Analysis of Grain Yield Differences Among Soybean Cultivars under Maize–Soybean Intercropping


Abstract
1. Introduction
2. Materials and Methods
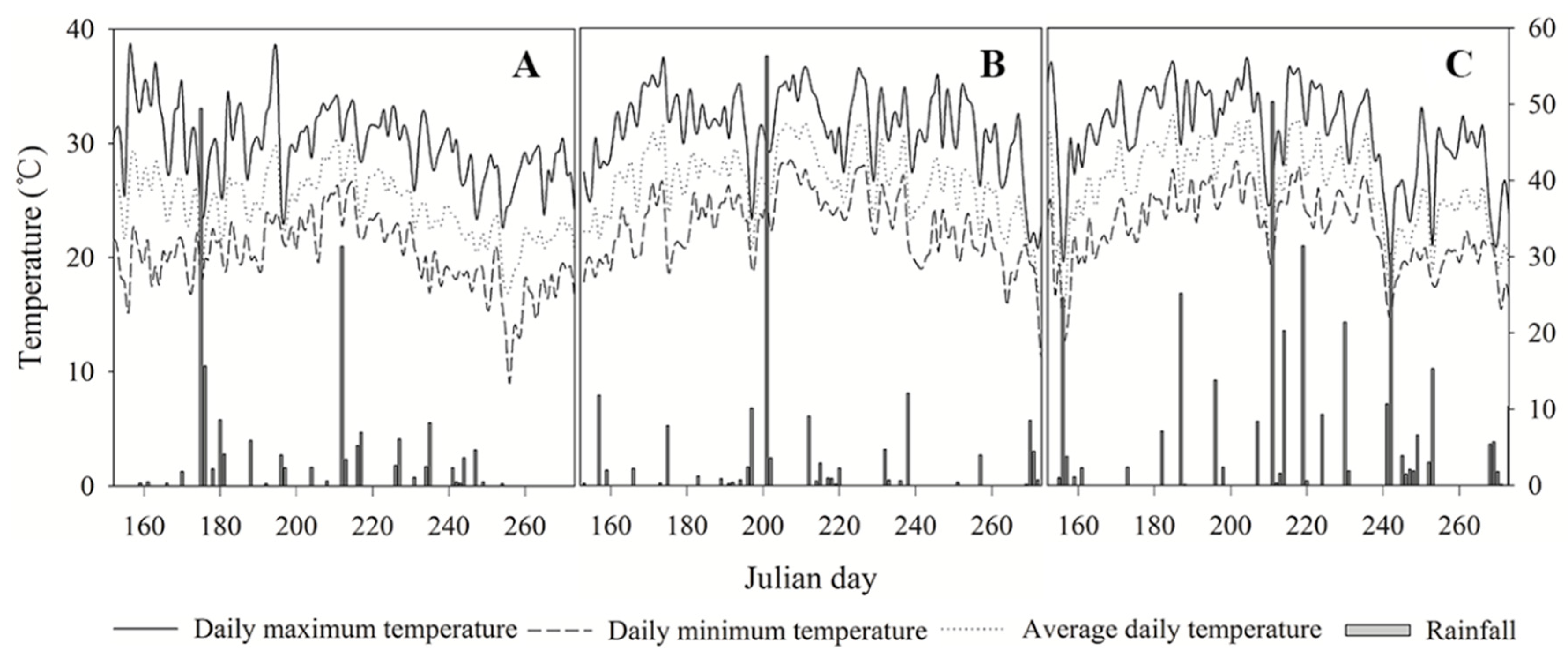
2.1. Experimental Site
2.2. Cultivar
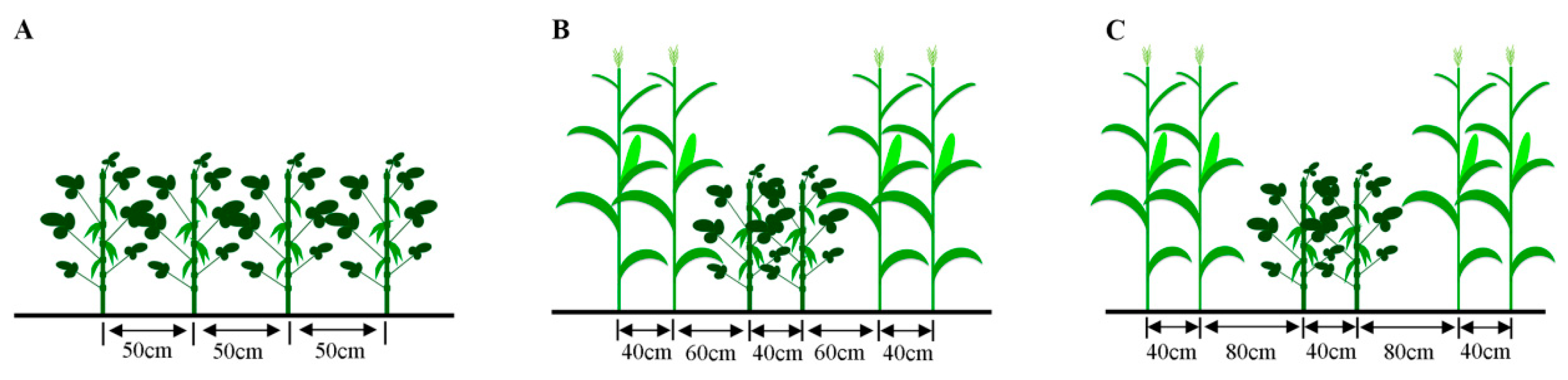
2.3. Experimental Design
2.4. Sampling Methods and Photosynthetic Production Analysis
2.5. Grain Yield and Yield Components
2.6. Reproductive Biomass and Characteristics Determinations
2.7. Statistical Analysis
3. Results
3.1. Yield and Yield Components
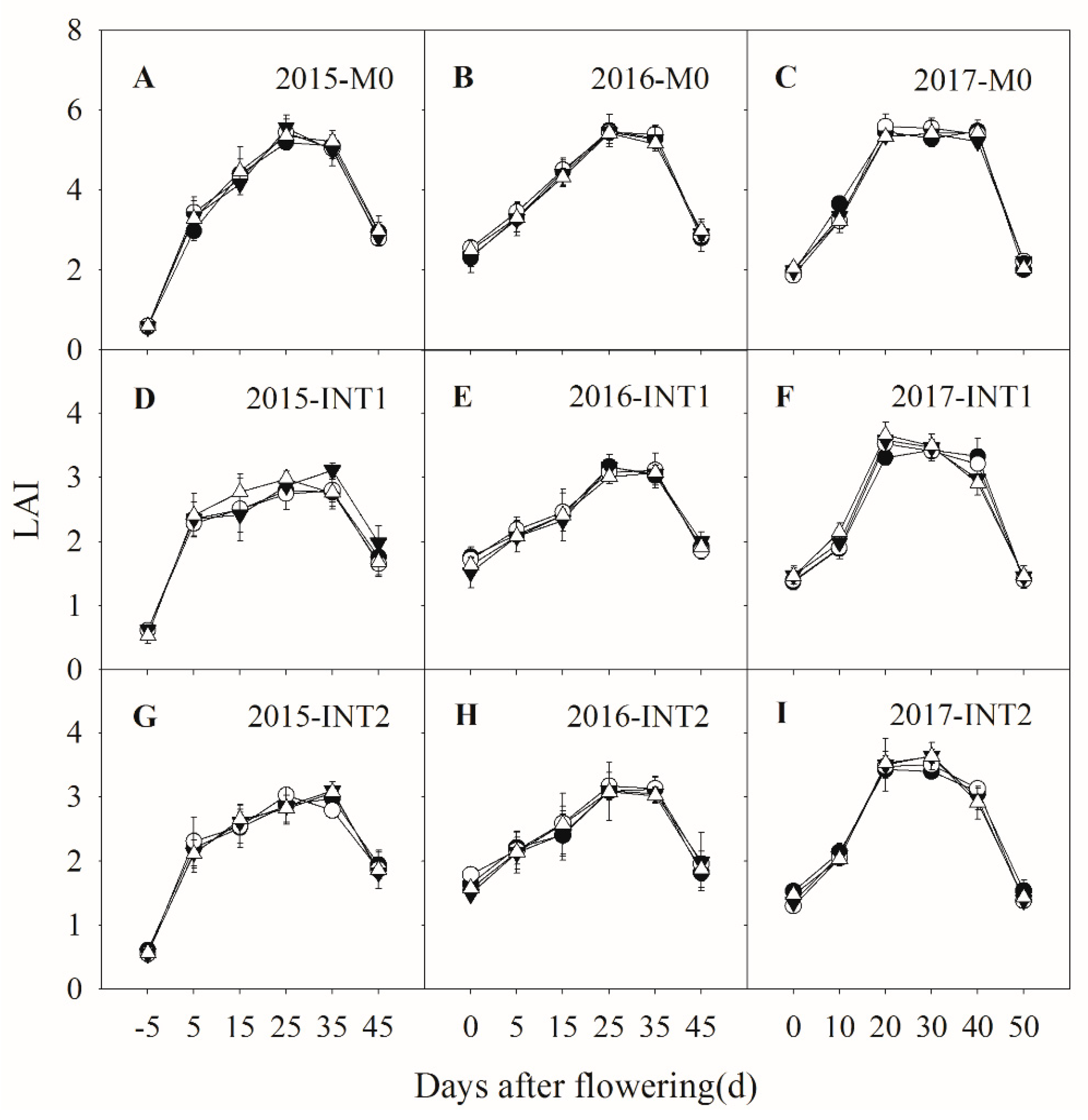
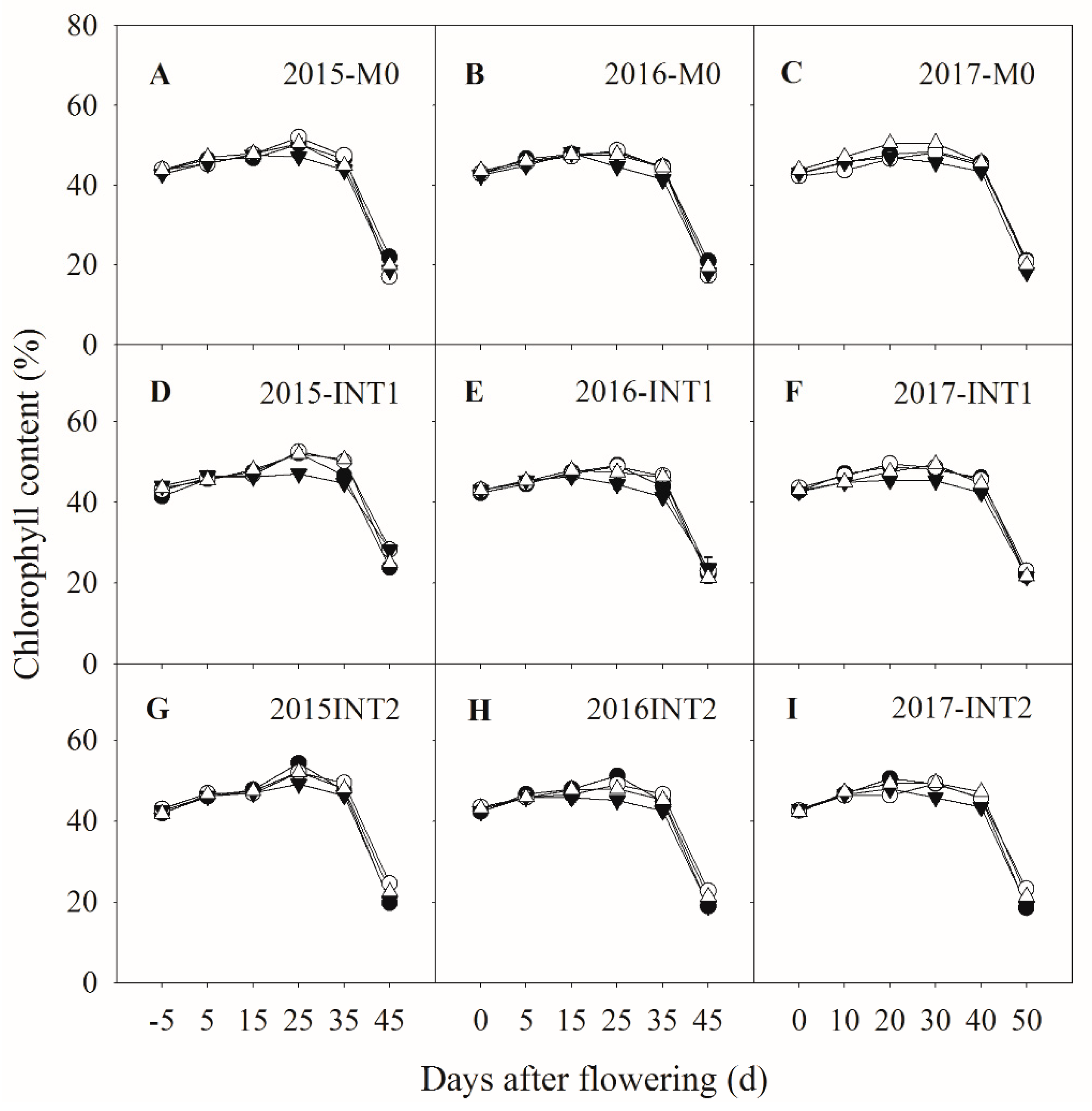
3.2. Photosynthetic Parameters
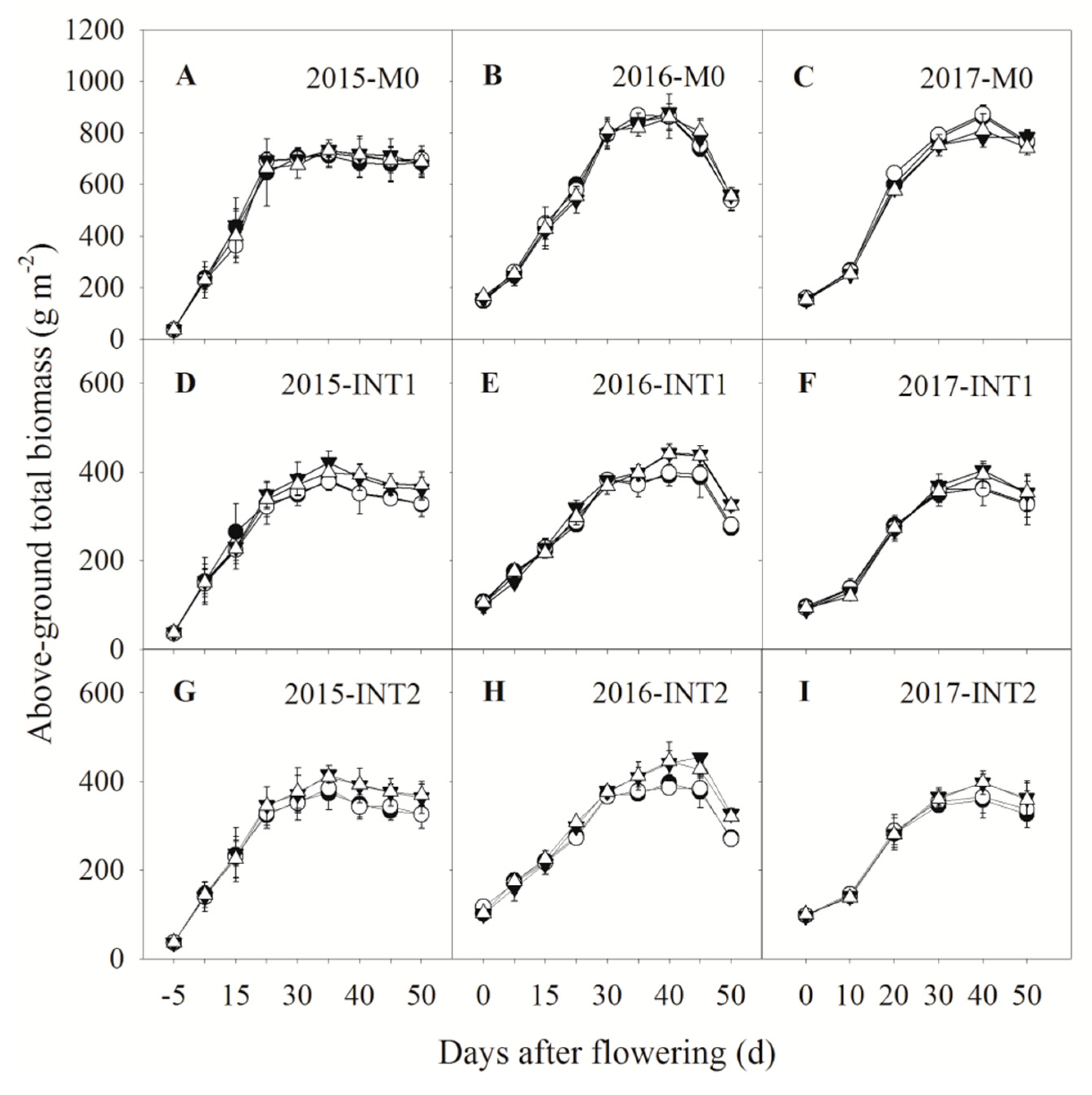
3.3. Total Biomass Accumulation
3.4. Reproductive Biomass and Characteristics
3.5. Correlation Analysis
4. Discussion
4.1. Variation of Yield and Yield Components among Soybean Cultivars in Strip Intercropping
4.2. Variation of Photosynthetic Production and Biomass Accumulation among Soybean Cultivars in Strip Intercropping
4.3. Variation of Seed Number Determinations among Soybean Cultivars in Strip Intercropping
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, F.; Li, L. Using competitive and facilitative interactions in intercropping systems enhances crop productivity and nutrient-use efficiency. Plant Soil 2003, 248, 305–312. [Google Scholar] [CrossRef]
- Lithourgidis, A.; Dordas, C.; Damalas, C.A.; Vlachostergios, D. Annual intercrops: An alternative pathway for sustainable agriculture. Aust. J. Crop Sci. 2011, 5, 396–410. [Google Scholar]
- Regehr, A.; Oelbermann, M.; Videla, C.; Echarte, L. Gross nitrogen mineralization and immobilization in temperate maize-soybean intercrops. Plant Soil 2015, 391, 353–365. [Google Scholar] [CrossRef]
