
Activation/Inhibition of Gene Expression Caused by Alcohols: Relationship with the Viscoelastic Property of a DNA Molecule



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Luciferase Assay for Gene Expression (TX-TL)
2.3. Luciferase Assay for translation (TL)
2.4. Fluorescence Microscopy (FM) Observation
3. Results
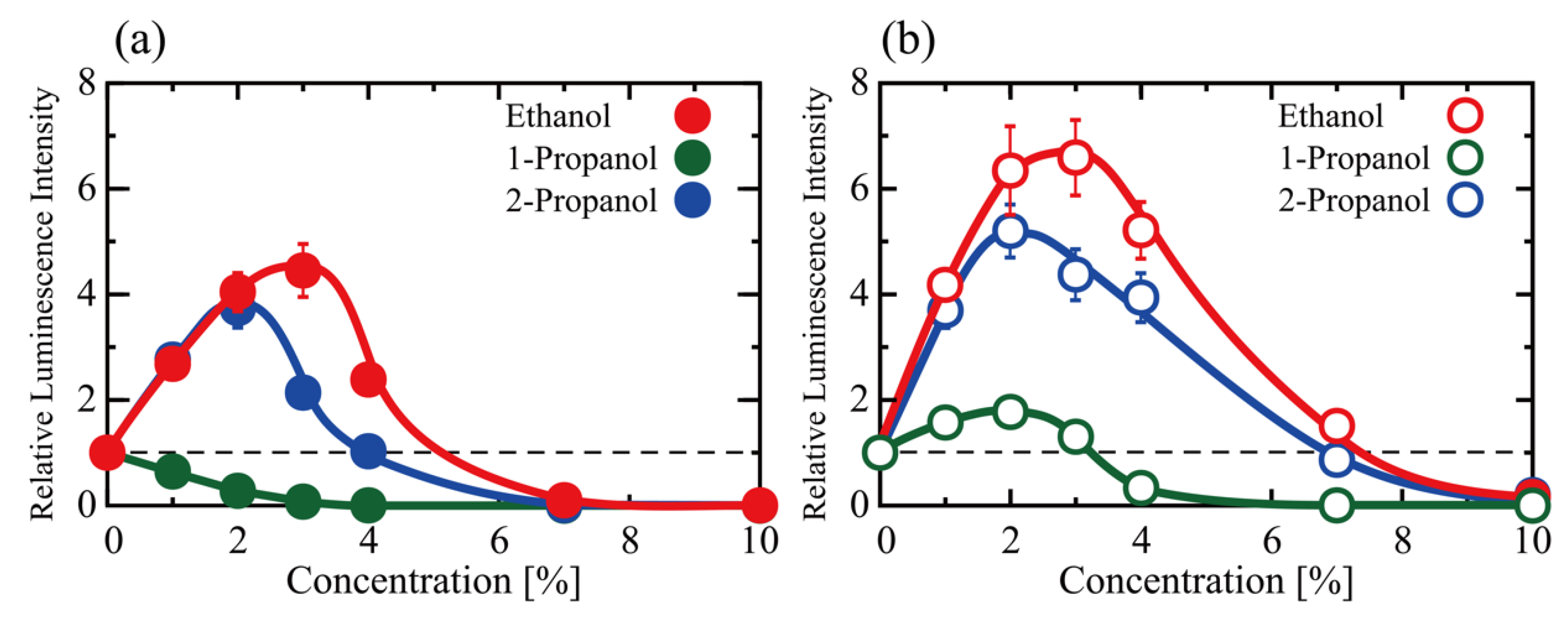
3.1. Effects of Alcohol Concentration on the Efficiency of Gene Transcription/Translation
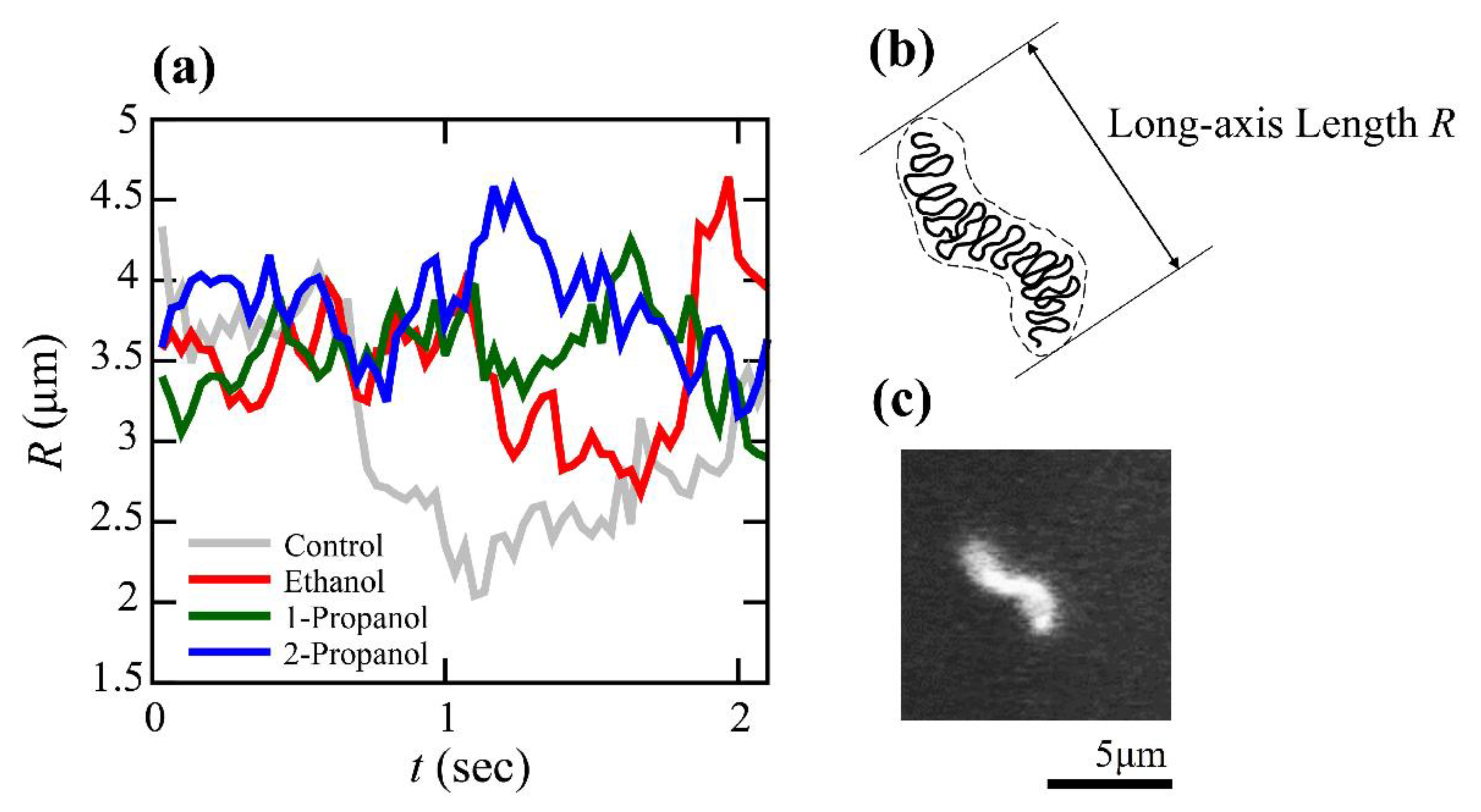
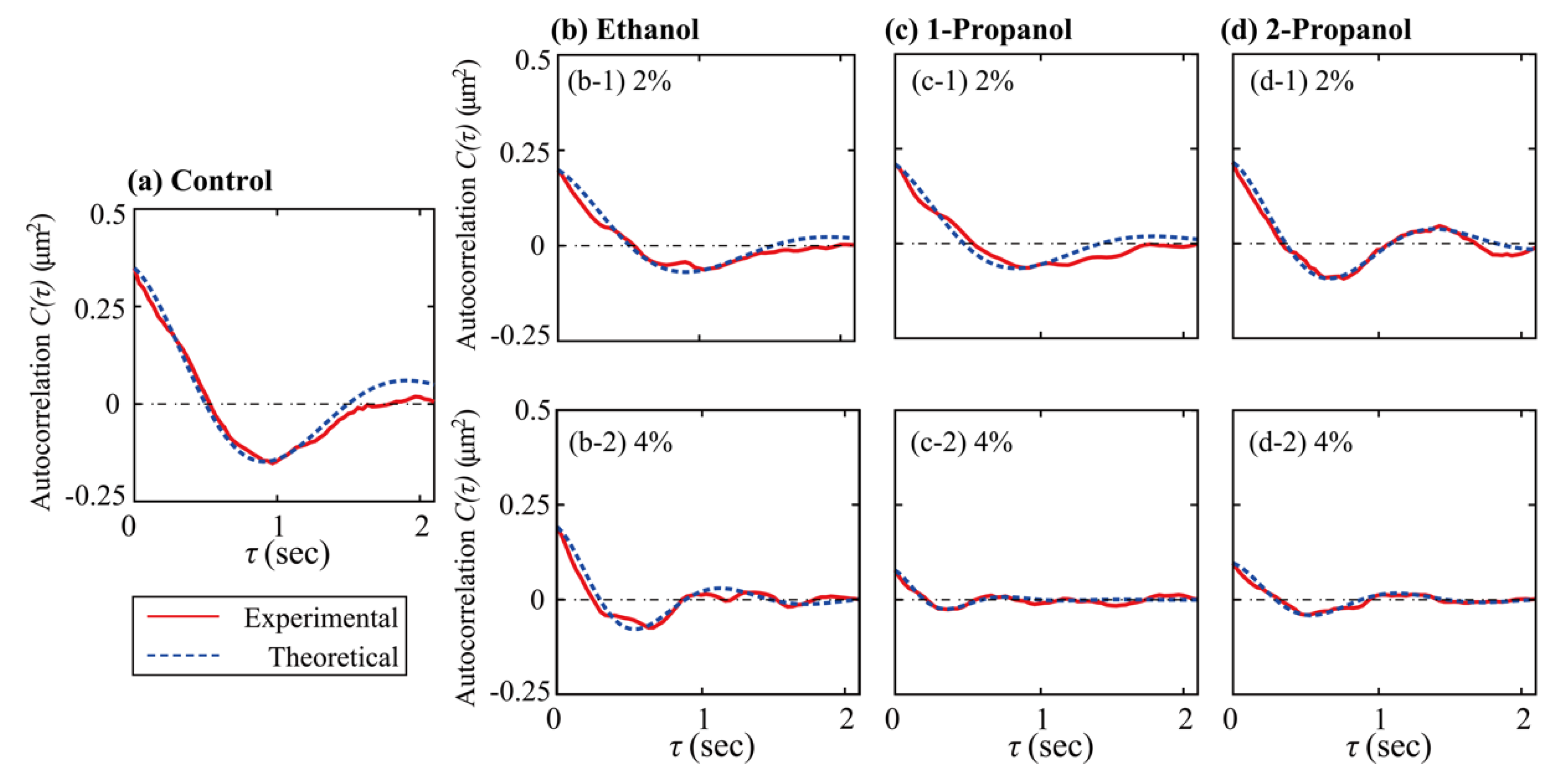
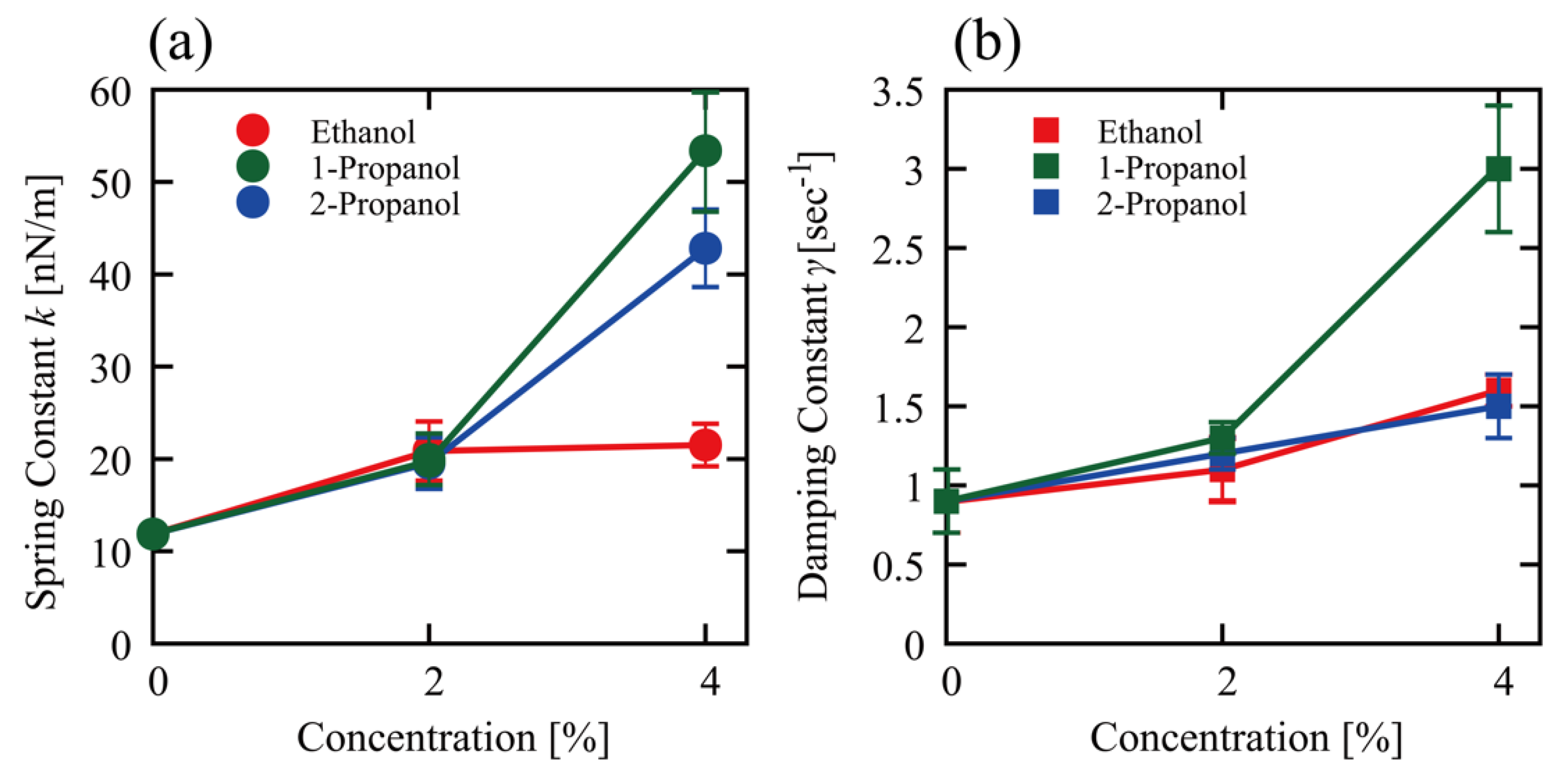
3.2. Effect of Alcohols on the Viscoelasticity of a Single DNA Molecule Evaluated Using Thermal Fluctuations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bloomfield, V.A. DNA condensation. Curr. Opin. Struct. Biol. 1996, 6, 334–341. [Google Scholar] [CrossRef]
- Wang, Y.; Ran, S.; Man, B.; Yang, G. Ethanol induces condensation of single DNA molecules. Soft Matter 2011, 7, 4425–4434. [Google Scholar] [CrossRef]
- Oda, Y.; Sadakane, K.; Yoshikawa, Y.; Imanaka, T.; Takiguchi, K.; Hayashi, M.; Kenmotsu, T.; Yoshikawa, K. Highly concentrated ethanol solutions: Good solvents for DNA as revealed by single-molecule observation. ChemPhysChem 2016, 17, 471–473. [Google Scholar] [CrossRef]
- Marchetti, S.; Onori, G.; Cametti, C. Ethanol-induced compaction of DNA: A viscosimetry and dynamic light scattering study. Philos. Mag. 2007, 87, 525–534. [Google Scholar] [CrossRef]
- Nara-inui, H.; Akutsu, H.; Kyogoku, Y. Alcohol induced BA transition of DNAs with different base compositions studied by circular dichroism. J. Biochem. 1985, 98, 629–636. [Google Scholar] [CrossRef]
- Girod, J.C.; Johnson Jr, W.C.; Huntington, S.K.; Maestre, M.F. Conformation of deoxyribonucleic acid in alcohol solutions. Biochemistry 1973, 12, 5092–5096. [Google Scholar] [CrossRef]
- Ma, Y.; Yoshikawa, Y.; Oana, H.; Yoshikawa, K. Marked difference in the conformational transition of DNA caused by propanol isomer. Polymers 2020, 12, 1607. [Google Scholar] [CrossRef]
