17 | 0.84 | 4.5 | 4.33 | 5.17 | 2.1 | 2.69 | −1.76 |
| TZdEI 551 | 945 | 2.5 | 0.69 | 5.83 | 4.33 | 5.5 | 1.66 | 2.26 | −6.32 |
| TZEI 2 | 2291 | 1.33 | 0.93 | 4.17 | 4.17 | 4.5 | 1.12 | 1.93 | 4.61 |
| TZEI 3B | 1567 | 1.17 | 0.92 | 4.67 | 4.5 | 5.17 | 1.09 | 1.68 | −0.19 |
| TZEI 26 | 948 | 1.17 | 0.73 | 5.33 | 5.17 | 6.33 | 2.75 | 2.99 | −10.52 |
| TZEI 65 | 1789 | 1 | 0.93 | 4.83 | 3.5 | 3.67 | 0.68 | 1.48 | 5.36 |
| Overall mean | 1548 | 1.36 | 0.88 | 4.77 | 4.09 | 4.83 | 1.42 | 2.09 | |
| Hybrids | Grain Yield (kg ha−1) | Days to 50% Silking | SDR ‡ (WAP) | NESP (WAP) | Ear Aspect (Scale 1–9) | Ears per Plant | BI | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| STR | OPT | STR | OPT | 8 | 10 | 8 | 10 | STR | OPT | STR | OPT | ||
| TZdEI 173 × TZdEI 352 | 4676 | 4347 | 58 | 53 | 1.67 | 2.33 | 2.64 | 2.79 | 3.67 | 4 | 0.87 | 0.96 | 11.05 |
| TZdEI 173 × TZdEI 280 | 5362 | 6816 | 57 | 51 | 2.67 | 3.50 | 2.80 | 3.16 | 3.83 | 3.8 | 0.97 | 0.96 | 10.82 |
| TZdEI 352 × TZdEI 315 | 4821 | 5969 | 60 | 52 | 2.50 | 3.17 | 3.26 | 3.36 | 4.17 | 4.05 | 1.05 | 0.94 | 10.31 |
| TZdEI 71 × TZdEI 268 | 4432 | 7068 | 57 | 52 | 3.67 | 4.00 | 1.70 | 2.44 | 4.67 | 4.7 | 1.00 | 0.97 | 8.05 |
| TZdEI 82 × TZdEI 260 | 4946 | 6856 | 55 | 50 | 3.33 | 4.67 | 2.24 | 3.04 | 3.83 | 4 | 0.96 | 0.99 | 7.61 |
| TZdEI 260 × TZdEI 268 | 4193 | 6370 | 56 | 50 | 3.67 | 4.67 | 1.19 | 2.13 | 4.67 | 4.4 | 1.04 | 0.99 | 7.59 |
| TZdEI 357 × TZdEI 82 | 4230 | 6246 | 58 | 51 | 3.00 | 3.50 | 2.40 | 3.09 | 4.67 | 4.1 | 0.97 | 0.99 | 7.58 |
| TZdEI 314 × TZdEI 105 | 4385 | 5109 | 58 | 51 | 3.00 | 3.67 | 2.88 | 3.26 | 3.83 | 4.55 | 0.98 | 0.98 | 7.42 |
| TZdEI 378 × TZdEI 173 | 4688 | 6272 | 58 | 52 | 2.50 | 3.50 | 3.38 | 3.74 | 4.33 | 4.05 | 0.90 | 0.97 | 7.34 |
| TZdEI 268 × TZdEI 105 | 4215 | 4971 | 58 | 54 | 2.83 | 3.50 | 2.63 | 3.06 | 3.83 | 4.25 | 0.91 | 0.97 | 6.88 |
| TZdEI 280 × TZdEI 485 | 3510 | 5880 | 62 | 52 | 3.33 | 4.17 | 1.20 | 1.93 | 5.33 | 4.6 | 1.02 | 0.96 | 6.77 |
| TZdEI 268 × TZdEI 131 | 4681 | 5383 | 57 | 51 | 3.00 | 4.00 | 2.86 | 3.35 | 4.17 | 4.5 | 0.87 | 0.93 | 6.41 |
| TZdEI 105 × TZdEI 173 | 3993 | 5448 | 57 | 51 | 2.83 | 3.33 | 2.94 | 3.19 | 4.50 | 4.45 | 0.93 | 0.98 | 6.39 |
| TZdEI 352 × TZdEI 485 | 3814 | 6305 | 60 | 53 | 3.33 | 4.33 | 2.09 | 2.53 | 4.67 | 4.8 | 1.03 | 1.01 | 6.26 |
| TZdEI 98 × TZdEI 352 | 4368 | 5868 | 61 | 54 | 2.67 | 3.67 | 2.81 | 3.17 | 4.17 | 4.3 | 0.83 | 0.97 | 6.15 |