- Carruthers, K.; Prithiviraj, B.; Fe, Q.; Cloutier, D.; Martin, R.; Smith, D. Intercropping corn with soybean, lupin and forages: Yield component responses. Eur. J. Agron. 2000, 12, 103–115. [Google Scholar] [CrossRef]
- Prasad, R.; Brook, R. Effect of varying maize densities on intercropped maize and soybean in Nepal. Exp. Agric. 2005, 41, 365–382. [Google Scholar] [CrossRef]
- Gao, Y.; Duan, A.; Qiu, X.; Sun, J.; Zhang, J.; Liu, H.; Wang, H. Distribution and use efficiency of photosynthetically active radiation in strip intercropping of maize and soybean. Agron. J. 2010, 102, 1149–1157. [Google Scholar] [CrossRef]
- Yang, F.; Liao, D.; Wu, X.; Gao, R.; Fan, Y.; Raza, M.A.; Wang, X.; Yong, T.; Liu, W.; Liu, J. Effect of aboveground and belowground interactions on the intercrop yields in maize-soybean relay intercropping systems. Field Crop. Res. 2017, 203, 16–23. [Google Scholar] [CrossRef]
- Du, J.; Han, T.; Gai, J.; Yong, T.; Sun, X.; Wang, X.; Yang, F.; Liu, J.; Shu, K.; Liu, W. Maize-soybean strip intercropping: Achieved a balance between high productivity and sustainability. J. Integr. Agric. 2018, 17, 747–754. [Google Scholar] [CrossRef]
- Liu, X.; Rahman, T.; Song, C.; Yang, F.; Su, B.; Cui, L.; Bu, W.; Yang, W. Relationships among light distribution, radiation use efficiency and land equivalent ratio in maize-soybean strip intercropping. Field Crop. Res. 2018, 224, 91–101. [Google Scholar] [CrossRef]
- Yu, Y.; Stomph, T.-J.; Makowski, D.; Van Der Werf, W. Temporal niche differentiation increases the land equivalent ratio of annual intercrops: A meta-analysis. Field Crop. Res. 2015, 184, 133–144. [Google Scholar] [CrossRef]
- Andrade, J.F.; Cerrudo, A.; Rizzalli, R.H.; Monzon, J.P. Sunflower–soybean intercrop productivity under different water conditions and sowing managements. Agron. J. 2012, 104, 1049–1055. [Google Scholar] [CrossRef]