- Martino, C.; Zaramela, L.S.; Gao, B.; Embree, M.; Tarasova, J.; Parker, S.J.; Wang, Y.; Chu, H.; Chen, P.; Lee, K.C.; et al. Acetate reprograms gut microbiota during alcohol consumption. Nat. Commun. 2022, 13, 4630. [Google Scholar] [CrossRef]
- Bohnsack, J.P.; Zhang, H.; Wandling, G.M.; He, D.; Kyzar, E.J.; Lasek, A.W.; Pandey, S.C. Targeted epigenomic editing ameliorates adult anxiety and excessive drinking after adolescent alcohol exposure. Sci. Adv. 2022, 8, eabn2748. [Google Scholar] [CrossRef]
- Topiwala, A.; Taschler, B.; Ebmeier, K.P.; Smith, S.; Zhou, H.; Levey, D.F.; Codd, V.; Samani, N.J.; Gelernter, J.; Nichols, T.E.; et al. Alcohol consumption and telomere length: Mendelian randomization clarifies alcohol’s effects. Mol. Psychiatry 2022, 27, 4001–4008. [Google Scholar] [CrossRef]
- Khodja, Y.; Samuels, M.E. Ethanol-mediated upregulation of APOA1 gene expression in HepG2 cells is independent of de novo lipid biosynthesis. Lipids Health Dis. 2020, 19, 144. [Google Scholar] [CrossRef]
- Marballi, K.; Genabai, N.K.; Blednov, Y.A.; Harris, R.A.; Ponomarev, I. Alcohol consumption induces global gene expression changes in VTA dopaminergic neurons. Genes Brain Behav. 2016, 15, 318–326. [Google Scholar] [CrossRef]
- Mignogna, K.M.; Bacanu, S.A.; Riley, B.P.; Wolen, A.R.; Miles, M.F. Cross-species alcohol dependence-associated gene networks: Co-analysis of mouse brain gene expression and human genome-wide association data. PLoS ONE 2019, 14, e0202063. [Google Scholar] [CrossRef]
- Salvatore, J.E.; Han, S.; Farris, S.P.; Mignogna, K.M.; Miles, M.F.; Agrawal, A. Beyond genome-wide significance: Integrative approaches to the interpretation and extension of GWAS findings for alcohol use disorder. Addict. Biol. 2019, 24, 275–289. [Google Scholar] [CrossRef]
- Ivanov, V.I.; Minchenkova, L.E.; Minyat, E.E.; Frank-Kamenetskii, M.D.; Schyolkina, A.K. The B- to A- transition of DNA in solution. J. Mol. Biol. 1974, 87, 817–833. [Google Scholar] [CrossRef]