| TZdEI 441 × TZdEI 260 | 3821 | 7033 | 58 | 52 | 2.33 | 4.00 | 2.43 | 3.24 | 4.50 | 3.5 | 0.94 | 1.01 | 6.14 |
| TZdEI 268 × TZdEI 120 | 4023 | 5581 | 58 | 51 | 3.17 | 4.33 | 2.69 | 2.98 | 4.50 | 4.35 | 1.02 | 1.08 | 6.12 |
| TZdEI 485 × TZdEI 124 | 3415 | 5111 | 58 | 51 | 3.17 | 4.17 | 1.80 | 2.23 | 5.17 | 5.3 | 0.99 | 0.91 | 5.66 |
| TZdEI 120 × TZdEI 173 | 4264 | 5388 | 59 | 52 | 3.83 | 4.00 | 2.90 | 3.10 | 4.50 | 4.2 | 0.98 | 0.95 | 5.57 |
| TZdEI 492 × TZdEI 441 | 3963 | 6245 | 62 | 55 | 4.00 | 4.83 | 2.28 | 2.74 | 4.83 | 4.1 | 1.08 | 0.99 | 5.43 |
| TZdEI 485 × TZdEI 260 | 3589 | 6442 | 59 | 51 | 3.83 | 5.00 | 1.33 | 2.23 | 4.83 | 4.9 | 1.07 | 0.93 | 5.39 |
| TZdEI 124 × TZdEI 268 | 3440 | 3967 | 59 | 53 | 3.60 | 4.00 | 2.83 | 3.12 | 4.20 | 4.55 | 1.13 | 0.95 | 5.36 |
| TZdEI 82 × TZdEI 399 | 3745 | 5910 | 57 | 51 | 2.67 | 3.83 | 2.78 | 3.31 | 4.33 | 4.15 | 0.92 | 0.98 | 5.13 |
| TZdEI 479 × TZdEI 124 | 3482 | 6455 | 58 | 51 | 3.00 | 4.83 | 1.67 | 2.95 | 4.67 | 4.1 | 1.03 | 1.00 | 5.05 |
| TZdEI 352 × TZdEI 82 | 4215 | 5881 | 61 | 52 | 3.17 | 4.00 | 3.06 | 3.31 | 4.83 | 4.05 | 0.88 | 1.01 | 4.90 |
| Check 2 − TZEI 188 × TZEI 98 | 2681 | 5605 | 59 | 52 | 4.17 | 5.50 | 2.80 | 3.41 | 5.50 | 4.55 | 0.71 | 0.91 | −4.53 |
| Check 3 − TZEI 60 × TZEI 5 | 3143 | 6876 | 64 | 54 | 4.83 | 5.67 | 2.94 | 3.33 | 5.50 | 3.3 | 0.68 | 0.97 | −4.73 |
| TZdEI 105 × TZdEI 98 | 2151 | 4308 | 61 | 53 | 4.67 | 6.00 | 2.93 | 3.31 | 6.33 | 4.85 | 0.73 | 0.84 | −7.09 |
| TZEI 7 × TZdEI 378 | 2033 | 5921 | 62 | 51 | 5.17 | 6.00 | 3.31 | 3.73 | 5.83 | 4.45 | 0.81 | 0.93 | −7.79 |
| TZdEI 84 × TZdEI 485 | 1816 | 5731 | 59 | 52 | 5.67 | 6.33 | 2.25 | 2.71 | 5.83 | 4.8 | 0.75 | 0.98 | −8.52 |
| Check 1 − TZEI 60 × TZEI 86 | 2128 | 5333 | 62 | 53 | 5.00 | 6.17 | 3.01 | 3.58 | 5.67 | 4.65 | 0.67 | 0.98 | −8.76 |
| Check 5 − TZEI 2 × TZEI 87 | 1838 | 4167 | 61 | 52 | 5.17 | 6.17 | 3.24 | 3.64 | 6.00 | 5.175 | 0.74 | 0.91 | −9.23 |
| TZEI 31 × TZdEI 264 | 2108 | 5356 | 62 | 52 | 5.50 | 6.50 | 3.86 | 3.94 | 6.17 | 4.6 | 0.82 | 0.97 | −9.24 |
| Check 4 − TZEI 31 × TZEI 63 | 1961 | 4823 | 61 | 53 | 5.50 | 6.33 | 3.39 | 3.74 | 5.33 | 4.4 | 0.66 | 0.96 | −10.73 |
| Check 6 − TZEI 26 × TZEI 5 | 1134 | 4176 | 63 | 53 | 6.00 | 7.33 | 2.78 | 3.41 | 6.67 | 4.9 | 0.67 | 0.84 | −14.14 |
| Means | 3146 | 5601 | 60 | 52 | 3.97 | 4.91 | 2.82 | 3.29 | 5.12 | 4.45 | 0.89 | 0.97 | |
| Standard Error (SE)± | 343 | 312 | 0.86 | 0.46 | 0.37 | 0.36 | 0.30 | 0.25 | 0.29 | 0.19 | 0.06 | 0.04 | |