- Yang, F.; Wang, X.; Liao, D.; Lu, F.; Gao, R.; Liu, W.; Yong, T.; Wu, X.; Du, J.; Liu, J. Yield response to different planting geometries in maize–soybean relay strip intercropping systems. Agron. J. 2015, 107, 296–304. [Google Scholar] [CrossRef]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef]
- Li, H.; Jiang, D.; Wollenweber, B.; Dai, T.; Cao, W. Effects of shading on morphology, physiology and grain yield of winter wheat. Eur. J. Agron. 2010, 33, 267–275. [Google Scholar] [CrossRef]
- Wang, L.; Deng, F.; Ren, W. Shading tolerance in rice is related to better light harvesting and use efficiency and grain filling rate during grain filling period. Field Crop. Res. 2015, 180, 54–62. [Google Scholar] [CrossRef]
- Su, B.; Song, Y.; Song, C.; Cui, L.; Yong, T.; Yang, W. Growth and photosynthetic responses of soybean seedlings to maize shading in relay intercropping system in Southwest China. Photosynthetica 2014, 52, 332–340. [Google Scholar] [CrossRef]
- Jiang, H.; Egli, D. Shade induced changes in flower and pod number and flower and fruit abscission in soybean. Agron. J. 1993, 85, 221–225. [Google Scholar] [CrossRef]
- Egli, D.B.; Yu, Z.W. Crop growth rate and seeds per unit area in soybean. Crop Sci. 1991, 31, 439–442. [Google Scholar] [CrossRef]
- Kantolic, A.G.; Slafer, G.A. Photoperiod sensitivity after flowering and seed number determination in indeterminate soybean cultivars. Field Crop. Res. 2001, 72, 109–118. [Google Scholar] [CrossRef]
- Kantolic, A.G.; Peralta, G.E.; Slafer, G.A. Seed number responses to extended photoperiod and shading during reproductive stages in indeterminate soybean. Eur. J. Agron. 2013, 51, 91–100. [Google Scholar] [CrossRef]
- Board, J.; Tan, Q. Assimilatory capacity effects on soybean yield components and pod number. Crop Sci. 1995, 35, 846–851. [Google Scholar] [CrossRef]