- Baldini, G.; Fu-Hua, H.; Varani, G.; Cordone, L.; Fornili, S.; Onori, G. DNA melting induced by alcohols: Role of the solvent properties. Il Nuovo Cimento D 1985, 6, 618–630. [Google Scholar] [CrossRef]
- Frisman, E.; Veselkov, A.; Slonitsky, S.; Karavaev, L.; Vorob’ev, V. The influence of alcohol–water solvents on the conformation of deoxyribonucleic acid. Biopolymers 1974, 13, 2169–2178. [Google Scholar] [CrossRef]
- Carnerero, J.M.; Masuoka, S.; Baba, H.; Yoshikawa, Y.; Prado-Gotor, R.; Yoshikawa, K. Decorating a single giant DNA with gold nanoparticles. RSC Adv. 2018, 8, 26571–26579. [Google Scholar] [CrossRef]
- Kanemura, A.; Yoshikawa, Y.; Fukuda, W.; Tsumoto, K.; Kenmotsu, T.; Yoshikawa, K. Opposite effect of polyamines on in vitro gene expression: Enhancement at low concentrations but inhibition at high concentrations. PLoS ONE 2018, 13, e0193595. [Google Scholar] [CrossRef]
- Kitagawa, T.; Nishio, T.; Yoshikawa, Y.; Umezawa, N.; Higuchi, T.; Shew, C.-Y.; Kenmotsu, T.; Yoshikawa, K. Effects of structural isomers of spermine on the higher-order structure of DNA and gene expression. Int. J. Mol. Sci. 2021, 22, 2355. [Google Scholar] [CrossRef]
- Nishio, T.; Sugino, K.; Yoshikawa, Y.; Matsumoto, M.; Oe, Y.; Sadakane, K.; Yoshikawa, K. K+ promotes the favorable effect of polyamine on gene expression better than Na+. PloS ONE 2020, 15, e0238447. [Google Scholar] [CrossRef] [PubMed]
- Nishio, T.; Yoshikawa, Y.; Shew, C.-Y.; Umezawa, N.; Higuchi, T.; Yoshikawa, K. Specific effects of antitumor active norspermidine on the structure and function of DNA. Sci. Rep. 2019, 9, 14971. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.C.; Uhlenbeck, G.E. On the theory of the Brownian motion II. Rev. Mod. Phys. 1945, 17, 323–342. [Google Scholar] [CrossRef]
- Kubo, R. The fluctuation-dissipation theorem. Rep. Prog. Phys. 1966, 29, 255–284. [Google Scholar] [CrossRef]