| Traits | Striga-Infested | Non-Infested | ||||
|---|---|---|---|---|---|---|
| GCA | SCA | GCA | SCA | |||
| Male | Female | Male | Female | |||
| Grain yield | 33.25 | 31.54 | 35.21 | 31.08 | 33.66 | 35.26 |
| Days to anthesis | 25.8 | 31.05 | 43.15 | 31.09 | 30.17 | 38.74 |
| Days to silking | 26.29 | 38.2 | 35.51 | 31.43 | 32.92 | 35.64 |
| Anthesis-silking interval | 33.81 | 26.92 | 39.27 | 32.16 | 25.77 | 42.07 |
| Plant height | 35.55 | 37.81 | 26.64 | 28.72 | 34.22 | 37.06 |
| Ear height | 33.98 | 44.42 | 21.6 | 31.16 | 36.64 | 32.2 |
| Stalk lodging | 22.94 | 25.68 | 51.38 | 16.14 | 30.86 | 53 |
| Root lodging | - | - | - | 17.06 | 26.88 | 56.06 |
| Husk cover | 30.76 | 31.91 | 37.33 | 17.76 | 39.23 | 43 |
| Ear aspect | 23.12 | 41.31 | 35.58 | 23.9 | 24.79 | 51.32 |
| Plant aspect | - | - | - | 31.71 | 29.2 | 39.09 |
| Ears per plant | 21.11 | 29.26 | 49.63 | 21.03 | 24.85 | 54.13 |
| Striga damage rating at eight WAP | 31.6 | 35.11 | 33.29 | - | - | - |
| Striga damage rating at 10 WAP | 31.5 | 41.79 | 26.71 | - | - | - |
| Number of emerged Striga plants at eight WAP * | 33.35 | 28.88 | 37.77 | - | - | - |
| Number of emerged Striga plants at 10 WAP | 30.42 | 31.74 | 37.84 | - | - | - |
| Inbred Line | Grain Yield | Days to Silking | Striga Damage at 8 WAP | Striga Damage at 10 WAP | Number of Emerged Striga Plants 8 WAP | Number of Emerged Striga Plants 10 WAP | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GCAM | GCAF | GCAM | GCAF | GCAM | GCAF | GCAM | GCAF | GCAM | GCAF | GCAM | GCAF | |
| TZdEI 71 | −443.53 | 197.11 | 0.21 | −1.16 * | 0.55 ** | −0.03 | 0.49 * | −0.07 | −0.37 * | −0.06 | −0.32 * | −0.1 |
| TZdEI 124 | 325.70 | 40.81 | −0.99 * | −0.65 | −0.41 ** | −0.21 | −0.55 ** | −0.17 | 0.02 | 0.59 ** | 0.00 | 0.37 * |
| TZdEI 202 | −711.23 * | −422.78 | 0.78 | 1.12 * | 0.42 ** | 0.60 ** | 0.45 * | 0.39 | 0.00 | −0.20 | −0.12 | −0.26 |
| TZdEI 399 | 43.71 | −20.20 | 1.08 * | 1.14 * | −0.25 | −0.63 ** | −0.31 | −0.51 * | 0.54 ** | 0.21 | 0.45 ** | 0.30 |
| TZdEI 260 | 785.34 * | 205.06 | −1.09 * | −0.53 | −0314 | 0.27 | −0.08 | 0.36 | −0.19 | −0.53 ** | −0.00 | −0.30 |
| TZdEI 268 | 449.57 | 893.59 ** | −0.55 | −0.91 * | −0.28 | −0.71 ** | −0.54 ** | −0.62 ** | −0.67 ** | −0.35 * | −0.56 ** | −0.32 * |
| TZdEI 314 | 46.98 | 10.35 | −0.79 | −0.38 | 0.16 | −0.34 | 003 | −0.42 | 0.20 | 0.11 | 0.09 | −0.13 |