- Jiang, H.; Egli, D.B. Soybean seed number and crop growth rate during flowering. Agron. J. 1995, 87, 264–267. [Google Scholar] [CrossRef]
- Egli, D.B. Crop growth rate and the establishment of sink size: A comparison of maize and soybean. J. Crop Improv. 2019, 33, 346–362. [Google Scholar] [CrossRef]
- Rotundo, J.L.; Borrás, L.; De Bruin, J.; Pedersen, P. Physiological strategies for seed number determination in soybean: Biomass accumulation, partitioning and seed set efficiency. Field Crop. Res. 2012, 135, 58–66. [Google Scholar] [CrossRef]
- Charles-Edwards, D. On the ordered development of plants 1. An hypothesis. Ann. Bot. 1984, 53, 699–707. [Google Scholar] [CrossRef]
- Vega, C.R.C.; Andrade, F.H.; Sadras, V.O. Reproductive partitioning and seed set efficiency in soybean, sunflower and maize. Field Crop. Res. 2001, 72, 163–175. [Google Scholar] [CrossRef]
- Hiebsch, C.; Tetio-Kagho, F.; Chirembo, A.; Gardner, F. Plant density and soybean maturity in a soybean-maize intercrop. Agron. J. 1995, 87, 965–969. [Google Scholar] [CrossRef]
- Fehr, W.R.; Caviness, C.E. Stages of Soybean Development; Iowa State University. Agricultural and Home Economics Experiment Station: Ames, IA, USA, 1977. [Google Scholar]
- Board, J.; Wier, A.; Boethel, D. Source strength influence on soybean yield formation during early and late reproductive development. Crop Sci. 1995, 35, 1104–1110. [Google Scholar] [CrossRef]
- Liu, X.; Rahman, T.; Song, C.; Su, B.; Yang, F.; Yong, T.; Wu, Y.; Zhang, C.; Yang, W. Changes in light environment, morphology, growth and yield of soybean in maize-soybean intercropping systems. Field Crop. Res. 2017, 200, 38–46. [Google Scholar] [CrossRef]
- Slafer, G.; Calderini, D.; Miralles, D.; Dreccer, M. Preanthesis shading effects on the number of grains of three bread wheat cultivars of different potential number of grains. Field Crop. Res. 1994, 36, 31–39. [Google Scholar] [CrossRef]