- Vezeau, G.E.; Salis, H.M. Tuning cell-free composition controls the time delay, dynamics, and productivity of TX-TL expression. ACS Synth. Biol. 2021, 10, 2508–2519. [Google Scholar] [CrossRef]
- Vibhute, M.A.; Schaap, M.H.; Maas, R.J.; Nelissen, F.H.; Spruijt, E.; Heus, H.A.; Hansen, M.M.; Huck, W.T. Transcription and translation in cytomimetic protocells perform most efficiently at distinct macromolecular crowding conditions. ACS Synth. Biol. 2020, 9, 2797–2807. [Google Scholar] [CrossRef]
- Tan, C.; Saurabh, S.; Bruchez, M.P.; Schwartz, R.; LeDuc, P. Molecular crowding shapes gene expression in synthetic cellular nanosystems. Nat. Nanotechnol. 2013, 8, 602–608. [Google Scholar] [CrossRef]
- Zinchenko, A.; Hiramatsu, H.; Yamaguchi, H.; Kubo, K.; Murata, S.; Kanbe, T.; Hazemoto, N.; Yoshikawa, K.; Akitaya, T. Amino acid sequence of oligopeptide causes marked difference in DNA compaction and transcription. Biophys. J. 2019, 116, 1836–1844. [Google Scholar] [CrossRef]
- Yoshinaga, N.; Yoshikawa, K.; Kidoaki, S. Multiscaling in a long semiflexible polymer chain in two dimensions. J. Chem. Phys. 2002, 116, 9926–9929. [Google Scholar] [CrossRef]
- Main, K.H.; Provan, J.I.; Haynes, P.J.; Wells, G.; Hartley, J.A.; Pyne, A.L. Atomic force microscopy—A tool for structural and translational DNA research. APL Bioeng. 2021, 5, 031504. [Google Scholar] [CrossRef]
- Marini, M.; Falqui, A.; Moretti, M.; Limongi, T.; Allione, M.; Genovese, A.; Lopatin, S.; Tirinato, L.; Das, G.; Torre, B. The structure of DNA by direct imaging. Sci. Adv. 2015, 1, e1500734. [Google Scholar] [CrossRef]
- Bell, D.C.; Thomas, W.K.; Murtagh, K.M.; Dionne, C.A.; Graham, A.C.; Anderson, J.E.; Glover, W.R. DNA base identification by electron microscopy. Microsc. Microanal. 2012, 18, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Gao, H.; Bai, X.-c.; Yu, H. Cryo-EM structure of the human cohesin-NIPBL-DNA complex. Science 2020, 368, 1454–1459. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.-C.; Martin, T.G.; Scheres, S.H.; Dietz, H. Cryo-EM structure of a 3D DNA-origami object. Proc. Natl. Acad. Sci. USA 2012, 109, 20012–20017. [Google Scholar] [CrossRef]