| TZdEI 396 | 212.68 | 77.92 | 0.87 * | 0.35 | −0.46 ** | −0.31 | −0.24 | −0.22 | −0.04 | 0.31 | 0.03 | 0.24 |
| TZEI 7 | −171.96 | −481.62 | −0.23 | 0.79 | 0.24 | 0.49 * | 0.16 | 0.58 * | 0.09 | −0.12 | 0.07 | −0.05 |
| TZEI 31 | −537.28 * | −500.25 * | 0.71 | 0.15 | 0.67 ** | 0.86 ** | 0.59 ** | 0.68 ** | 0.42 * | 0.27 | 0.37 * | 0.25 |
| TZdEI 315 | −18.09 | −152.17 | −1.59 ** | −1.39 ** | 0.13 | 0.25 | 0.21 | −0.08 | 0.99 ** | 0.65 ** | 0.70 ** | 0.49 ** |
| TZdEI 479 | −369.27 | −91.22 | 0.25 | −1.02 | 0.13 | 0.12 | 0.27 | 0.52 * | −0.85 ** | −0.54 ** | −0.53 * | −0.32 * |
| TZdEI 82 | 480.30 | 351.36 | −0.62 | −0.85 | 0.09 | −0.01 | −0.19 | −0.25 | 0.04 | −0.08 | −0.10 | −0.05 |
| TZdEI 485 | −246.67 | −157.88 | 0.68 | 1.35 * | −0.01 | 0.09 | 0.11 | 0.09 | −0.51 ** | −0.63 ** | −0.37 * | −0.57 ** |
| TZdEI 441 | 153.72 | 49.91 | 1.28 * | 1.91 ** | −0.34 * | −0.45 * | −0.39 * | −0.28 | 0.41 * | 0.60 * | 0.31 | 0.45 * |
| TZdEI 352 | 410.03 | 632.85 * | 0.49 | 1.28 * | −0.75 ** | −0.64 ** | −0.59 ** | −0.89 ** | −0.05 | 0.14 | −0.15 | −0.06 |
| TZdEI 84 | −759.90 * | −949.04 ** | −0.91 * | −1.42 ** | 0.45 * | 0.63* | 0.47 * | 1.07 ** | −0.44 * | −0.19 | −0.32 * | −0.22 |
| TZdEI 280 | 358.73 | −152.47 | −0.71 | −0.85 | −0.05 | 0.06 | 0.04 | 0.14 | −0.01 | 0.14 | −0.06 | 0.16 |
| TZdEI 357 | −145.54 | 137.60 | 0.92* | 0.41 | −0.05 | −0.41 * | −0.13 | −0.59 * | 0.25 | 0.16 | 0.22 | 0.39 * |
| TZdEI 492 | 136.67 | 331.06 | 0.22 | 0.58 | 0.41 * | 0.36 * | 0.21 | 0.27 | 0.25 | −0.25 | 0.30 * | −0.27 |
| TZdEI 98 | −91.65 | 181.42 | −0.01 | −0.85 | −0.29 | −0.15 | 0.05 | −0.13 | −0.24 | −0.07 | −0.21 | −0.18 |
| TZdEI 157 | −408.50 | −163.55 | 0.33 | 1.45 ** | 0.24 | −0.19 | 0.21 | 0.11 | 0.46 | 0.24 | 0.42* | 0.28 |
| TZdEI 173 | 996.70 ** | 671.98 * | −1.941 ** | −1.78 ** | −0.83 ** | −0.62 ** | −0.99 ** | −0.76 ** | −0.06 | −0.47 * | −0.12 | −0.38 |
| TZdEI 283 | −221.39 | −233.95 | 0.89* | 0.15 | 0.54 ** | 0.21 | 0.35 | 0.24 | −0.05 | 0.10 | −0.04 | 0.06 |
| TZEI 18 | −275.16 | −455.90 | 0.73 | 1.02 * | 0.34 * | 0.75 ** | 0.38 * | 0.54 * | −0.12 | 0.20 | −0.05 | 0.22 |
| TZdEI 105 | 194.83 | −189.63 | −0.88 * | −0.74 | −034 * | −0.03 | −0.32 * | 0.01 | −0.03 | −0.03 | 0.07 | 0.00 |
| TZdEI 120 | −57.93 | 134.39 | −0.58 | 0.13 * | 0.16 | 0.27 | 0.08 | −0.05 | −0.30 | −0.20 | −0.35 | −0.24 |
| TZdEI 131 | 461.84 | −315.13 | −0.75 | 0.06 | −0.64 ** | 0.07 | −0.55 ** | −0.05 | 0.03 | 0.09 | 0.12 | 0.02 |