- Lesoing, G.W.; Francis, C.A. Strip intercropping effects on yield and yield components of corn, grain sorghum, and soybean. Agron. J. 1999, 91, 807–813. [Google Scholar] [CrossRef]
- Hay, R. Harvest index: A review of its use in plant breeding and crop physiology. Ann. Appl. Biol. 1995, 126, 197–216. [Google Scholar] [CrossRef]
- Egli, D.B.; Guffy, R.; Leggett, J. Partitioning of Assimilate Between Vegetative and Reproductive Growth in Soybean. Agron. J. 1985, 77, 917–922. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar † | Branch Number (Plant−1) ‡ | Growth Habit | Released (year) | Origin from Institute |
|---|---|---|---|---|
| HD19 | 1.4 | Determinate | 2010 | Heze Academy of Agricultural Sciences |
| HD20 | 1.9 | Determinate | 2010 | Heze Academy of Agricultural Sciences |
| ZH39 | 1.8 | Determinate | 2010 | Chinese Academy of Agricultural Sciences |
| QH34 | 1.9 | Determinate | 2012 | Shandong Academy of Agricultural Sciences |
| Phenology | 2015 | 2016 | 2017 | |||
|---|---|---|---|---|---|---|
| Date | Days after Sowing (d) | Date | Days after Sowing (d) | Date | Days after Sowing (d) | |
| Sowing | 17 June | 0 | 14 June | 0 | 13 June | 0 |
| VE | 23 June | 6 | 19 June | 5 | 18 June | 5 |
| R1 | 27 July | 40 (0) † | 22 July | 38 (0) | 22 July | 39 (0) |
| R5 | 20 August | 64 (24) | 17 August | 64 (26) | 17 August | 65 (26) |
| R6 | 6 September | 81 (41) | 2 September | 80 (42) | 1 September | 80 (41) |
| R7 | 21 September | 96 (56) | 18 September | 96 (58) | 18 September | 97 (58) |
| Year | Cultivar | Grain Yield (kg ha−1) | Pod Number (No.m−2) | Seed Number (No.m−2) | Seed Size (mg) | Harvest Index | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M0 | INT1 | INT2 | M0 | INT1 | INT2 | M0 | INT1 | INT2 | M0 | INT1 | INT2 | M0 | INT1 | INT2 | ||