- Shon, M.J.; Rah, S.-H.; Yoon, T.-Y. Submicrometer elasticity of double-stranded DNA revealed by precision force-extension measurements with magnetic tweezers. Sci. Adv. 2019, 5, eaav1697. [Google Scholar] [CrossRef] [PubMed]
- Salomo, M.; Kegler, K.; Gutsche, C.; Struhalla, M.; Reinmuth, J.; Skokow, W.; Hahn, U.; Kremer, F. The elastic properties of single double-stranded DNA chains of different lengths as measured with optical tweezers. Colloid Polym. Sci. 2006, 284, 1325–1331. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.D.; Yin, H.; Landick, R.; Gelles, J.; Block, S.M. Stretching DNA with optical tweezers. Biophys. J. 1997, 72, 1335–1346. [Google Scholar] [CrossRef]
- Camunas-Soler, J.; Ribezzi-Crivellari, M.; Ritort, F. Elastic properties of nucleic acids by single-molecule force spectroscopy. Annu. Rev. Biophys. 2016, 45, 65–84. [Google Scholar] [CrossRef]
- Baumann, C.G.; Bloomfield, V.A.; Smith, S.B.; Bustamante, C.; Wang, M.D.; Block, S.M. Stretching of single collapsed DNA molecules. Biophys. J. 2000, 78, 1965–1978. [Google Scholar] [CrossRef]
- Appleyard, D.C.; Vandermeulen, K.; Lee, H.; Lang, M.J. Optical trapping for undergraduates. Am. J. Phys. 2007, 75, 5–14. [Google Scholar] [CrossRef]
- Marko, J.F.; Siggia, E.D. Stretching DNA. Macromolecules 1995, 28, 8759–8770. [Google Scholar] [CrossRef]
- Schiessel, H. The physics of chromatin. J. Phys. Condens. Matter 2003, 15, R699–R774. [Google Scholar] [CrossRef]
- Schroeder, C.M. Single polymer dynamics for molecular rheology. J. Rheol. 2018, 62, 371–403. [Google Scholar] [CrossRef]
- Matsumoto, M.; Sakaguchi, T.; Kimura, H.; Doi, M.; Minagawa, K.; Matsuzawa, Y.; Yoshikawa, K. Direct observation of Brownian motion of macromolecules by fluorescence microscope. J. Polym. Sci. Part B Polym. Phys. 1992, 30, 779–783. [Google Scholar] [CrossRef]