| TZdEI 264 | −50.63 | −21.88 | 1.391 ** | 1.49 ** | 0.33 * | 0.01 | 0.41 * | 0.31 | 0.12 | −0.38 | −0.04 | −0.24 |
| TZdEI 378 | −548.11 * | 348.50 | −0.94 | −0.4 | 0.49 ** | −0.33 | 0.38 * | −0.32 | 0.18 | 0.52 * | 0.19 | 0.46 * |
| SE ± | 269.81 | 251..63 | 0.46 | 0.46 | 0.16 | 0.18 | 0.17 | 0.21 | 0.15 | 0.17 | 0.15 | 0.16 |
| Traits | Mid-Parent Heterosis | Better Parent Heterosis |
|---|---|---|
| Striga Infested | Striga Infested | |
| Grain yield | 96.2 | 73.07 |
| Days to anthesis | −2.95 | −4.27 |
| Days to silking | −2.28 | −3.76 |
| Ear aspect | 9.35 | 15.26 |
| Ears per plant | 3.97 | −2.59 |
| Striga damage rating at eight WAP | 1.62 | 8.94 |
| Striga damage rating at 10 WAP | 5.22 | 13.45 |
| Number of emerged Striga plants at eight WAP * | 93.21 | 238.39 |
| Number of emerged Striga plants at 10 WAP | 48.9 | 94.23 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akaogu, I.C.; Badu-Apraku, B.; Gracen, V.; Tongoona, P.; Gedil, M.; Unachukwu, N.; Offei, S.K.; Dzidzienyo, D.K.; Hearne, S.; Garcia-Oliveira, A.L. Genetic Diversity and Inter-Trait Relationships among Maize Inbreds Containing Genes from Zea diploperennis and Hybrid Performance under Contrasting Environments . Agronomy 2020, 10, 1478. https://doi.org/10.3390/agronomy10101478
Akaogu IC, Badu-Apraku B, Gracen V, Tongoona P, Gedil M, Unachukwu N, Offei SK, Dzidzienyo DK, Hearne S, Garcia-Oliveira AL. Genetic Diversity and Inter-Trait Relationships among Maize Inbreds Containing Genes from Zea diploperennis and Hybrid Performance under Contrasting Environments . Agronomy. 2020; 10(10):1478. https://doi.org/10.3390/agronomy10101478
Chicago/Turabian StyleAkaogu, Ijeoma Chinyere, Baffour Badu-Apraku, Vernon Gracen, Pangirayi Tongoona, Melaku Gedil, Nnanna Unachukwu, Samuel Kwame Offei, Daniel Kwadjo Dzidzienyo, Sarah Hearne, and Ana Luisa Garcia-Oliveira. 2020. "Genetic Diversity and Inter-Trait Relationships among Maize Inbreds Containing Genes from Zea diploperennis and Hybrid Performance under Contrasting Environments " Agronomy 10, no. 10: 1478. https://doi.org/10.3390/agronomy10101478
APA StyleAkaogu, I. C., Badu-Apraku, B., Gracen, V., Tongoona, P., Gedil, M., Unachukwu, N., Offei, S. K., Dzidzienyo, D. K., Hearne, S., & Garcia-Oliveira, A. L. (2020). Genetic Diversity and Inter-Trait Relationships among Maize Inbreds Containing Genes from Zea diploperennis and Hybrid Performance under Contrasting Environments . Agronomy, 10(10), 1478. https://doi.org/10.3390/agronomy10101478