| 2015 | HD19 | 3422a | 950b | 927b | 721a | 217b | 236b | 1446a | 394b | 390b | 237b | 241a | 238b | 0.48a | 0.45a | 0.44b |
| HD20 | 3323a | 979b | 909b | 746a | 239b | 233b | 1353b | 408b | 386b | 245b | 240a | 236b | 0.49a | 0.46a | 0.45ab | |
| ZH39 | 3417a | 1448a | 1214a | 751a | 306a | 323a | 1430a | 603a | 518a | 239b | 240a | 235b | 0.48a | 0.46a | 0.48a | |
| QH34 | 3293a | 1450a | 1198a | 655b | 302a | 310a | 1266c | 572a | 463ab | 260a | 254a | 259a | 0.48a | 0.46a | 0.46ab | |
| 2016 | HD19 | 3471a | 863b | 1083ab | 635a | 212b | 240b | 1475a | 369b | 445a | 235b | 234b | 243ab | 0.47ab | 0.45ab | 0.44b |
| HD20 | 3227b | 982b | 1059b | 624a | 223b | 239b | 1333bc | 404b | 444a | 242ab | 243ab | 239b | 0.48ab | 0.44b | 0.44b | |
| ZH39 | 3301ab | 1256a | 1296a | 638a | 330a | 316a | 1394ab | 530a | 547a | 237b | 237b | 238b | 0.49a | 0.46a | 0.47a | |
| QH34 | 3170b | 1240a | 1218ab | 619a | 305a | 307a | 1238c | 488ab | 473a | 257a | 254a | 258a | 0.46b | 0.45ab | 0.45ab | |
| 2017 | HD19 | 3510a | 1087c | 1031c | 698a | 244b | 250c | 1507a | 489b | 431b | 233b | 223b | 240b | 0.48a | 0.44b | 0.44b |
| HD20 | 3558a | 1137bc | 1098bc | 664b | 235b | 234c | 1489a | 495b | 492b | 239b | 230b | 223b | 0.48a | 0.44b | 0.45ab | |
| ZH39 | 3410a | 1511a | 1472a | 674ab | 334a | 354a | 1512a | 681a | 651a | 226b | 222b | 226b | 0.48a | 0.47a | 0.46a | |
| QH34 | 3456a | 1431ab | 1371ab | 669ab | 318a | 313b | 1326b | 564ab | 537ab | 260a | 254a | 256a | 0.49a | 0.47a | 0.45ab | |
| ANOVA | df | |||||||||||||||
| Cultivar (C) | 3 | 26.1 ** | 67.9 ** | 26.5 ** | 45.5 ** | 9.7 ** | ||||||||||
| Planting pattern (P) | 2 | 3699.9 ** | 4584.7 ** | 2931.9 ** | 1.5 ns | 83.3 ** | ||||||||||
| Year (Y) | 2 | 13.0 ** | 21.2 ** | 20.6 ** | 9.8 ** | 3.2 * | ||||||||||