- Yanao, T.; Yoshikawa, K. Chiral symmetry breaking of a double-stranded helical chain through bend-writhe coupling. Phys. Rev. E 2014, 89, 062713. [Google Scholar] [CrossRef] [PubMed]
- Nomidis, S.K.; Skoruppa, E.; Carlon, E.; Marko, J.F. Twist-bend coupling and the statistical mechanics of the twistable wormlike-chain model of DNA: Perturbation theory and beyond. Phys. Rev. E 2019, 99, 032414. [Google Scholar] [CrossRef]
- Moroz, J.D.; Nelson, P. Entropic elasticity of twist-storing polymers. Macromolecules 1998, 31, 6333–6347. [Google Scholar] [CrossRef]
- Marko, J.F.; Siggia, E.D. Bending and twisting elasticity of DNA. Macromolecules 1994, 27, 981–988. [Google Scholar] [CrossRef]
- Nomura, S.i.M.; Tsumoto, K.; Hamada, T.; Akiyoshi, K.; Nakatani, Y.; Yoshikawa, K. Gene expression within cell-sized lipid vesicles. ChemBioChem 2003, 4, 1172–1175. [Google Scholar] [CrossRef]
- Tsuji, A.; Yoshikawa, K. ON− OFF switching of transcriptional activity of large DNA through a conformational transition in cooperation with phospholipid membrane. J. Am. Chem. Soc. 2010, 132, 12464–12471. [Google Scholar] [CrossRef]
- Luckel, F.; Kubo, K.; Tsumoto, K.; Yoshikawa, K. Enhancement and inhibition of DNA transcriptional activity by spermine: A marked difference between linear and circular templates. FEBS Lett. 2005, 579, 5119–5122. [Google Scholar] [CrossRef] [PubMed]
- Yamada, A.; Kubo, K.; Nakai, T.; Yoshikawa, K.; Tsumoto, K. All-or-none switching of transcriptional activity on single DNA molecules caused by a discrete conformational transition. Appl. Phys. Lett. 2005, 86, 223901. [Google Scholar] [CrossRef]
- Tsumoto, K.; Yoshikawa, K. RNA switches the higher-order structure of DNA. Biophys. Chem. 1999, 82, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Akitaya, T.; Tsumoto, K.; Yamada, A.; Makita, N.; Kubo, K.; Yoshikawa, K. NTP concentration switches transcriptional activity by changing the large-scale structure of DNA. Biomacromolecules 2003, 4, 1121–1125. [Google Scholar] [CrossRef] [PubMed]
- Nishio, T.; Yoshikawa, Y.; Yoshikawa, K.; Sato, S.-i. Longer DNA exhibits greater potential for cell-free gene expression. Sci. Rep. 2021, 11, 11739. [Google Scholar] [CrossRef] [PubMed]
- Maeshima, K.; Matsuda, T.; Shindo, Y.; Imamura, H.; Tamura, S.; Imai, R.; Kawakami, S.; Nagashima, R.; Soga, T.; Noji, H. A Transient Rise in Free Mg 2 Ions Released from ATP-Mg Hydrolysis Contributes to Mitotic Chromosome Condensation. Curr. Biol. 2018, 28, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rudiuk, S.; Venancio-Marques, A.; Baigl, D. Enhancement and Modulation of Enzymatic Activity through Higher-Order Structural Changes of Giant DNA–Protein Multibranch Conjugates. Angew. Chem. 2012, 124, 12866–12870. [Google Scholar] [CrossRef]
- Oosawa, F. Polyelectrolytes; Marcel Dekker: New York, NY, USA, 1971. [Google Scholar]
- Manning, G. The molecular theory of polyelectrolyte solutions with applications to the electrostatic properties of polynucleotides. Q. Rev. Biophys. 1978, 2, 179–246. [Google Scholar] [CrossRef]
- Deserno, M.; Holm, C.; May, S. Fraction of condensed counterions around a charged rod: Comparison of Poisson− Boltzmann theory and computer simulations. Macromolecules 2000, 33, 199–206. [Google Scholar] [CrossRef]
- Debye, P.W.; Hückel, E. Zur Theorie der Elektrolyte. I. Gefrierpunktserniedrigung und verwandte Erscheinungen. Physikal. Zeitschr. 1924, 24, 185–206. [Google Scholar]
- Tongu, C.; Kenmotsu, T.; Yoshikawa, Y.; Zinchenko, A.; Chen, N.; Yoshikawa, K. Divalent cation shrinks DNA but inhibits its compaction with trivalent cation. J. Chem. Phys. 2016, 144, 205101. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Mu, Y.; Nordenskiöld, L.; van der Maarel, J.R. Molecular dynamics simulation of multivalent-ion mediated attraction between DNA molecules. Phys. Rev. Lett. 2008, 100, 118301. [Google Scholar] [CrossRef] [PubMed]