| C × P | 6 | 11.2 ** | 21.9 ** | 14.5 ** | 1.6 ns | 3.1 ** | ||||||||||
| C × Y | 6 | 0.9 ns | 2.2 ns | 1.1 ns | 1.3 ns | 0.8 ns | ||||||||||
| P × Y | 4 | 2.8 * | 19.2 ** | 1.8 ** | 1.0 ns | 0.5 ns | ||||||||||
| C × P × Y | 12 | 0.5 ns | 2.0 * | 0.4 ns | 0.4 ns | 0.9 ns | ||||||||||
| Year | Cultivar | Reproductive Biomass (g m−2) | Crop Growth Rate (g m−2 d-1) | Partitioning Coefficient (g g−1) | Seed set efFiciency (Seed g−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M0 | INT1 | INT2 | M0 | INT1 | INT2 | M0 | INT1 | INT2 | M0 | INT1 | INT2 | ||
| 2015 | HD19 | 199.6a | 73.6b | 79.0a | 19.5a | 9.0a | 9.1a | 0.34a | 0.27b | 0.29a | 7.2ab | 5.4b | 5.0b |
| HD20 | 182.1b | 73.6b | 79.4a | 20.0a | 9.0a | 9.1a | 0.30b | 0.27b | 0.29a | 7.5ab | 5.5b | 4.9b | |
| ZH39 | 184.2b | 92.7a | 87.1a | 20.1a | 9.9a | 9.8a | 0.30b | 0.31a | 0.30a | 7.8a | 6.5a | 5.9a | |
| QH34 | 180.5b | 87.8a | 82.3a | 19.2a | 9.6a | 9.8a | 0.31ab | 0.31a | 0.28a | 7.0b | 6.5a | 5.6a | |
| 2016 | HD19 | 215.1a | 98.8a | 94.2a | 21.1a | 8.3b | 8.1ab | 0.34a | 0.40a | 0.39ab | 6.9a | 3.7c | 4.7b |
| HD20 | 204.1ab | 93.3a | 94.3a | 20.4ab | 8.7b | 7.7b | 0.33a | 0.36ab | 0.41a | 6.5ab | 4.3bc | 4.7b | |
| ZH39 | 199.5b | 97.8a | 97.8a | 20.0b | 9.5a | 8.7a | 0.33a | 0.34b | 0.37ab | 7.0a | 5.5a | 5.6a | |
| QH34 | 210.1ab | 101.1a | 95.2a | 20.1b | 8.4b | 8.7a | 0.35a | 0.40a | 0.36b | 5.9b | 4.9ab | 5.0ab | |
| 2017 | HD19 | 191.2a | 92.4b | 85.8b | 22.2a | 9.0a | 8.8a | 0.29a | 0.34b | 0.33b | 7.9a | 5.3ab | 5.0a |
| HD20 | 193.8a | 97.8b | 93.4b | 22.7a | 9.5a | 9.1a | 0.28a | 0.35b | 0.34b | 7.7a | 5.0b | 5.3a | |
| ZH39 | 180.2b | 109.4a | 110.2a | 21.3b | 9.7a | 9.3a | 0.28a | 0.37a | 0.39a | 8.4a | 6.2a | 5.9a | |
| QH34 | 178.3b | 106.2a | 104.8a | 21.2b | 9.4a | 9.4a | 0.28a | 0.37a | 0.38a | 7.4a | 5.3ab | 5.1a | |