- Manning, G. Counterion condensation theory of attraction between like charges in the absence of multivalent counterions. Eur. Phys. J. E 2011, 34, 132. [Google Scholar] [CrossRef]
- Minhas, V.; Sun, T.; Mirzoev, A.; Korolev, N.; Lyubartsev, A.P.; Nordenskiöld, L. Modeling DNA flexibility: Comparison of force fields from atomistic to multiscale levels. J. Phys. Chem. B 2020, 124, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, H.R.; Zhang, H.; Chen, Y.; Bohnsack, J.P.; Shieh, A.W.; Kusumo, H.; Drnevich, J.; Liu, C.; Grayson, D.R.; Maienschein-Cline, M.; et al. Unraveling the epigenomic and transcriptomic interplay during alcohol-induced anxiolysis. Mol. Psychiatry 2022, 27, 4624–4632. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujino, K.; Nishio, T.; Fujioka, K.; Yoshikawa, Y.; Kenmotsu, T.; Yoshikawa, K. Activation/Inhibition of Gene Expression Caused by Alcohols: Relationship with the Viscoelastic Property of a DNA Molecule. Polymers 2023, 15, 149. https://doi.org/10.3390/polym15010149
Fujino K, Nishio T, Fujioka K, Yoshikawa Y, Kenmotsu T, Yoshikawa K. Activation/Inhibition of Gene Expression Caused by Alcohols: Relationship with the Viscoelastic Property of a DNA Molecule. Polymers. 2023; 15(1):149. https://doi.org/10.3390/polym15010149
Chicago/Turabian StyleFujino, Kohei, Takashi Nishio, Keita Fujioka, Yuko Yoshikawa, Takahiro Kenmotsu, and Kenichi Yoshikawa. 2023. "Activation/Inhibition of Gene Expression Caused by Alcohols: Relationship with the Viscoelastic Property of a DNA Molecule" Polymers 15, no. 1: 149. https://doi.org/10.3390/polym15010149
APA StyleFujino, K., Nishio, T., Fujioka, K., Yoshikawa, Y., Kenmotsu, T., & Yoshikawa, K. (2023). Activation/Inhibition of Gene Expression Caused by Alcohols: Relationship with the Viscoelastic Property of a DNA Molecule. Polymers, 15(1), 149. https://doi.org/10.3390/polym15010149