| ANOVA | df | ||||||||||||
| Cultivar (C) | 3 | 3.4 * | 2.6 ns | 2.2 ns | 19.2 ** | ||||||||
| Planting pattern (P) | 2 | 2907.7 ** | 5451.4 ** | 33.2 ** | 208.2 ** | ||||||||
| Year (Y) | 2 | 63.3 ** | 30.3 ** | 125.0 ** | 37.7 ** | ||||||||
| C × P | 6 | 10.0 ** | 5.8 ** | 4.0 ** | 3.8 ** | ||||||||
| C × Y | 6 | 2.7 * | 1.7 ns | 4.4 ** | 0.9 ns | ||||||||
| P × Y | 4 | 16.2 ** | 19.1 ** | 32.8 ** | 6.8 ** | ||||||||
| C × P × Y | 12 | 1.6 ns | 1.0 ns | 3.3 ** | 0.7 ns | ||||||||
| Source | GY | SN | RB | CGRR1–R5 | PartCoef | SetEff |
|---|---|---|---|---|---|---|
| GY | - | 0.956 ** | 0.646 ** | 0.536 ** | 0.253 | 0.732 ** |
| SN | - | 0.677 ** | 0.556 ** | 0.267 | 0.746 ** | |
| RB | - | 0.001 | 0.838 ** | 0.021 | ||
| CGRR1–R5 | - | −0.536 ** | 0.754 ** | |||
| PartCoef | - | −0.39 | ||||
| SetEff | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wu, X.; Ding, G.; Yang, F.; Yong, T.; Wang, X.; Yang, W. Analysis of Grain Yield Differences Among Soybean Cultivars under Maize–Soybean Intercropping. Agronomy 2020, 10, 110. https://doi.org/10.3390/agronomy10010110
Wang X, Wu X, Ding G, Yang F, Yong T, Wang X, Yang W. Analysis of Grain Yield Differences Among Soybean Cultivars under Maize–Soybean Intercropping. Agronomy. 2020; 10(1):110. https://doi.org/10.3390/agronomy10010110
Chicago/Turabian StyleWang, Xingcai, Xiaoling Wu, Guohui Ding, Feng Yang, Taiwen Yong, Xiaochun Wang, and Wenyu Yang. 2020. "Analysis of Grain Yield Differences Among Soybean Cultivars under Maize–Soybean Intercropping" Agronomy 10, no. 1: 110. https://doi.org/10.3390/agronomy10010110
APA StyleWang, X., Wu, X., Ding, G., Yang, F., Yong, T., Wang, X., & Yang, W. (2020). Analysis of Grain Yield Differences Among Soybean Cultivars under Maize–Soybean Intercropping. Agronomy, 10(1), 110. https://doi.org/10.3390/agronomy10010110